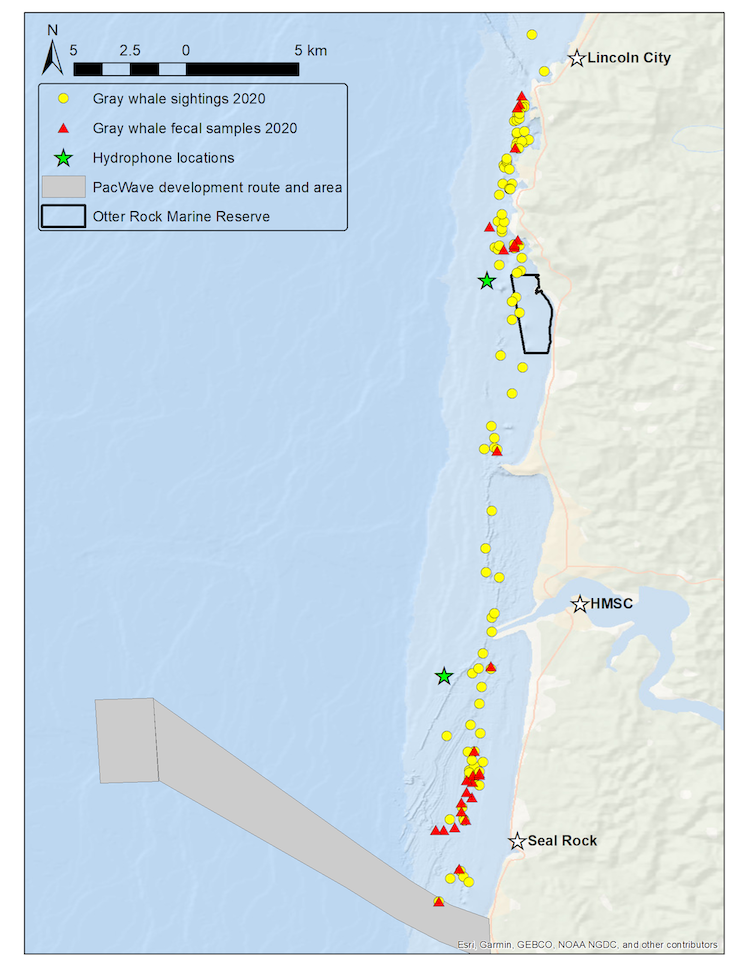
By Jasen C. White, GEMM Lab summer intern, OSU senior, Department of Fisheries, Wildlife, and Conservation Sciences
Field work is predictably unpredictable. Even with years of experience and exhaustive planning, nature always manages to throw a few curveballs, and this gray whale foraging ecology field season is no exception. We are currently in our sixth week of data collection here in Port Orford, and we have been battling the weather, our equipment, and a notable lack of whales and their zooplankton prey. Throughout all of these setbacks, Team “Heck Yeah” has lived up to its mantra as we have approached each day ready to hit the ground running. When faced with any of our myriad of problems, we have managed to work collaboratively to assess our options and develop solutions to keep the project on track.
For those of you that are unfamiliar with Port Orford, it is windy here, and when it is not, it can be foggy. Both of these weather patterns have the potential to make unsafe paddling conditions for our kayak sampling team. This summer we have frequently delayed or altered our field work routines to accommodate these weather patterns. Occasionally, we had to call off kayaking altogether as the winds and swell precluded us from maintaining our boat “on station” at the predetermined GPS coordinates during our samples, only for the winds to die down once we had returned to shore and completed the daily gear maintenance. Despite weather challenges, we have made the most of our data collection opportunities over these past six weeks, and we have only been forced to give up four total days of data collection. Flexibility to take advantage of the good weather windows when they arrive is the key!
Equipment issues can be even more unpredictable than the weather. The first major stumbling block for our equipment was a punctured membrane in the dissolved oxygen probe that we lower into the water at each of our twelve sample locations. This puncture was likely the result of a stray urchin’s spine that was in the wrong place at the wrong time. Soon after noticing the problem, we quickly rallied to refurbish the membrane, recalibrate the sensor, and design a protective housing using some plumbing parts from the local hardware store to prevent any future damage to the membrane (Figures 1a-d). Within 6 days, we were back up and running with the dissolved oxygen sensor.

The next major equipment issue involved a GoPro camera whose mounting hardware snapped while being retrieved at a sample site. This event was captured on the camera itself (see below). Fortunately, thanks to our collaborators at the Oregon Institute of Marine Biology, we were soon able to recover the lost GoPro camera, and in the meantime, we relied on our spare to continue sampling.

The most recent equipment problem was a fraying cable (Figure 2) on our downrigger. We use the downrigger as a winch to lower and raise our sensor array and zooplankton nets into the water to obtain our samples. Fortunately, keen eyes on our team noticed the fray before it fully separated while the sensor array was in the water which could have resulted in losing our gear. We were quickly able to find the necessary repair part locally and get back on the water to finish out our sample regime within an hour of noticing the problem.
Finally, as Damian mentioned in his post last week, this season seemed to start much slower than the previous field seasons. In the early weeks, many of our zooplankton sampling nets repeatedly came up almost empty. There was often nothing but murky water to see in the GoPro videos that accompany the zooplankton samples. Likely due to the lack of prey, we have only managed to spot a couple of transitory whales that rarely entered our study area. Those few whales that we did observe were difficult to track as the relatively high winds and waves quickly dissipated the tell-tale blows and camouflaged their briefly exposed backs and flukes.
Our determination and perseverance have recently started to pay off, however, as the prey abundance in at least some of our sample sites has begun to increase. This increase in prey has also corresponded to a slight increase in whale sightings. One whale even spent nearly 30 minutes around the sampling station that consistently yields the most prey, likely indicating foraging behavior. These modest increases in zooplankton prey and whale sightings provide more evidence in support of the hypothesis Damian mentioned last week that reduced whale abundance in the area is likely the result of low prey abundance.

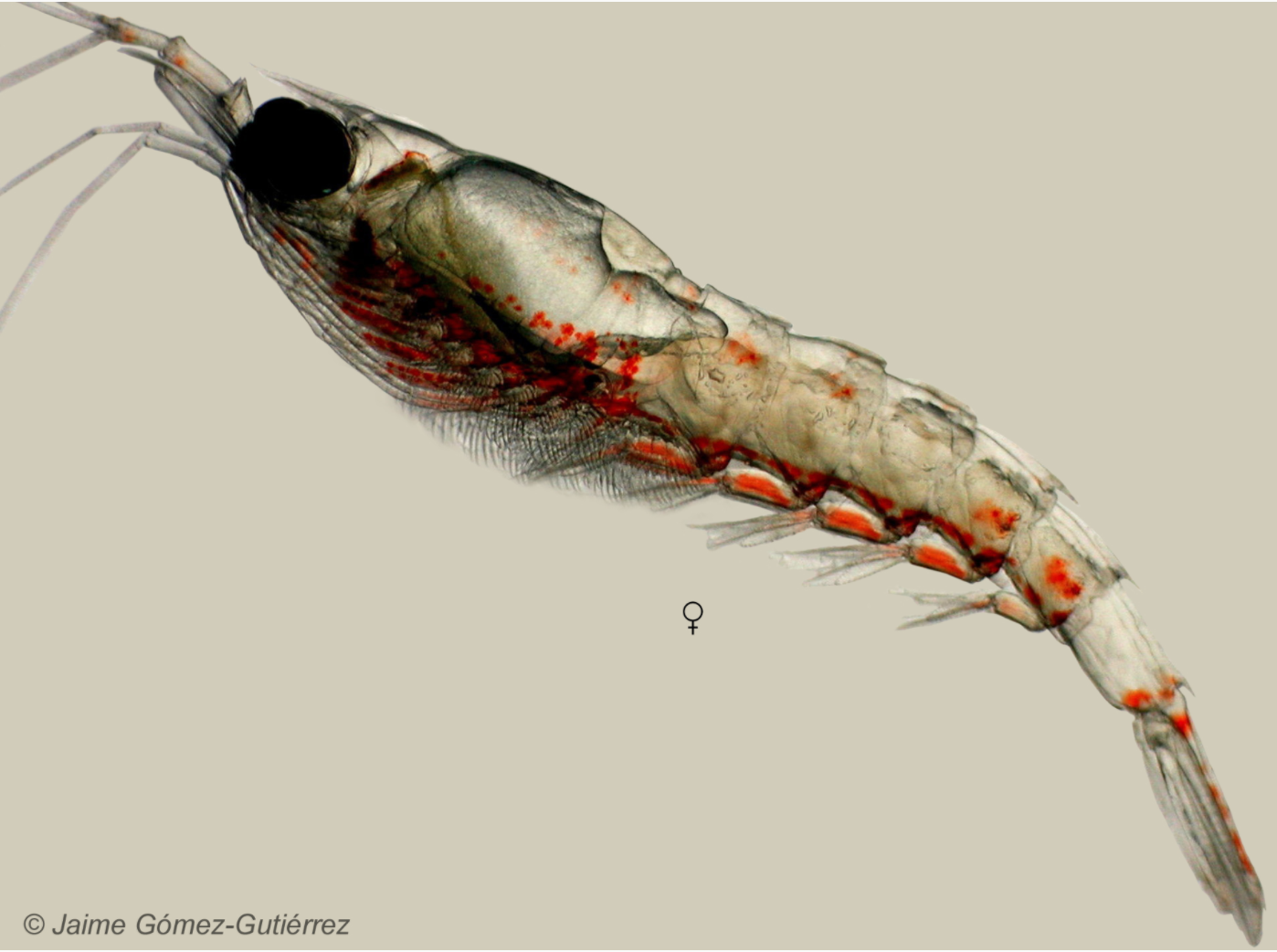
As the zooplankton abundance finally started to increase, we noticed an interesting shift in the kinds of prey that we are capturing compared to previous seasons. Donovan Burns, an intern from the 2019 field season, noted in his blog post that the two most common types of zooplankton they found in their samples were the mysid species Holmesimysis sculpta and members of the genus Neomysis. While Neomysis mysid shrimp are continuing to make up a large proportion of our prey samples this year, we have noticed that many of our samples are dominated by a different type of mysid shrimp (Figure 3) which, in previous years, was a very rare capture. After searching through several mysid identification guides, this unknown mysid appears to be a member of the genus Lucifer, identified based on the presence of some distinctive characteristics that are unique to this genus (Omori 1992).
This observation is interesting because historically, Lucifer mysid shrimp are typically found in warmer tropical and subtropical waters and were rarely reported in the eastern North Pacific Ocean before the year 1992 (Omori 1992). Additionally, a key to common coastal mysid shrimp of Oregon, Washington, and British Columbia does not include members of the Lucifer genus, nor does it include any examples of mysids that resemble these new individuals showing up in our zooplankton nets (Daly and Holmquist 1986). If our initial identification of this mysid species is correct, then the sudden rise in the abundance of a typically warm water mysid species in Port Orford may indicate some fascinating shifts in oceanographic conditions that could lend some insight into why our prey and subsequent whale observations are so different this year than in years past.

As the 2021 field season draws to a close in Port Orford, I cannot help but reflect on what a wonderful opportunity we have been given through this summer internship program. I have loved the short time that I have spent living in this small but lively community for these past five weeks. Most days we could either be found kayaking around the nearshore to sample for the tiny creatures that our local gray whales call dinner, or we were on a cliff, gazing at the tirelessly beautiful, rugged coastline (Figure 4), hoping to glimpse the blow of a foraging whale so that we could track its course with our theodolite. Though the work can be physically exhausting during long and windy kayaking trips, mentally taxing when processing the data for each of the new samples after a full day of fieldwork, or incredibly frustrating with equipment failures, weather delays and shy whales, it is also tremendously satisfying to know that I contributed in a small but meaningful way to the mission of the GEMM Lab. I cannot imagine a better way to obtain the experience that my fellow interns and I have gained from this work, and I know that it will serve each of us well in our future ambitions.
References
Daly, K. L., and C. Holmquist. 1986. A key to the Mysidacea of the Pacific Northwest. Canadian Journal of Zoology 64:1201–1210.
Omori, M. 1992. Occurrence of Two Species of Lucifer (Dendrobranchiata: Sergestoidea: Luciferidae) off the Pacific Coast of America. Journal of Crustacean Biology 12:104–110.


























{kind=link}