1PhD student, Oregon State University College of Earth, Ocean, and Atmospheric Sciences and Department of Fisheries, Wildlife, and Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
What do peanut butter m&ms, killer whales, affogatos, tired eyes, and puffins all have in common? They were all major features of the recent Northern California Current (NCC) ecosystem survey cruise.
The science party of the May 2022 Northern California Current ecosystem cruise.
We spent May 6–17 aboard the NOAA vessel Bell M. Shimada in northern California, Oregon, and Washington waters. This fabulously interdisciplinary cruise studies multiple aspects of the NCC ecosystem three times per year, and the GEMM lab has put marine mammal observers aboard since 2018.
This cruise was a bit different than usual for the GEMM lab: we had eyes on both the whales and their prey. While Dawn Barlow and Clara Bird observed from sunrise to sunset to sight and identify whales, Rachel Kaplan collected krill data via an echosounder and samples from net tows in order to learn about the preyscape the whales were experiencing.
From left, Rachel, Dawn, and Clara after enjoying some beautiful sunset sightings.
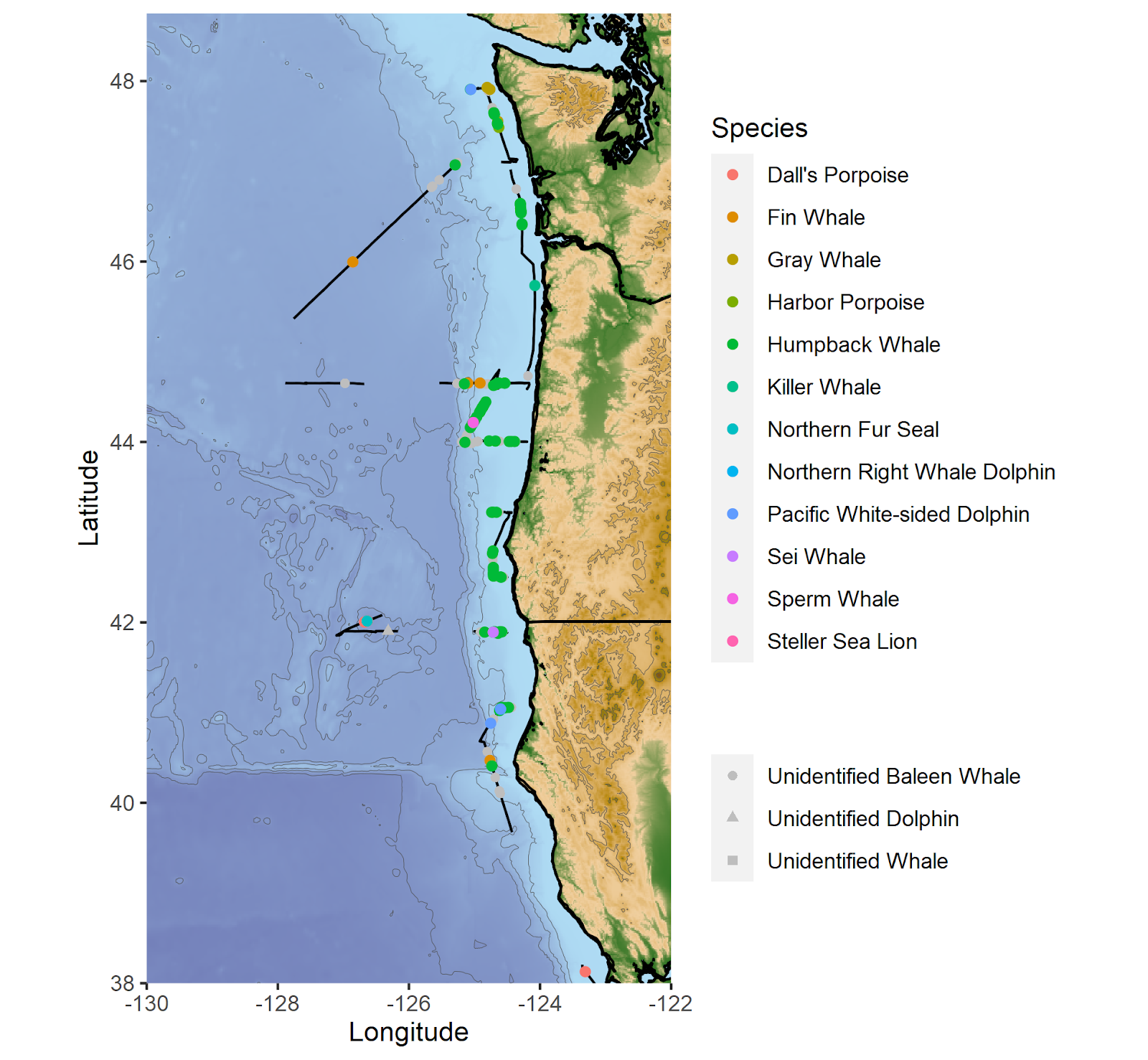
We sailed out of Richmond, California and went north, sampling as far north as La Push, Washington and up to 200 miles offshore. Despite several days of challenging conditions due to wind, rain, fog, and swell, the team conducted a successful marine mammal survey. When poor weather prevented work, we turned to our favorite hobbies of coding and snacking.
Rachel attends “Clara’s Beanbag Coding Academy”.
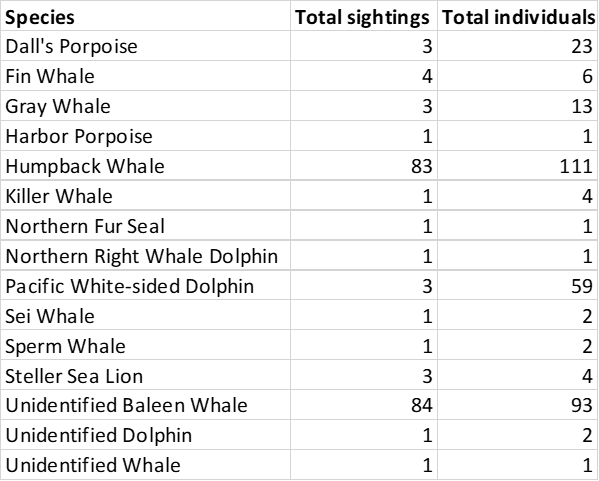
Cruise highlights included several fin whales, sperm whales, killer whales, foraging gray whales, fluke slapping and breaching humpbacks, and a visit by 60 pacific white-sided dolphins. While being stopped at an oceanographic sampling station typically means that we take a break from observing, having more time to watch the whales around us turned out to be quite fortunate on this cruise. We were able to identify two unidentified whales as sei whales after watching them swim near us while paused on station.
Marine mammal observation segments (black lines) and the sighting locations of marine mammal species observed during the cruise.
On one of our first survey days we also observed humpbacks surface lunge feeding close to the ship, which provided a valuable opportunity for our team to think about how to best collect concurrent prey and whale data. The opportunity to hone in on this predator-prey relationship presented itself in a new way when Dawn and Clara observed many apparently foraging humpbacks on the edge of Heceta Bank. At the same time, Rachel started observing concurrent prey aggregations on the echosounder. After a quick conversation with the chief scientist and the officers on the bridge, the ship turned around so that we could conduct a net tow in order to get a closer look at what exactly the whales were eating.
Success! Rachel collects krill samples collected in an area of foraging humpback whales.
This cruise captured an interesting moment in time: southerly winds were surprisingly common for this time of year, and the composition of the phytoplankton and zooplankton communities indicated that the seasonal process of upwelling had not yet been initiated. Upwelling brings deep, cold, nutrient-rich waters to the surface, generating a jolt of productivity that brings the ecosystem from winter into spring. It was fascinating to talk to all the other researchers on the ship about what they were seeing, and learn about the ways in which it was different from what they expected to see in May.
Experiencing these different conditions in the Northern California Current has given us a new perspective on an ecosystem that we’ve been observing and studying for years. We’re looking forward to digging into the data and seeing how it can help us understand this ecosystem more deeply, especially during a period of continued climate change.
The total number of each marine mammal species observed during the cruise.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly message when we post a new blog. Just add your nameand email into the subscribe box below.
Pursuing a graduate degree as a member of the Marine Mammal Institute (MMI) comes with many advantages. Developing associations with curious, industrious researchers and working with advanced technological methods are certainly two of them. Particularly, as a member of the HALO project, I have the pleasure of working alongside not only the GEMM’s, but also acoustician Dr. Holger Klinck and his bioacoustics team at the K. Lisa Yang Center for Conservation Bioacoustics at the Cornell Lab (CCB) who have made significant contributions to advance the field for marine mammal research.
When the HALO project kicked off in October, 2021, Holger and graduate student, Marissa Garcia, arrived for our initial voyage off the Oregon coast with three specialized acoustic recording devices, called Rockhoppers. We deployed each Rockhopper at their designated locations, where they will remain and be replaced every six months, to collect continuous passive acoustic data of cetacean vocalizations. These data are significant because they gather information on all vocalizing whales and dolphins within a detectable range of the Rockhoppers, supporting not only my thesis work concerning fin whale distribution in the Northern California Current (NCC) but has the potential to inform multiple other research projects as well.
Figure 1. Craig Hayslip, Holger Klinck, and Marissa Garcia prepare a Rockhopper for deployment during the first HALO cruise off the Oregon coast.
Passive acoustic monitoring (PAM) is a non-invasive underwater method of recording acoustic output of cetaceans (Zimmer, 2011), and the Rockhopper is specialized for this task. The Rockhopper relatively small (each weighing ~90lbs.) and can be easily deployed with a minimal team from almost any vessel (Fig 1). The mooring is a simple system that anchors the Rockhopper to the sea floor after it sinks through the water column, tolerating depths up to 3,500 m (Klinck et al., 2020). The device can stay on the ocean floor for up to seven months continuously collecting high-frequency data (up to 197 kHz, 24 bits; Klinck et al., 2020). To recover the Rockhopper, the mooring system (Fig 2) includes an acoustic release; when the correct acoustic signal is transmitted by scientists from the vessel and received down at the seafloor, the Rockhopper is released. It’s positive buoyancy allows it to float to the surface where it is recovered. By developing the Rockhopper with these capabilities, the bioacoustics team at Cornell University have taken several steps to enhance cetacean research.
Figure 2. Rockhopper mooring system (Klinck et al., 2020).
According to one of it’s designers, David Winiarski, the Rockhopper development team, consisting of himself, Holger Klinck, Raymond Mack, Christopher Tessaglia-Hymes, Dmitri Ponirakis, Peter Dugan, Christopher Jones, and Haru Matsumoto, initiated it’s construction in 2015. Winiarski states that Jones developed the Rockhopper’s initial PAM electronics at Embedded Ocean Systems (EOS), Boston, MA and then the rest of the team developed the remainder of the device in 2017. The Rockhopper contains the electronic system and a 10.8 V Lithium battery pack in an oil-filled Vitrovex 43 cm glass sphere that is encased in hard polyethelene. Two 64 GB memory cards store the collected acoustic data. About every hour the internal processing unit moves the data to two 4 Terabyte solid-state drives in a process that ensures the data is not lost (Klinck et al., 2020). Winiarski attests that it was quite a hectic process to get six complete Rockhoppers ready for their initial deployment, however the team succeeded and in May 2018 they were deployed in the Gulf of Mexico. The Rockhoppers were recovered in 2019 after six months, returning an amazing 21,522 hours of continuous acoustic data (Klinck et al., 2020).
Learning this information about the acoustic devices that will be responsible for collecting my Master’s thesis data is encouraging. I am eager to see the fin whale energy captured within the Rockhopper records. The HALO team, along with myself, Holger, and Marissa, will head back out off the Oregon coast to retrieve our three HALO-designated Rockhoppers in early June (next month). We will then spend the summer at Cornell reading through our first six months of data.
So, why call this acoustic device, the “Rockhopper”? Winiarski explained that since the CCB is a subsect of the Cornell Lab of Ornithology their projects tend to be named after birds. The Rockhopper team thought that this device should respectively be named after a cool marine megafauna. Hence the rockhopper penguin was chosen. I do agree that such an outstanding device is well suited in relation with an equally remarkable marine species.
Left: Rockhopper penguins on a New Zealand hillside. https://nzbirdsonline.org.nz/species/eastern-rockhopper-penguin Upper right: Chris Tessaglia-Hymes and David Winiarski with a Rockhopper acoustic device. Lower right: The first six complete Rockhopper acoustic devices developed at the Cornell Center of Bioacoustics in 2017.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get weekly updates and more! Just add your name into the subscribe box on the left panel.
References
Klinck, H., Winiarski, D., Mack, R., Tessaglia-Hymes, C., Ponirakis, D., Dugan, P., Jones, C., Matsumoto, H. 2020. The Rockhopper: a compact and extensible marine autonomous passive acoustic recording system,” Global Oceans 2020: Singapore – U.S. Gulf Coast: 1-7. https://ieeexplore.ieee.org/document/9388970.
Zimmer, W. 2011. Passive acoustic monitoring of cetaceans. Cambridge University Press, Cambridge, UK.
In September 2020, I was hired as a postdoc in the GEMM Lab and was tasked to conduct the analyses necessary for the OPAL project. This research project has the ambitious, yet essential, goal to fill a knowledge gap hindering whale conservation efforts locally: where and when do whales occur off the Oregon coast? Understanding and predicting whale distribution based on changing environmental conditions is a key strategy to assess and reduce spatial conflicts with human activities, specifically the risk of entanglement in fixed fishing gear.
Starting a new project is always a little daunting. Learning about a new region and new species, in an alien research and conservation context, is a challenge. As I have specialized in data science over the last couple of years, I have been confronted many times with the prospect of working with massive datasets collected by others, from which I was asked to tease apart the biases and the ecological patterns. In fact, I have come to love that part of my job: diving down the data rabbit hole and making my way through it by collaborating with others. Craig Hayslip, faculty research assistant in MMI, was the observer who conducted the majority of the 102 helicopter surveys that were used for this study. During the analysis stage, his help was crucial to understand the data that had been collected and get a better grasp of the field work biases that I would later have to account for in my models. Similarly, it took hours of zoom discussions with Dawn Barlow, the GEMM lab’s latest Dr, to be able to clean and process the 75 days of survey effort conducted at sea, aboard the R/V Shimada and Oceanus.
Once the data is “clean”, then comes the time for modeling. Running hundreds of models, with different statistical approaches, different environmental predictors, different parameters etc. etc. That is when you realize what a blessing it is to work with a supervisor like Leigh Torres, head of the GEMM Lab. As an early career researcher, I really appreciate working with people who help me take a step back and see the bigger picture within which the whole data wrangling work is included. It is so important to have someone help you stay focused on your goals and the ecological questions you are trying to answer, as these may easily get pushed back to the background during the data analysis process.
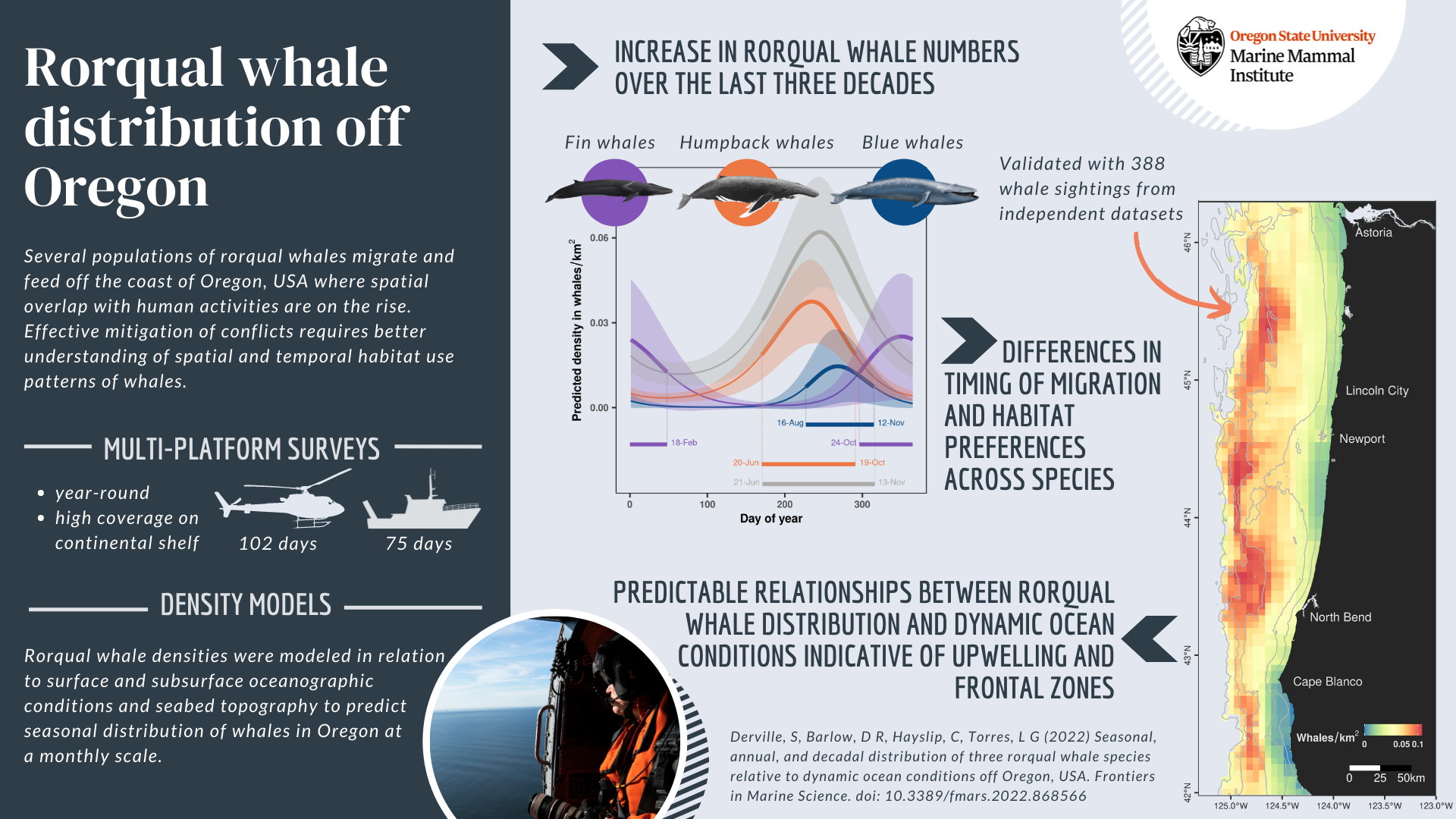
And here we are today, with the first scientific publication from the OPAL project published, a little more than three years after Leigh and Craig started collecting data onboard the United States Coast Guard helicopters off the coast of Oregon in February 2019. Entitled “Seasonal, annual, and decadal distribution of three rorqual whale species relative to dynamic ocean conditions off Oregon, USA”, our study published in Frontiers in Marine Science presents modern and fine-scale predictions of rorqual whale distribution off Oregon, as well as a description of their phenology and a comparison to whale numbers observed across three decades in the region (Figure 1). This research focuses on three rorqual species sharing some ecological and biological traits, as well as similar conservation status: humpback whales (Megaptera novaeangliae), blue whales (Balaenoptera musculus musculus), and fin whales (Balaenoptera physallus); all of which migrate and feed over the US West coast (see a previous blog to learn more about these species here).
Figure 1: Graphical abstract of our latest paper published in Frontiers in Marine Science.
We demonstrate (1) an increase in rorqual numbers over the last three decades in Oregon waters, (2) differences in timing of migration and habitat preferences between humpback, blue, and fin whales, and (3) predictable relationships of rorqual whale distribution based on dynamic ocean conditions indicative of upwellings and frontal zones. Indeed, these ocean conditions are likely to provide suitable biological conditions triggering increased prey abundance. Three seasonal models covering the months of December-March (winter model), April-July (spring) and August-November (summer-fall) were generated to predict rorqual whale densities over the Oregon continental shelf (in waters up to 1,500 m deep). As a result, maps of whale densities can be produced on a weekly basis at a resolution of 5 km, which is a scale that will facilitate targeted management of human activities in Oregon. In addition, species-specific models were also produced over the period of high occurrence in the region; that is humpback and blue whales between April and November, and fin whales between August and March.
As we outline in our concluding remarks, this work is not to be considered an end-point, but rather a stepping stone to improve ecological knowledge and produce operational outputs that can be used effectively by managers and stakeholders to prevent spatial conflict between whales and human activities. As of today, the models of fin and blue whale densities are limited by the small number of observations of these two species over the Oregon continental shelf. Yet, we hope that continued data collection via fruitful research partnerships will allow us to improve the robustness of these species-specific predictions in the future. On the other hand, the rorqual models are considered sufficiently robust to continue into the next phase of the OPAL project that aims to assess overlap between whale distribution and Dungeness crab fishing gear to estimate entanglement risk.
The curse (or perhaps the beauty?) of species distribution modeling is that it never ends. There are always new data to be added, new statistical approaches to be tested, and new predictions to be made. The OPAL models are no exception to this rule. They are meant to be improved in future years, thanks to continued helicopter and ship-based survey efforts, and to the addition of new environmental variables meant to better predict whale habitat selection. For instance, Rachel Kaplan’s PhD research specifically aims at understanding the distribution of whales in relation to krill. Her results will feed into the more general efforts to model and predict whale distribution to inform management in Oregon.
This first publication therefore paves the way for more exciting and impactful research!
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly message when we post a new blog. Just add your name and email into the subscribe box below.
Reference
Derville, S., Barlow, D. R., Hayslip, C. E., and Torres, L. G. (2022). Seasonal, Annual, and Decadal Distribution of Three Rorqual Whale Species Relative to Dynamic Ocean Conditions Off Oregon, USA. Front. Mar. Sci. 9, 1–19. doi:10.3389/fmars.2022.868566.
Acknowledgments
We gratefully acknowledge the immense contribution of the United State Coast Guard sectors North Bend and Columbia River who facilitated and piloted our helicopter surveys. We would like to also thank NOAA Northwest Fisheries Science Center for the ship time aboard the R/V Bell M. Shimada. We thank the R/V Bell M. Shimada (chief scientists J. Fisher and S. Zeman) and R/V Oceanus crews, as well as the marine mammal observers F. Sullivan, C. Bird and R. Kaplan. We give special recognition and thanks to the late Alexa Kownacki who contributed so much in the field and to our lives. We also thank T. Buell and K. Corbett (ODFW) for their partnership over the OPAL project. We thank G. Green and J. Brueggeman (Minerals Management Service), J. Adams (US Geological Survey), J. Jahncke (Point blue Conservation), S. Benson (NOAA-South West Fisheries Science Center), and L. Ballance (Oregon State University) for sharing validation data. We thank J. Calambokidis (Cascadia Research Collective) for sharing validation data and for logistical support of the project. We thank A. Virgili for sharing advice and custom codes to produce detection functions.
Dr. KC Bierlich, Postdoctoral Scholar, OSU Department of Fisheries, Wildlife, & Conservation Sciences, Geospatial Ecology of Marine Megafauna (GEMM) Lab
In a previous blog, I discussed the importance of incorporating measurement uncertainty in drone-based photogrammetry, as drones with different sensors, focal length lenses, and altimeters will have varying levels of measurement accuracy. In my last blog, I discussed how to incorporate photogrammetric uncertainty when combining multiple measurements to estimate body condition of baleen whales. In this blog, I will highlight our recent publication in Frontiers in Marine Science (https://doi.org/10.3389/fmars.2022.867258) led by GEMM Lab’s Dr. Leigh Torres, Clara Bird, and myself that used these methods in a collaborative study using imagery from four different drones to compare gray whale body condition on their breeding and feeding grounds (Torres et al., 2022).
Most Eastern North Pacific (ENP) gray whales migrate to their summer foraging grounds in Alaska and the Arctic, where they target benthic amphipods as prey. A subgroup of gray whales (~230 individuals) called the Pacific Coast Feeding Group (PCFG), instead truncates their migration and forages along the coastal habitats between Northern California and British Columbia, Canada (Fig. 1). Evidence from a recent study lead by GEMM Lab’s Lisa Hildebrand (see this blog) found that the caloric content of prey in the PCFG range is of equal or higher value than the main amphipod prey in the Arctic/sub-Arctic regions (Hildebrand et al., 2021). This implies that greater prey density and/or lower energetic costs of foraging in the Arctic/sub-Arctic may explain the greater number of whales foraging in that region compared to the PCFG range. Both groups of gray whales spend the winter months on their breeding and calving grounds in Baja California, Mexico.
Figure 1. The GEMM Lab field team following a Pacific Coast Feeding Group (PCFG) gray whale swimming in a kelp bed along the Oregon Coast during the summer foraging season.
In January 2019 an Unusual Mortality Event (UME) was declared for gray whales due to the elevated numbers of stranded gray whales between Mexico and the Arctic regions of Alaska. Most of the stranded whales were emaciated, indicating that reduced nutrition and starvation may have been the causal factor of death. It is estimated that the population dropped from ~27,000 individuals in 2016 to ~21,000 in 2020 (Stewart & Weller, 2021).
During this UME period, between 2017-2019, the GEMM Lab was using drones to monitor the body condition of PCFG gray whales on their Oregon coastal feeding grounds (Fig. 1), while Christiansen and colleagues (2020) was using drones to monitor gray whales on their breeding grounds in San Ignacio Lagoon (SIL) in Baja California, Mexico. We teamed up with Christiansen and colleagues to compare the body condition of gray whales in these two different areas leading up to the UME. Comparing the body condition between these two populations could help inform which population was most effected by the UME.
The combined datasets consisted of four different drones used, thus different levels of photogrammetric uncertainty to consider. The GEMM Lab collected data using a DJI Phantom 3 Pro, DJI Phantom 4, and DJI Phantom 4 Pro, while Christiansen et al., (2020) used a DJI Inspire 1 Pro. By using the methodological approach described in my previous blog (here, also see Bierlich et al., 2021a for more details), we quantified photogrammetric uncertainty specific to each drone, allowing cross-comparison between these datasets. We also used Body Area Index (BAI), which is a standardized relative measure of body condition developed by the GEMM Lab (Burnett et al., 2018) that has low uncertainty with high precision, making it easier to detect smaller changes between individuals (see blog here, Bierlich et al., 2021b).
While both PCFG and ENP gray whales visit San Ignacio Lagoon in the winter, we assume that the photogrammetry data collected in the lagoon is mostly of ENP whales based on their considerably higher population abundance. We also assume that gray whales incur low energetic cost during migration, as gray whale oxygen consumption rates and derived metabolic rates are much lower during migration than on foraging grounds (Sumich, 1983).
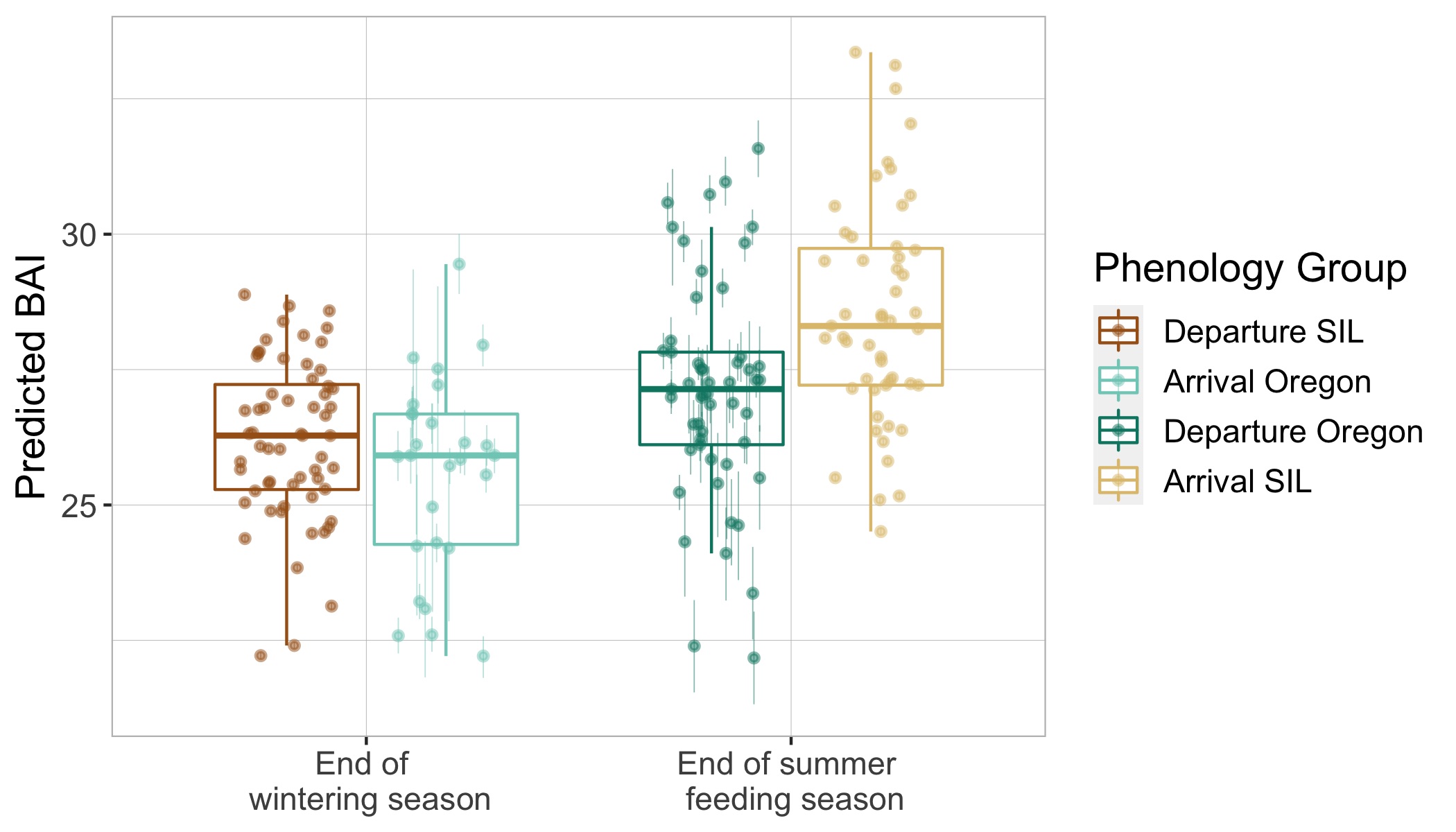
Interestingly, we found that gray whale body condition on their wintering grounds in San Ignacio Lagoon deteriorated across the study years leading up to the UME (2017-2019), while the body condition of PCFG whales on their foraging grounds in Oregon concurrently increased. These contrasting trajectories in body condition between ENP and PCFG whales implies that dynamic oceanographic processes may be contributing to temporal variability of prey available in the Arctic/sub-Arctic and PCFG range. In other words, environmental conditions that control prey availability for gray whales are different in the two areas. For the ENP population, this declining nutritive gain may be associated with environmental changes in the Arctic/sub-Arctic region that impacted the predictability and availability of prey. For the PCFG population, the increase in body condition across years may reflect recovery of the NE Pacific Ocean from the marine heatwave event in 2014-2016 (referred to as “The Blob”) that resulted with a period of low prey availability. These findings also indicate that the ENP population was primarily impacted in the die-off from the UME.
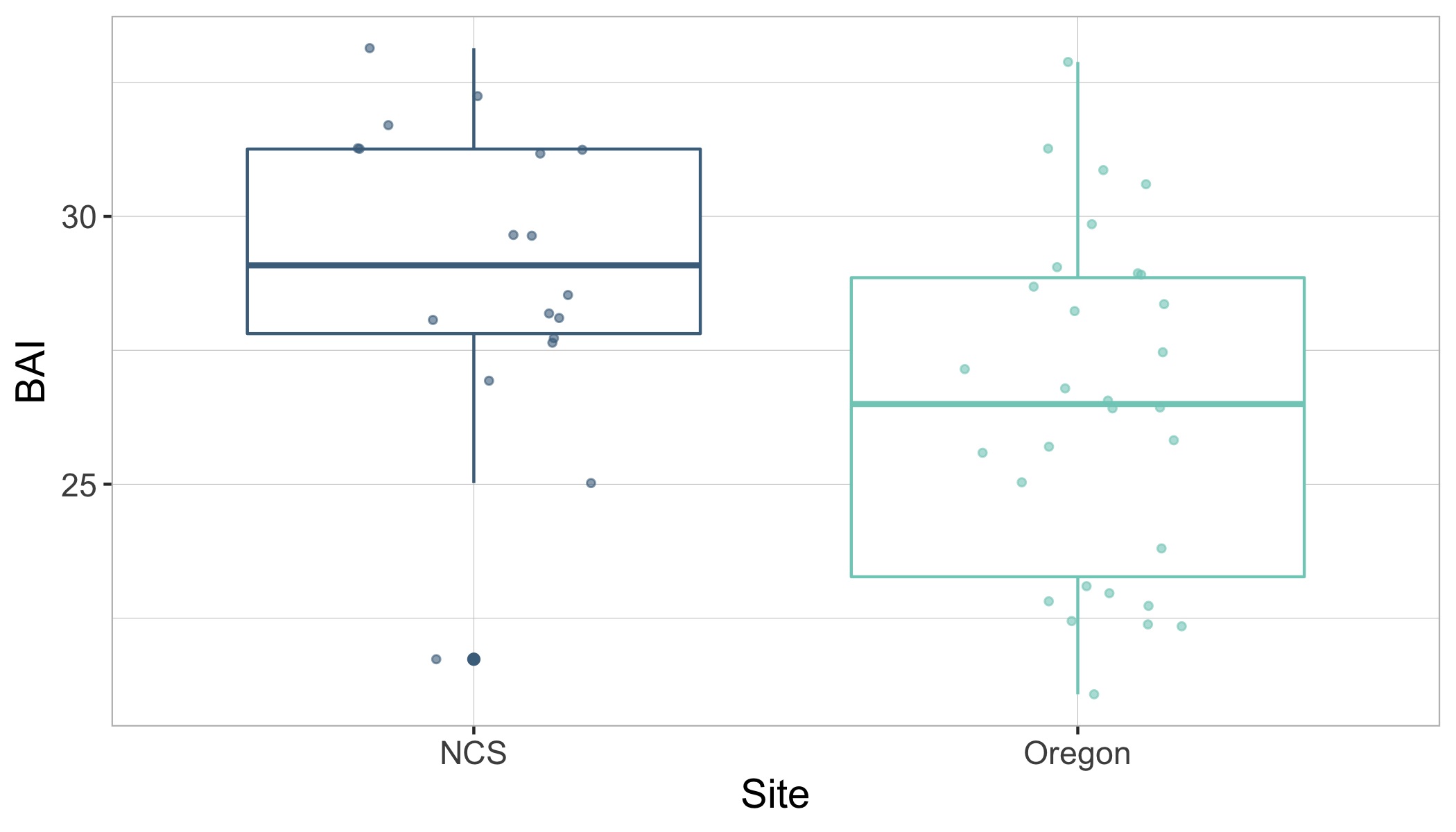
Surprisingly, the body condition of PCFG gray whales in Oregon was regularly and significantly lower than whales in San Ignacio Lagoon (Fig. 2). To further investigate this potential intrinsic difference in body condition between PCFG and ENP whales, we compared opportunistic photographs of gray whales feeding in the Northeastern Chukchi Sea (NCS) in the Arctic collected from airplane surveys. We found that the body condition of PCFG gray whales was significantly lower than whales in the NCS, further supporting our finding that PCFG whales overall have lower body condition than ENP whales that feed in the Arctic (Fig. 3).
Figure 2. Boxplots showing the distribution of Body Area Index (BAI) values for gray whales imaged by drones in San Ignacio Lagoon (SIL), Mexico and Oregon, USA. The data is grouped by phenology group: End of summer feeding season (departure Oregon vs. arrival SIL) and End of wintering season (arrival Oregon vs. departure SIL). The group median (horizontal line), interquartile range (IQR, box), maximum and minimum 1.5*IQR (vertical lines), and outliers (dots) are depicted in the boxplots. The overlaid points represent the mean of the posterior predictive distribution for BAI of an individual and the bars represents the uncertainty (upper and lower bounds of the 95% HPD interval). Note how PCFG whales at then end of the feeding season (dark green) typically have lower body condition (as BAI) compared to ENP whales at the end of the feeding season when they arrive to SIL after migration (light brown).Figure 3. Boxplots showing the distribution of Body Area Index (BAI) values of gray whales from opportunistic images collected from a plane in Northeaster Chukchi Sea (NCS) and from drones collected by the GEMM Lab in Oregon. The boxplots display the group median (horizontal line), interquartile range (IQR box), maximum and minimum 1.5*IQR (vertical lines), and outlies (dots). The overlaid points are the BAI values from each image. Note the significantly lower BAI of PCFG whales on Oregon feeding grounds compared to whales feeding in the Arctic region of the NCS.
This difference in body condition between PCFG and ENP gray whales raises some really interesting and prudent questions. Does the lower body condition of PCFG whales make them less resilient to changes in prey availability compared to ENP whales, and thus more vulnerable to climate change? If so, could this influence the reproductive capacity of PCFG whales? Or, are whales that recruit into the PCFG adapted to a smaller morphology, perhaps due to their specialized foraging tactics, which may be genetically inherited and enables them to survive with reduced energy stores?
These questions are on our minds here at the GEMM Lab as we prepare for our seventh consecutive field season using drones to collect data on PCFG gray whale body condition. As discussed in a previous blog by Dr. Alejandro Fernandez Ajo, we are combining our sightings history of individual whales, fecal hormone analyses, and photogrammetry-based body condition to better understand gray whales’ reproductive biology and help determine what the consequences are for these PCFG whales with lower body condition.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly message when we post a new blog. Just add your name and email into the subscribe box below.
References
Bierlich, K. C., Hewitt, J., Bird, C. N., Schick, R. S., Friedlaender, A., Torres, L. G., … & Johnston, D. W. (2021). Comparing Uncertainty Associated With 1-, 2-, and 3D Aerial Photogrammetry-Based Body Condition Measurements of Baleen Whales. Frontiers in Marine Science, 1729.
Bierlich, K. C., Schick, R. S., Hewitt, J., Dale, J., Goldbogen, J. A., Friedlaender, A.S., et al. (2021b). Bayesian Approach for Predicting Photogrammetric Uncertainty in Morphometric Measurements Derived From Drones. Mar. Ecol. Prog. Ser. 673, 193–210. doi: 10.3354/meps13814
Burnett, J. D., Lemos, L., Barlow, D., Wing, M. G., Chandler, T., & Torres, L. G. (2018). Estimating morphometric attributes of baleen whales with photogrammetry from small UASs: A case study with blue and gray whales. Marine Mammal Science, 35(1), 108–139.
Christiansen, F., Rodrı́guez-González, F., Martı́nez-Aguilar, S., Urbán, J., Swartz, S., Warick, H., et al. (2021). Poor Body Condition Associated With an Unusual Mortality Event in Gray Whales. Mar. Ecol. Prog. Ser. 658, 237–252. doi:10.3354/meps13585
Hildebrand, L., Bernard, K. S., and Torres, L. G. (2021). Do Gray Whales Count Calories? Comparing Energetic Values of Gray Whale Prey Across Two Different Feeding Grounds in the Eastern North Pacific. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.683634
Stewart, J. D., and Weller, D. (2021). Abundance of Eastern North Pacific Gray Whales 2019/2020 (San Diego, CA: NOAA/NMFS)
Sumich, J. L. (1983). Swimming Velocities, Breathing Patterns, and Estimated Costs of Locomotion in Migrating Gray Whales, Eschrichtius Robustus. Can. J. Zoology. 61, 647–652. doi: 10.1139/z83-086
Torres, L.G., Bird, C., Rodrigues-Gonzáles, F., Christiansen F., Bejder, L., Lemos, L., Urbán Ramírez, J., Swartz, S., Willoughby, A., Hewitt., J., Bierlich, K.C. (2022). Range-wide comparison of gray whale body condition reveals contrasting sub-population health characteristics and vulnerability to environmental change. Frontiers in Marine Science. 9:867258. https://doi.org/10.3389/fmars.2022.867258
Allison Dawn, GEMM Lab Master’s student, OSU Department of Fisheries, Wildlife and Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
As the first year of my Master’s is coming to an end, I am excited to have completed the first milestone of writing my research proposal. During the formation of my initial hypotheses, I have been thinking deeply about the potential drivers of zooplankton variability, and how these metrics relate to the Pacific Coast Feeding Group (PCFG) of gray whales foraging in Port Orford. One topic that continues to appear in the literature and throughout my coursework is that of the extreme marine heat wave (MHW) event (2013-2016) in the Pacific Ocean, otherwise known as the “warm blob”. In Dawn’s (now Dr. Barlow!) blog about this MHW, she discusses how whale habitat in California was compressed due to shifts in prey availability, and how this led to an increased number of whale entanglements (Santora et al., 2020). While sea surface temperature (SST) is only one of many factors that influence prey metrics, it is nevertheless an important factor to consider, especially as these heat waves are expected to increase in intensity and duration due to climate change (Joh and Di Lorenzo, 2017). As Lisa mentioned in her last blog, the “warm blob” exacerbated the loss of kelp and sea stars, which is now impacting multiple trophic levels in Port Orford. For my first thesis chapter, I plan to dive into how SST anomalies impact the mosaic of interactions at our study site in Port Orford, and ultimately try to better understand food availability for the PCFG whales.
Cavole et al., 2016 is one of the early comprehensive studies to discuss the impact of the blob on a variety of planktonic marine species. Their sea surface temperature anomaly figure (Figure 1) shows where the anomaly began in 2013 and how it migrated from the Northern Pacific to the Southern Pacific coast.
Figure 1. Plots showing the SST anomalies as the “warm blob” migrated from the Northern Pacific to the Southern Pacific from 2013 until 2016.
Among many other impacts, this MHW caused a reduction in phytoplankton, the major food source for zooplankton. The decline of this food source subsequently caused significant changes in zooplankton populations. Specifically, studies on copepod diversity and biomass show that in a typical California Current System (CCS) there is a seasonal oscillation between warm-water with subtropical species and cold-water with subarctic species. In the winter, the CCS is characterized by a high diversity of subtropical species, due to a southern water source. In the spring, northern cold water advection brings low-diversity, subarctic copepods. While the timing of these shifts is subject to change due to changes in the Pacific Decadal Oscillation (PDO), it remains that these subtropical copepod species are known to be smaller and less nutritious than subarctic copepod species regardless of arrival time (Kintisch, 2015; Leising et al., 2015). However, in 2015, this shift to cold water copepod species did not occur, but rather coastal sampling along the Oregon coast saw subtropical copepod species prevail. Specifically, there were 17 main subtropical copepod species that dominated the species composition while the nutrient-rich arctic species were rare. This occurrence of major copepod shifts alone points to the overall concern for the ecosystem imbalance, to the detriment of top predators like marine mammals and seabirds (the “losers”), and others gaining advantage (the “winners”) (Figure 2).
Figure 2. Figure showing the “losers” (right column) and “winners” (left column) of MHW impacts. Species are organized by trophic level, with top predators at the bottom. Taken from Cavole et al., 2016.
More recent studies found that in certain areas, impacts from the “warm blob” outlived the duration of the larger scale anomaly. In fact, large, positive SST anomalies have lingered on the Oregon shelf until at least September 2017 (Peterson et al., 2017). During this time period, anomalously high abundances of nearshore larval North Pacific krill (Euphausia pacifica) were collected off of the Newport Hydrographic Station (Morgan et al., 2019). Additionally, Brodeur et al. (2019) demonstrate that while indicator species in the nearshore have consistent annual variability, there were substantial differences between community composition between 2011-2014 (low diversity) and 2015-2016 (high diversity). This work also documented the shift from crustacean species (like krill and mysids) to more low-quality gelatinous taxa. As the authors acknowledge, this change in prey community assemblage could have major negative impacts on trophic interactions. This is especially true in the context of whales, as they are not known to rely on gelatinous taxa for energy.
Just like our summer sampling in Port Orford, these studies only provide a “snapshot” of plankton species abundance and composition during a particular time of year. However, even a snapshot can reveal significant changes in prey variability, which then may help us understand the drivers of PCFG habitat utilization. We are actively investigating whether there have been significant changes in the variability of several zooplankton metrics (abundance, distribution, size class, composition) relative to SST changes in Port Orford over the last 6 years (2016-2021).
We will also consider multiple other static and dynamic factors that could influence zooplankton patterns (e.g., upwelling strength, kelp health, tidal height, topography); however, given these documented strong relationships between the zooplankton community and SST across the North Pacific, we hypothesize similar impacts in our Port Orford study region. For example, in certain sampling years, net tows seemed to be comprised of smaller size classes of zooplankton than usual. We will consider how size class availability has changed and if this was driven by SST variability. Gray whales are drawn to this area for enhanced feeding opportunities, and understanding the drivers of zooplankton, especially high quality prey, is a key step to understanding whale use of the area.
Please stay tuned for more updates as we continue working towards the answer to these pressing questions!
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly message when we post a new blog. Just add your name and email into the subscribe box below.
References
Brodeur, R. D., Auth, T. D., & Phillips, A. J. (2019). Major shifts in pelagic micronekton and macrozooplankton community structure in an upwelling ecosystem related to an unprecedented marine heatwave. Frontiers in Marine Science, 6, 212.
Cavole, L. M., Demko, A. M., Diner, R. E., Giddings, A., Koester, I., Pagniello, C. M., … & Franks, P. J. (2016). Biological impacts of the 2013–2015 warm-water anomaly in the Northeast Pacific: winners, losers, and the future. Oceanography, 29(2), 273-285.
Joh, Y., & Di Lorenzo, E. (2017). Increasing coupling between NPGO and PDO leads to prolonged marine heatwaves in the Northeast Pacific. Geophysical Research Letters, 44(22), 11-663.
Kintisch, E. (2015). ‘The Blob’ invades Pacific, flummoxing climate experts.
Leising, A. W., Schroeder, I. D., Bograd, S. J., Abell, J., Durazo, R., Gaxiola-Castro, G., … & Warybok, P. (2015). State of the California Current 2014-15: Impacts of the Warm-Water” Blob”. California Cooperative Oceanic Fisheries Investigations Reports, 56.
Morgan, C. A., Beckman, B. R., Weitkamp, L. A., & Fresh, K. L. (2019). Recent ecosystem disturbance in the Northern California current. Fisheries, 44(10), 465-474.
NOAA Fisheries. 2015b. California Current Integrated Ecosystem Assessment (CCIEA) State of the California Current Report, 2015. NMFS Report 2. Santora, J. A., Mantua, N. J., Schroeder, I. D., Field, J. C., Hazen, E. L., Bograd, S. J., … & Forney, K. A. (2020). Habitat compression and ecosystem shifts as potential links between marine heatwave and record whale entanglements. Nature communications, 11(1), 1-12.