Sophia Kormann, NSF REU Intern in the GEMM Lab, St. Olaf College
Hello! My name is Sophia Kormann and I am an NSF REU intern this summer in the GEMM lab being mentored by PI Leigh Torres, Allison Dawn, and Clara Bird. I was introduced in last week’s blog as part of our awesome whale team (deemed “Team Protein”) working out of Port Orford. I am a rising senior at St. Olaf College where I am studying statistics and biology. One of my personal goals for this summer was to get to the bottom of what is next for me. A pretty small task if you ask me… I really want to figure out if research is the route I want to go within the intersection of these two subjects or if something else would be a better fit. When looking into internships I wanted to find something where I could analyze data and see how research works as a career first hand, but not be stuck at a desk all day. I pretty much struck gold with the GEMM lab.
This summer I get to participate in field work that involves ocean kayaking, tracking whales, and identifying zooplankton, while also conducting statistical analysis on data collected from the past two years of this decade-long project. In 2022, the TOPAZ project introduced a new sensor to the data collection procedures, the RBR concerto,which records for dissolved oxygen and temperature readings during a “cast” through the water column. My big task for the summer was to explore how temperature and dissolved oxygen affect zooplankton abundance data that were simultaneously collected via a GoPro during the 2022 and 2023 field seasons.

Figure 1. Me (!) doing my first zooplankton sampling in our kayak R/V Robustus.
My project involves modeling zooplankton abundance as a response to temperature and dissolved oxygen. The ultimate goal would be to be able to plug in the dissolved oxygen and temperature to an equation and get back an accurate prediction for the zooplankton abundance, but this is often tricky to do with data that has been collected from the field. I needed to get to the bottom of what causes a change in zooplankton abundance. After a lot of trial and error, I eventually determined that temperature and dissolved oxygen at the lowest depth of each cast has the best relationship with zooplankton abundance along the whole cast, and thus produce the most accurate predictions of zooplankton abundance from the model. I literally had to go to the bottom of the ocean to get to the bottom of the relationship.
In hindsight, these relationships make a lot of sense for the Port Orford ecosystem. Ask anyone at the field station this summer and we can all tell you that it can be VERY windy here. This abundance of wind mixed with the shallow depths of the system make for very well mixed water, which means that there is little variation in the temperature and dissolved oxygen in the entire water column from the surface to the floor (Kämpf 2017). The wind here causes an increase of upwelling, which is the process of moving surface water away from the coast and allowing for deeper water to replace it. This upwelling brings cold, nutrient dense water that is low on oxygen to the surface (Bograd 2023). Since this Port Orford ecosystem is so well mixed, the bottom is likely the most stable in terms of temperature and dissolved oxygen (Ni 2016). Therefore, it would make sense that this stability would then lead to a better prediction of zooplankton as it is less affected by other factors that could be affecting the zooplankton abundance such as wind speed, land temperature, turbidity and other variables that we did not take into account while modeling.
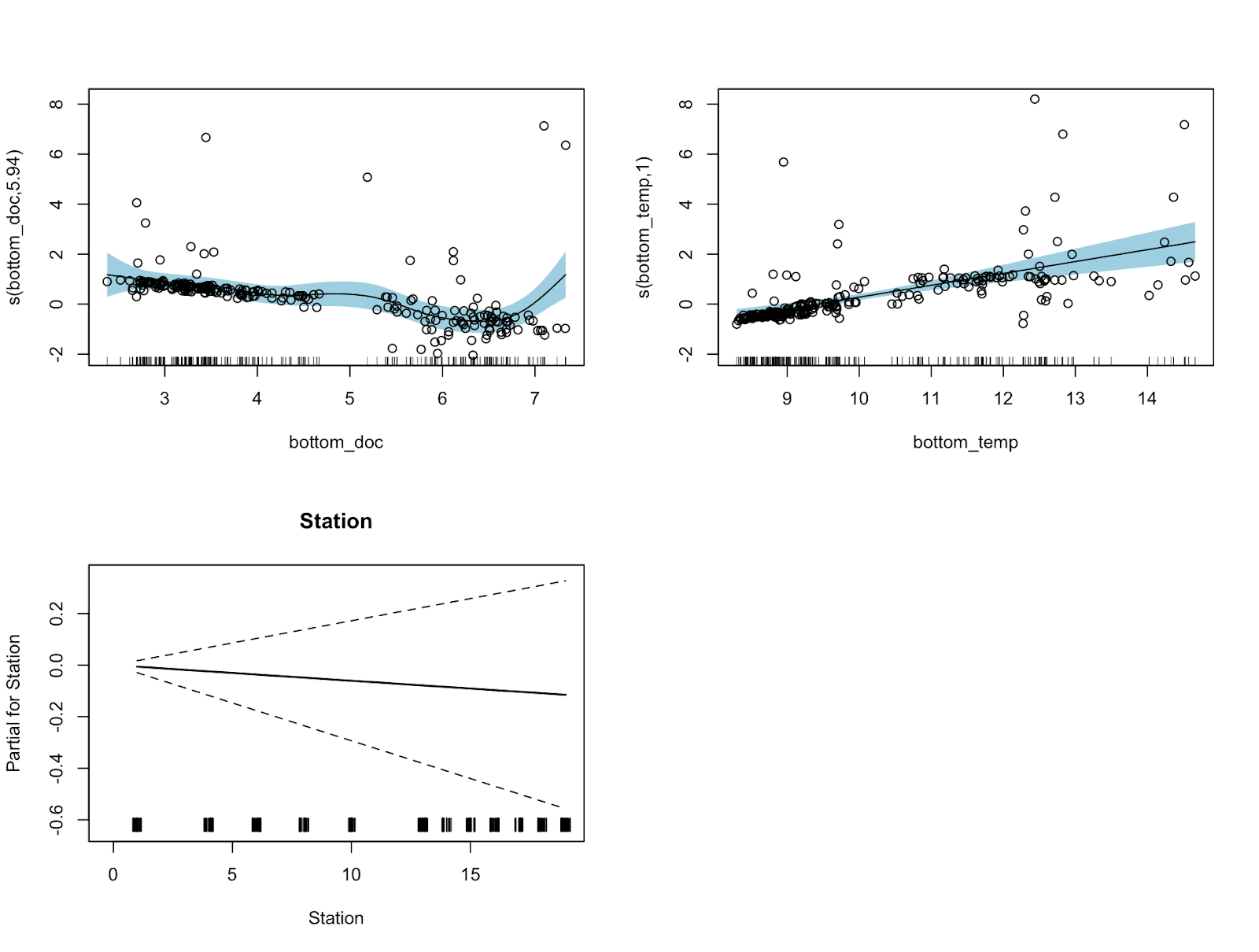
Figure 2. Functional response curves produced from a general additive model for zooplankton abundance in response to bottom dissolved oxygen (top left), bottom temperature (top right), and station (bottom).
Table 1. Zooplankton abundance is significantly affected by bottom dissolved oxygen and bottom temperature.

At this point during my summer. I have made a lot of progress in completing the data analysis and I also have made a lot of progress in getting to the bottom of “what’s next?” for me. Thankfully, this effort did not involve going to the bottom of the ocean, although I aced my kayak safety and basic life safety training since being here, so I would definitely be able to self-rescue even if I did end up there. Anyhow, one thing that helped me with this process is that I had the privilege of attending the Decadal Celebration for the TOPAZ/JASPER project. I got the chance to interact with so many people that had been in my exact place as an intern on this project over the last nine years. We discussed gap years, masters programs, and just got to hear about so many different pathways to current roles. There truly is no one “right way” to go from here.
This internship experience also taught me that I really enjoy sharing what I have discovered in this research. Whether answering the “What are you doing?” questions we get almost everyday from tourists while we are doing cliff work, or creating templates of my code for future researchers to use, or teaching Leigh Torres Gen-Z slang over dinner (ask her what “I’m dead” and “Let him cook” mean… she knows now!), I have found out that I love sharing information with others.

Figure 3. Me teaching the other interns and our new team lead how to analyze the GoPro footage.
Part of what has drawn me to statistics is the ability to turn a long string of data into an easily digestible graph for the general public. Being a part of this opportunity has allowed me to really figure out my interests and I have discovered a very genuine passion for making sense of the unknown through data analysis. With this experience I know I will be happy with whatever comes next for me as long as there is someone to share results with and a challenging question for me to get to the bottom of.
We have four more weeks of work for this field season which means more time on the ocean and hopefully more time with whales! I am very excited to see what the near future holds for me and what more we will be able to uncover this summer. With our community presentation in front of us, I am excited to share our summer with those in Port Orford. I also get to present my own research in our REU poster symposium. I look back on the almost six weeks that have already flown by with gratefulness for all I’ve already been able to learn and look forward to the next four weeks with excitement for what’s yet to be discovered.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly message when we post a new blog. Just add your name and email into the subscribe box below.
References
Bograd, SJ, Jacox, MG, Hazen, EL, Lovecchio, E, Montes, I, Pozo Buil, M, Shannon, LJ, Sydeman, WJ, Rykaczewski, RR (2023) Climate change impacts on eastern boundary upwelling systems. Annual Review of Marine Science 15
Kämpf, J (2017) Wind-driven overturning, mixing and upwelling in shallow water: A nonhydrostatic modeling study. Journal of Marine Science and Engineering, 5(4), 47. https://doi.org/10.3390/jmse5040047
Ni, X, Huang, D, Zeng, D, Zhang, T, Li, H, & Chen, J (2016) The impact of wind mixing on the variation of bottom dissolved oxygen off the Changjiang Estuary during summer. Journal of Marine Systems, 154, 122–130. https://doi.org/10.1016/j.jmarsys.2014.11.010














