Throughout the world, humans rely on coastal regions for shipping and commerce, fisheries, industrial development, and increasingly for the development of marine renewable energy such as wind and wave energy [1]. Nearshore environments, including the coastal waters of the Northern California Current (NCC), are therefore coupled social-ecological systems, at the intersection of human and biological productivity [2].
The NCC supports a diverse food web of ecologically and commercially important species [3]. The nearshore region of the NCC is further shaped by a rich mosaic of complex features including rocky reefs, kelp forests, and sloping sandy bottom substrate [4], creating habitat for numerous species of conservation interest, including invertebrates, fish, seabirds, and marine mammals [5]. Despite its importance, this realm poses significant challenges for vessel-based data collection, and therefore it remains relatively poorly monitored and understood.
The view from Cape Foulweather, showing the complex mosaic of nearshore habitat features. Photo: D. Barlow.
I am excited to introduce a new project focused on these important nearshore waters, in which we will be Examining Marine mammal Ecology through Region-wide Assessment of Long-term Data (EMERALD). Since 1992, standardized surveys have been conducted between San Francisco Bay, CA, and the Columbia River, OR, to monitor the abundance of marbled murrelets, a seabird of conservation concern. Each spring and summer, researchers have simultaneously been diligently documenting the locations of harbor porpoise and gray whale sightings—two iconic marine mammal species that rely on the nearshore waters of the NCC. This rich and extensive record is rare for marine mammal data, particularly in the challenging, turbulent nearshore environment. Furthermore, harbor porpoises are cryptic, making visual sampling particularly challenging, and gray whales can be sparsely distributed, yielding low sample sizes in the absence of long-term data collection.
Left: The survey team collecting data; Right: Marbled murrelet floating on the water.
For the EMERALD project, we will investigate spatial and temporal distribution patterns of harbor porpoises and gray whales in relation to fluctuations in key environmental drivers. The primary goals of the project are to (1) Identify persistent hotspots in harbor porpoise and gray whale sightings over time, and (2) Examine the environmental drivers of sighting hotspots through spatial and temporal analyses.
A harbor porpoise surfacing off the central Oregon coast. Photo: L. Torres.
From a first look at the data, we are already excited by some emerging patterns. In total, the dataset contains sightings of 6,763 harbor porpoise (mean 233 per year) and 530 gray whales (mean 18 per year). Preliminary data exploration reveals that harbor porpoise sightings increased in 2011-2012, predominantly between Cape Blanco, OR, and Cape Mendocino, CA. Gray whale sightings appear to follow an oscillating, cyclical pattern with peaks approximately every three years, with notable disruption of this pattern during the marine heatwave of 2014-2015. What are the drivers of sighting hotspots and spatial and temporal fluctuations in sighting rates? Time—and a quantitative analytical approach involving density estimation, timeseries analysis, and species distribution modeling—will tell.
A gray whale forages in kelp forest habitat over a nearshore rocky reef. Photo: T. Chandler.
I recently completed my PhD on the ecology and distribution of blue whales in New Zealand (for more information, see the OBSIDIAN project). Now, I am excited to apply the spatial analysis skills have been honing to a new study system and two new study species as I take on a new role in the GEMM Lab as a Postdoctoral Scholar. The EMERALD project will turn my focus to the nearshore waters close to home that I have grown to love over the past six years as a resident of coastal Oregon. The surveys I will be working with began before I was born, and I am truly fortunate to inherit such a rich dataset—a rare treat for a marine mammal biologist, and an exciting prospect for a statistical ecologist.
Dawn and Quin the dog, enjoying views of Oregon’s complex and important nearshore waters. Both are thrilled to remain in Oregon for the EMERALD project. Photo: R. Kaplan.
So, stay tuned for our findings as the project unfolds. In the meantime, I want express gratitude to Craig Strong of Crescent Coastal Research who has led the dedicated survey effort for the marbled murrelet monitoring program, without whom none of the data would exist. This project is funded by the Oregon Gray Whale License Plate funds, and we thank the gray whale license plate holders for their support of marine mammal research.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly alert when we make a new post! Just add your name into the subscribe box below!
References:
1. Jouffray, J.-B., Blasiak, R., Norström, A. V., Österblom, H., and Nyström, M. (2020). The Blue Acceleration: The Trajectory of Human Expansion into the Ocean. One Earth 2, 43–54.
2. Sjostrom, A.J.C., Ciannelli, L., Conway, F., and Wakefield, W.W. (2021). Gathering local ecological knowledge to augment scientific and management understanding of a living coastal resource: The case of Oregon’s nearshore groundfish trawl fishery. Mar. Policy 131, 104617.
3. Bograd, S.J., Schroeder, I., Sarkar, N., Qiu, X., Sydeman, W.J., and Schwing, F.B. (2009). Phenology of coastal upwelling in the California Current. Geophys. Res. Lett. 36, 1–5.
4. Romsos, G., Goldfinger, C., Robison, R., Milstein, R., Chaytor, J., and Wakefield, W. (2007). Development of a regional seafloor surficial geologic habitat map for the continental margins of Oregon and Washington, USA. Mapp. Seafloor Habitat Charact. Geol. Assoc. Canada, Spec. Pap., 219–243.
5. Oregon Department of Fish and Wildlife (2016). Oregon Nearshore Strategy. Available at: https://oregonconservationstrategy.org/oregon-nearshore-strategy/ [Accessed January 10, 2022].
About 10 months have passed since I started working on OPAL, a project that aims to identify the co-occurrence between whales and fishing effort in Oregon to reduce entanglement risk. During this period, you would be surprised to know how little ecology I have actually done and how much time has been devoted to data processing! I compiled several million GPS trackline positions, processed hundreds of marine mammal observations, wrote several thousand lines of R code, downloaded and extracted a couple Gb of environmental data… before finally reaching the modeling phase of the OPAL project. And with it, finally comes the time to look more closely at the ecology and behavior of my species of interest. While the previous steps of the project were pretty much devoid of ecological reasoning, the literature homework now comes in handy to guide my choices regarding habitat use models, such as selecting environmental predictors of whale occurrence, deciding on what seasons should be modeled, and choosing the spatio-temporal scale at which the data should be aggregated.
Whale diversity on the US west coast
The productive waters off the US west coast host a great diversity of cetaceans. Eight species of baleen whales are reported to occur there by NOAA fisheries: blue whales, Bryde’s whales, fin whales, gray whales, humpback whales, minke whales, North Pacific right whales and sei whales. Among them, no less than five are listed as Endangered under the Endangered Species Act. Whether they are only passing by or spending months feeding in the region, the timing and location where these animals are observed varies greatly by species and by population.
During the 113 hours of aerial survey effort and 264 hours of boat-based search conducted for the OPAL project, 563 groups of baleen whales have been observed to-date (up to mid-May 2021 to be exact… more data coming soon!). Among the observations where animals could be identified to the species level, humpback whales are preponderant, as they represent about half of the whale groups observed (n = 293). Blue (n = 41) and gray whales (n = 46) come next, the latter being observed in more nearshore waters. Finally, a few fin whale groups were observed (n = 28). The other baleen whale species reported by NOAA in the US west coast species list were very rarely or not observed at all during OPAL surveys.
The OPAL aerial surveys conducted in partnership with the United States Coast Guard (USCG) were specifically designed to study whales occurring on the continental shelf along the coast of Oregon. Hence, most of this survey effort is located in waters from 800 m to 30 m deep, which may explain the relatively low number of gray whales detected. Indeed, gray whales observed in Oregon may either be migrating along the coast to and from their breeding grounds in Baja California, or be part of the small Pacific Coast Feeding Group that forage in Oregon nearshore and shallow waters during the summer. This group of whales is one the main GEMM lab’s research focus, being at the core of no less than three ongoing research projects: AMBER, GRANITE, and TOPAZ.
So today, let’s turn our eyes to the sea horizon and talk about some other members of the baleen whale community: rorquals. Conveniently, the three species of baleen whales (gray whales aside) most commonly observed during OPAL surveys are all part of the rorqual family, a.k.a Balaenopteridae: humpback whales, blue whales and fin whales (Figure 1). They are morphologically characterized by the pleated throat grooves that allow them to engulf large quantities of food and water, for instance when lunge-feeding. Known cases of hybridization between these three species demonstrate their close relatedness (Jefferson et al., 2021). They all have worldwide distributions and display unequally understood migratory behaviors, seasonally traveling between warm tropical breeding grounds and temperate-polar feeding grounds. They occur in great numbers in productive waters such as the upwelling system of the California Current.
The three accomplices
Figure 1: Aerial view of three rorquals species: a humpback whale (left), a fin whale (center), and a blue whale (right). Photo credit: Leigh Torres and Craig Hayslip. Photos taken off the Oregon coast under NOAA/NMFS permit during USCG helicopter flights conducted as part of the OPAL project
Humpback whales (Megaptera novaeangliae) are easily differentiated from other rorquals because of their long pectoral fins (up to one third of their body length!), which inspired their scientific name, Megaptera, « big-winged » (Figure 1). Individuals observed in Oregon mostly belong to a mix of two Distinct Population Segments (DPS): the threatened Mexico and endangered Central American DPS. Although humpback whales from different DPS do not show any morphological differences, they are genetically distinct because they have been mating separately in distinct breeding grounds for generations and generations. This genetic differentiation has great implications in terms of conservation since the Central American DPS is recovering at a lesser rate than the Mexican and is therefore subject to different management measures (recovery plan, monitoring plan, designated critical habitats). Humpback whales migrate and feed off the US west coast, with a peak in abundance in the mid to late summer. Compared to other rorquals that are found in the open ocean, humpback whales are mostly observed on the continental shelf (Becker et al., 2019). They are considered to have a relatively generalist diet, as they feed on a mix of krill (Euphausiids) and fishes (e.g. anchovy, sardines) and are capable of switching their feeding behavior depending on relative prey availability (Fleming, Clark, Calambokidis, & Barlow, 2016; Fossette et al., 2017).
Blue whales (Balaenoptera musculus) are the largest animals ever known (max length 33 m, Jefferson et al., 2008), and sadly the most at risk of global extinction among our three species of interest (listed as « endangered » in the IUCN red list). They have a distinctive mottled blue and light gray skin, a slender body and a broad U-shaped head (or as some say « like a gothic arch », Figure 1). Blue whales tend to be open ocean animals, but they regroup seasonally to feed in highly productive nearshore areas such as the Southern California Bight (Becker et al. 2019, Abrahms et al. 2019). Blue whales migrating or feeding along the US west coast belong to the Eastern North Pacific stock and are subject to great research and conservation efforts. Contrary to their other rorqual counterparts, blue whales are quite picky eaters, as they exclusively feed on krill. This difference in diet leads to resource partitioning facilitating rorqual coexistence in the California Current (Fossette et al., 2017). These differences in feeding strategies have important implications for designing predictive models of habitat use.
Fin whales (Balaenoptera physalus) are nicknamed « greyhounds of the sea » due to their exceptional swim speed (max 46 km/h). They are a little smaller than blue whales (max length 27 m, Jefferson, Webber, & Pitman, 2008) but share a similar sleek and streamlined shape. Their coloration is their most distinctive feature: the left lower jaw being mostly dark while the right is white. V-shaped light-gray « chevrons » color their back, behind the head (Figure 1). The California/Oregon/Washington is one of the three stocks recognized in the North Pacific (NOAA Fisheries, 2018). Within this region, there is genetic evidence for a geographic separation north and south of Point Conception, CA (Archer et al., 2013). Like other rorquals, they are migratory, but their seasonal distribution is relatively less well understood as they appear to spend a lot of time in open oceans. For instance, a meta-analysis for the North Pacific found little evidence for fin whales using distinct calving areas (Mizroch, Rice, Zwiefelhofer, Waite, & Perryman, 2009). In the California Current System, satellite tracking has provided great insights into their space-use patterns. In the Southern California Bight, fin whales show year-round residency and seasonal shifts in habitat use as they move further offshore and north during the spring/summer (Scales et al., 2017). The Northern California Current offshore waters appeared to be used during the summer months by the whales tagged in the Southern California Bight. Yet, fin whales are observed year-round in Oregon (NOAA Fisheries, 2018).
Towards predictive models of rorqual distribution
Enough observations have now been collected as part of the OPAL project to be able to model the habitat use of some of these rorqual species. Based on 12 topographic (i.e., depth, slope, distance to canyons) and physical variables (temperature, chlorophyll-a, water column stratification, etc.), I have made my first attempt at predicting seasonal distribution patterns of humpback whales and blue whales in Oregon. These models will be improved in the coming months, with more data pouring in and refined parametrizations, but they already bring insights into the shared habitat use patterns of these species, as well as their specificities.
Across multiple cross-validations of the species-specific models, sea surface temperature, sea surface height and depth were recurrently selected among the most important variables influencing both humpback and blue whale distributions. Predicted densities of blue whales were relatively higher at less than 40 fathoms compared to humpback whales, although both species’ hotspots were located outside this newly implemented seasonal fishing limit (Figure 2). Higher densities were generally predicted off Newport and Port Orford, and north of North Bend.
Figure 2: Predicted densities of humpback and blue whales during the month of September 2018, 2019, and 2020 in Oregon waters (OPAL project). Core areas of use (predicted densities in the top 25%) are represented, with darker shades of blue and orange showing higher predicted densities. Dashed lines represent the tracklines followed by USCG monthly aerial surveys. The black line represents the 40 fathom isobath. Grey boxes overlayed on predictions delineate the areas of extrapolation where environmental conditions are non-analogous to the conditions in which the models were trained. Disclaimer: these model outputs are preliminary and should be interpreted with caution.
Once our rorqual models are finalized, we will work with our partners at the Oregon Department of Fisheries and Wildlife to overlay predicted whale hotspots with areas of high crab pot densities. This overlap analysis will help us understand the times and places where co-occurrence of suitable whale habitat and fishing activities put whales at risk of entanglement.
Becker, E. A., Forney, K. A., Redfern, J. V, Barlow, J., Jacox, M. G., Roberts, J. J., & Palacios, D. M. (2019). Predicting cetacean abundance and distribution in a changing climate. Diversity and Distributions, 25(4), 626–643. https://doi.org/10.1111/ddi.12867
Fleming, A. H., Clark, C. T., Calambokidis, J., & Barlow, J. (2016). Humpback whale diets respond to variance in ocean climate and ecosystem conditions in the California Current. Global Change Biology, 22, 1214–1224. https://doi.org/10.1111/gcb.13171
Fossette, S., Abrahms, B., Hazen, E. L., Bograd, S. J., Zilliacus, K. M., Calambokidis, J., … Croll, D. A. (2017). Resource partitioning facilitates coexistence in sympatric cetaceans in the California Current. Ecology and Evolution, 7, 9085–9097. https://doi.org/10.1002/ece3.3409
Jefferson, T. A., Palacios, D. M., Clambokidis, J., Baker, S. C., Hayslip, C. E., Jones, P. A., … Schulman-Janiger, A. (2021). Sightings and Satellite Tracking of a Blue / Fin Whale Hybrid in its Wintering and Summering Ranges in the Eastern North Pacific. Advances in Oceanography & Marine Biology, 2(4), 1–9. https://doi.org/10.33552/AOMB.2021.02.000545
Jefferson, T. A., Webber, M. A., & Pitman, R. L. (2008). Marine Mammals of the World. A comprehensive guide to their identification. Elsevier, London, UK.
Mizroch, S. A., Rice, D. W., Zwiefelhofer, D., Waite, J., & Perryman, W. L. (2009). Distribution and movements of fin whales in the North Pacific Ocean. Mammal Review, 39(3), 193–227. https://doi.org/10.1111/j.1365-2907.2009.00147.x
NOAA Fisheries. (2018). Fin whale stock assessment report ( Balaenoptera physalus physalus ): California / Oregon / Washington Stock.
Scales, K. L., Schorr, G. S., Hazen, E. L., Bograd, S. J., Miller, P. I., Andrews, R. D., … Falcone, E. A. (2017). Should I stay or should I go? Modelling year-round habitat suitability and drivers of residency for fin whales in the California Current. Diversity and Distributions, 23(10), 1204–1215. https://doi.org/10.1111/ddi.12611
What I mean is that the vastness of the ocean is very hard to mentally visualize. When facing a conservation issue such as increased whale entanglement along the US West Coast (see OPAL project ), a tempting solution may be to suggest « let’s go see where the whales are and report their location to the fishermen?! ». But, it only takes a little calculation to realize how impractical this idea is.
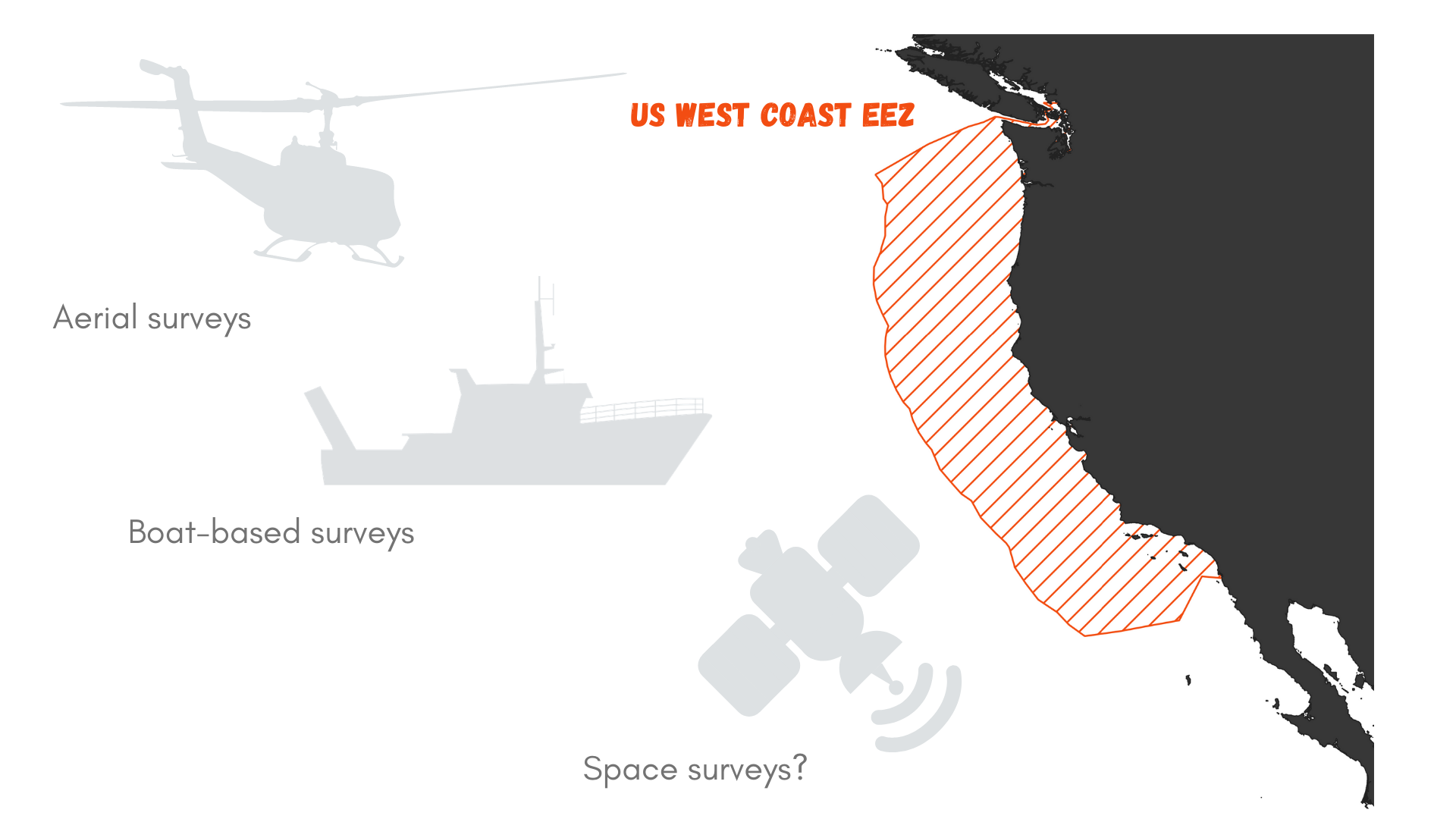
Let’s roll out the numbers. The US West Coast exclusive economic zone (EEZ) stretches from the coast out to 200 nautical miles offshore, as prescribed by the 1982 United Nations Convention on the Law of the Sea. It covers an area of 825,549 km² (Figure 1). Now, imagine that you wish to survey this area for marine mammals. Using a vessel such as the R/V Bell M. Shimada that is used for the Northern California Current Ecosystem surveys cruises (NCC cruises, see Dawn and Rachel’s last blog), we may detect whales at a distance of roughly 6 km (based on my preliminary results). This distance of detection depends on the height of the observer, hence the height of the flying bridge where she/he is standing (the observer’s height may also be accounted for, but unless she/he is a professional basket-ball player, I think it can be neglected here). The Shimada is quite a large ship and it’s flying bridge is 13 meters above the water. Two observers may survey the water on each side of the trackline.
Considering that the vessel is moving at 8 knots (~15 km/h), we may expect to be effectively surveying 180 km² per hour (6x2x15). That’s not too bad, right?
Again, perspective is the key. If we divide the West Coast EEZ surface by 180 km² we can estimate that it would take 2,752 hours to survey this entire region. With an average of 12 hours of daylight, this takes us to…
382 DAYS OF SURVEY, searching for marine mammals over the US West Coast. Considering that observations cannot be undertaken on days with bad weather (fog, heavy rain, strong winds…), it might take more than a year and a half to complete the survey! And what would the marine mammals have done in the meantime? Move…
This little math exercise proves that exhaustively searching for the needle in the haystack from a vessel is not the way to go if we are to describe whale distribution and help mitigate the risk of entanglement. And using another platform of observation is not necessarily the solution. The OPAL project has relied on a great collaboration with the United States Coast Guard to survey Oregon waters. The USCG helicopters travel fast compared to a vessel, about 90 knots (167 km/h). As a result, more ground is covered but the speed at which it is traveling prevents the observer from detecting whales that are very far away. Based on the last analysis I ran for the OPAL project, whales are usually detected up to 3 km from the helicopter (only 5 % of sightings exceed that distance). In addition, the helicopter generally only has capacity for one observer at a time.
If we replicate the survey time calculation from above for the USCG helicopter, we realize that even with a fast-moving aerial survey platform it would still take 137 days to cover the West Coast EEZ.
Figure 1. What is the best survey method to document marine mammal occurrence in the US West Coast Exclusive Economic zone (EEZ)?
First, we can model and extrapolate. This approach is the path we are taking with the OPAL project: we survey Oregon waters in 4 different areas along the coast each month, then model observed whale densities as a function of topographic and oceanographic variables, and then predict whale probability of presence over the entire region. These predictions are based on the assumption that our survey design effectively sampled the variety of environmental conditions experienced by whales over the study region, which it certainly did considering that all sites are surveyed year-round.
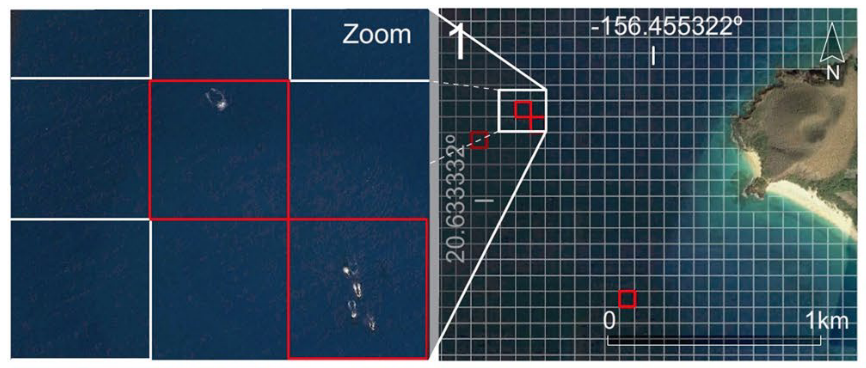
An alternative approach that has been recently discussed in the GEMM Llab, is the use of satellite images to detect whales along the coast. A communication entitled « The Potential of Satellite Imagery for Surveying Whales » was published last month in the Sensors Journal (Höschle et al., 2021) and presents the opportunities offered by this relatively new technology. The WorldView-3 satellite, owned by the company Digitalglobe and launched in 2016, has made it possible to commercialize imagery with a resolution never reached before, of the order of 30 cm per pixel. These very high resolution (VHR) satellite images make it possible to identify several species of large whales (Cubaynes et al. al., 2019) and to estimate their density (Bamford et al., 2020). Furthermore, machine learning algorithms, such as Neural Networks, have proved quite efficient at automatically detecting whales in satellite images (Guirado et al., 2019, Figure 2). While several new ultra-high resolution imaging satellites are expected to be launched in 2021 (by Maxar Technologies and Airbus), this “remote” approach looks like a promising avenue to detect whales over vast regions while drinking a cup of coffee at the office.
Figure 2. Illustration of a whale detection algorithm working on a gridded satellite image (DigitalGlobe). Source: Guirado et al., 2019.
But like any other data collection method, satellites have their drawbacks. We recently discovered that these VHR satellites are routinely switched off while passing above the ocean. Specific inquiries would need to be made to acquire data over our study areas, which would be at great expense. One of the cheapest provider I found is the Soar platform, that provides images at 50 cm resolution in partnership with the Chinese Aerospace Science and Technology Corporation. They advertise daily images anywhere on earth at $10 USD per km². This might sound cheap at first glance, but circling back to our US West Coast EEZ area calculations, we estimate that surveying this region entirely with satellite imagery would cost more than $8 million USD.
Yet, we have to look forward. The use of satellite imagery is likely to broaden and increase in the coming years, with a possible decrease in cost. Quoting Höschle et al. (2021) ‘To protect our world’s oceans, we need a global effort and we need to create opportunities for that to happen’.
Will satellites soon save whales?
References
Bamford, C. C. G. et al. A comparison of baleen whale density estimates derived from overlapping satellite imagery and a shipborne survey. Sci. Rep. 10, 1–12 (2020).
Cubaynes, H. C., Fretwell, P. T., Bamford, C., Gerrish, L. & Jackson, J. A. Whales from space: Four mysticete species described using new VHR satellite imagery. Mar. Mammal Sci. 35, 466–491 (2019).
Guirado, E., Tabik, S., Rivas, M. L., Alcaraz-Segura, D. & Herrera, F. Whale counting in satellite and aerial images with deep learning. Sci. Rep. 9, 1–12 (2019).
Höschle, C., Cubaynes, H. C., Clarke, P. J., Humphries, G. & Borowicz, A. The potential of satellite imagery for surveying whales. Sensors 21, 1–6 (2021).
1PhD student, Oregon State University College of Earth, Ocean, and Atmospheric Sciences and Department of Fisheries and Wildlife, Geospatial Ecology of Marine Megafauna Lab
“Hurry up and wait.” A familiar phrase to anyone who has conducted field research. A flurry of preparations, followed by a waiting game—waiting for the weather, waiting for the right conditions, waiting for unforeseen hiccups to be resolved. We do our best to minimize unknowns and unexpected challenges, but there is always uncertainty associated with any endeavor to collect data at sea. We cannot control the whims of the ocean; only respond as best we can.
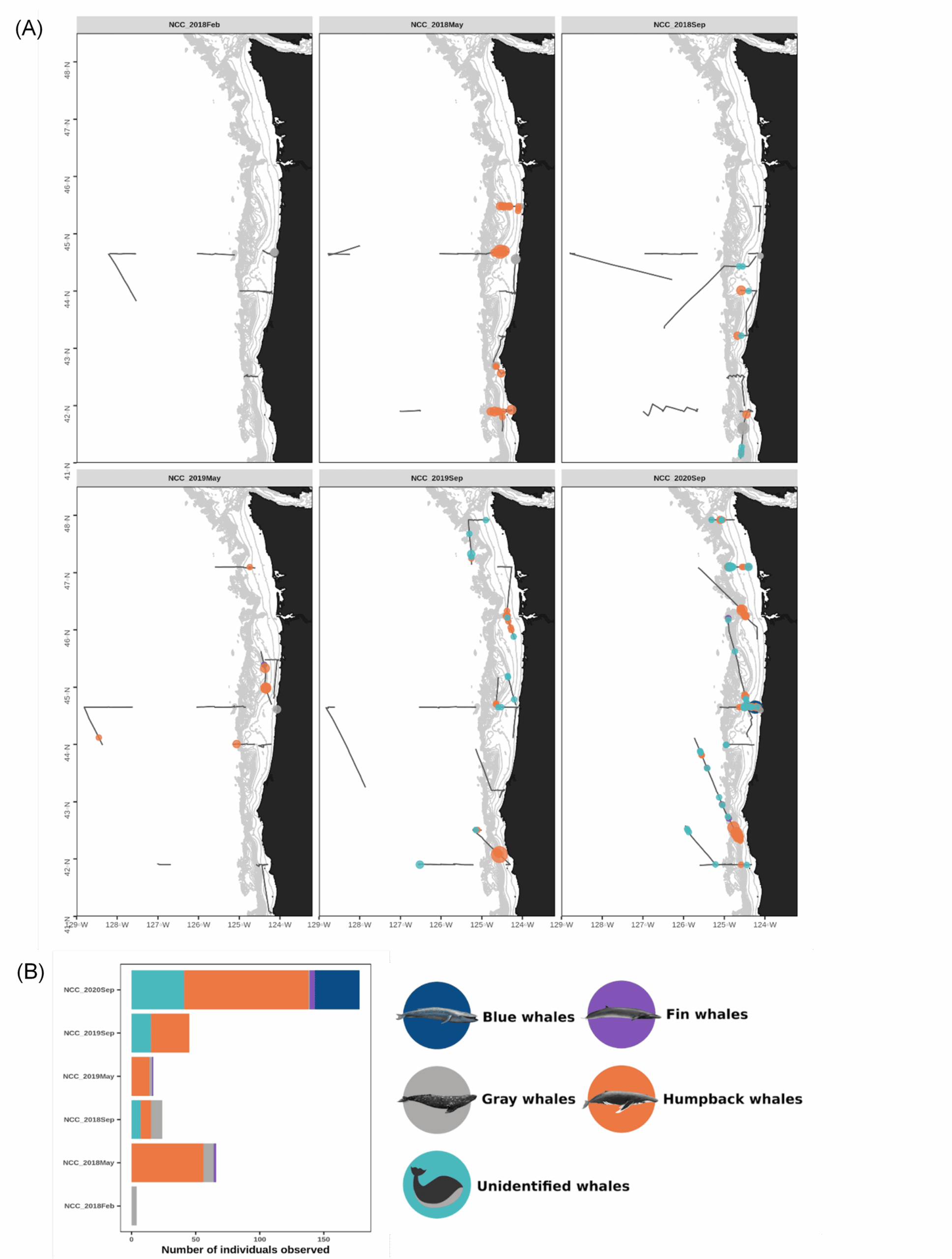
On 15 February 2021, we were scheduled to board the NOAA Ship Bell M. Shimada as marine mammal observers for the Northern California Current (NCC) ecosystem survey, a recurring research cruise that takes place several times each year. The GEMM Lab has participated in this multidisciplinary data collection effort since 2018, and we are amassing a rich dataset of marine mammal distribution in the region that is incorporated into the OPAL project. February is the middle of wintertime in the North Pacific, making survey conditions challenging. For an illustration of this, look no further than at the distribution of sightings made during the February 2018 cruise (Fig. 1), when rough sea conditions meant only a few whales were spotted.
Figure 1. (A) Map of marine mammal survey effort (gray tracklines) and baleen whale sightings recorded onboard the NOAA ship R/V Shimada during each of the NCC research cruises to-date and (B) number of individuals sighted per cruise since 2018. Note the amount of survey effort conducted in February 2018 (top left panel) compared to the very low number of whales sighted. Data summary and figures courtesy of Solene Derville.
Now, this is February 2021and the world is still in the midst of navigating the global coronavirus pandemic that has affected every aspect of our lives. The September 2020 NCC cruise was the first NOAA fisheries cruise to set sail since the pandemic began, and all scientists and crew followed a strict shelter-in-place protocol among other COVID risk mitigation measures. Similarly, we sheltered in place in preparation for the February 2021 cruise. But here’s where the weather comes in yet again. Not only did we have to worry about winter weather at sea, but the inclement conditions across the country meant our COVID tests were delayed in transit—and we could not board the ship until everyone tested negative. By the time our results were in, the marine forecast was foreboding, and the Captain determined that the weather window for our planned return to port had closed.
So, we are still on shore. The ship never left the dock, and NCC February 2021 will go on the record as “NAs” rather than sightings of marine mammal presence or absence. So it goes. We can dedicate all our energy to studying the ocean and these spectacularly dynamic systems, but we cannot control them. It is an important and humbling reminder. But as we have continued to learn over the past year, there are always silver linings to be found.
Even though we never made it to the ship, it turns out there’s a lot you can get done onshore. Dawn has sailed on several NCC cruises before, and one of the goals this time was to train Rachel for her first stint at marine mammal survey work. This began at Dawn’s house in Newport, where we sheltered in place together for the week prior to our departure date.
We walked through the iPad program we use to enter data, looked through field guides, and talked over how to respond in different scenarios we might encounter while surveying for marine mammals at sea. We also joined Solene, a postdoc working on the OPAL project, for a Zoom meeting to edit the distance sampling protocol document. It was great training to discuss the finer points of data collection together, with respect to how that data will ultimately be worked into our species distribution models.
The February NCC cruise is famously rough, and a tough time to sight whales (Fig. 1). This low sighting rate arises from a combination of factors: baleen whales typically spend the winter months on their breeding grounds in lower latitudes so their density in Oregon waters is lower, and the notorious winter sea state makes sighting conditions difficult. Solene signed off our Zoom call with, “Go collect that high-quality absence data, girls!” It was a good reminder that not seeing whales is just as important scientifically as seeing them—though sometimes, of course, it’s not possible to even get out where you can’t see them. Furthermore, all absence data is not created equal. The quality of the absence data we can collect deteriorates along with the weather conditions. When we ultimately use these survey data to fuel species distribution models, it’s important to account for our confidence in the periods with no whale sightings.
In addition to the training we were able to conduct on land, the biggest silver lining came just from sheltering in place together. We had only met over Zoom previously, and spending this time together gave us the opportunity to get to know each other in real life and become friends. The week involved a lot of fabulous cooking, rainy walks, and an ungodly number of peanut butter cups. Even though the cruise couldn’t happen, it was such a rich week. The NCC cruises take place several times each year, and the next one is scheduled for May 2021. We’ll keep our fingers crossed for fair winds and negative COVID tests in May!
Figure 2. Dawn’s dog Quin was a great shelter in place buddy. She was not sad that the cruise was canceled.
As a GEMM lab post-doc working on the OPAL project, my main goal for 2021 will be to produce accurate predictive models of baleen whale distribution off the Oregon coast to reduce entanglement risk. For the past months, I have been compiling, cleaning, and processing about two years of data collected by Leigh Torres and Craig Hayslip during monthly repeat surveys conducted onboard United States Coast Guard (USCG) helicopters. These standardized surveys record where and when whales are observed off the Oregon coast. These presence and absence data may now be modeled in relation to habitat, while accounting for effort and detection (as several parameters, such as weather and sea state, can affect the capacity of observers to detect whales at the surface). Considering that several baleen whale species (namely, humpback, fin, blue and gray whales) are known to feed in the area, prey availability is expected to be a major driver of their distribution.
As prey distribution data are frequently the lacking component in the habitat model equation, whale ecologists often resort to using environmental proxies. Variables such as topography (e.g., the depth or slope of the seafloor), water physical and chemical characteristics (e.g., temperature, salinity, oxygen concentration) or ocean circulation (e.g., currents, turbulence) have proved to be good predictors for fish or krill distribution, and in turn potential predictors for whale suitable habitats. In my search for such environmental variables to be tested in our future OPAL models, I have been focusing my research on a fascinating ocean feature: sea height.
Sea height varies both temporally and spatially under the influence of multiple factors, from internal mass of the solid Earth to the orbital revolution of the moon. After reading this blog you will realize that the flatness of the horizon at sea is a deceiving perspective (Figure 1) …
Figure 1: Flat? Really? (source: Pixabay)
Gravity and the geoid
We all know of Newton’s s discovery of gravity: the attraction force exerted by any object with a given mass on its surroundings. Yet, it is puzzling to think that the rate of acceleration of the apple falling on Newton’s head would have been different if Newton had been anywhere else on Earth.
Why is that and what does it have to do with sea height? On Earth, the standard gravity g is set at 9.80665 m/s2. This constant is called a “standard” because in fact, gravity varies at the surface of our planet, even if estimated at a fixed altitude. Indeed, as gravity is caused by mass, any change in relief or rock composition results in a change in gravity. For instance, magmatic activity in the upper mantle of the Earth and the crust causes a change in rock density and results in a change in gravity measured at the surface.
Gravity therefore is the first reason why the ocean surface is not flat. Gravity shapes an irregular surface called the “geoid”. This hypothetical ocean surface has equal gravitational potential anywhere on Earth and differs from the ellipsoid of reference by as much as 100 m! So to the question whether Earth is round or flat, I would say it is potato shaped (Figure 2)!
Figure 2: Exaggerated view of the gravitational potential of Earth. View a video animation here. (credit: European Space Agency)
The geoid is an essential reference for understanding ocean currents and monitoring changes in sea-level. Hypothetically, if ocean water had equal density everywhere and at any depth, the sea surface should match with the geoid… but that’s not the case. Let’s see why.
Ocean dynamic topography
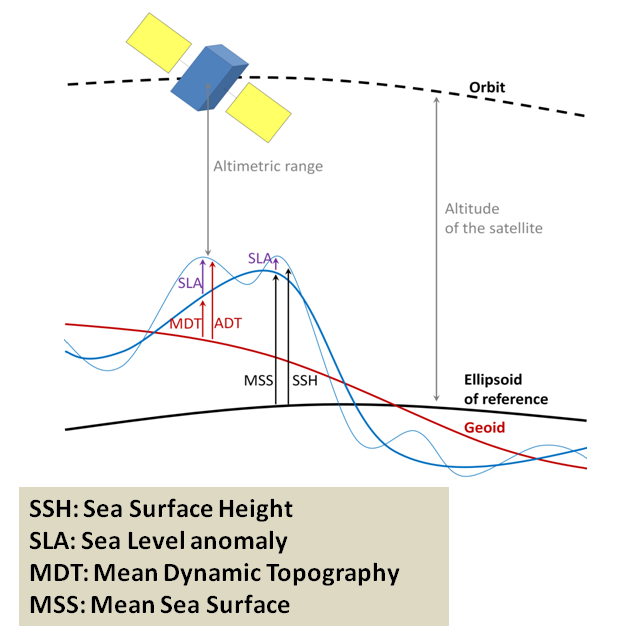
Not unlike the hills and valleys covering landscapes, the ocean surface also has its highs and lows. Except that in the ocean, the surface topography is ever changing. Sea surface height (SSH) measures the average height difference between the observed sea level and the ellipsoid of reference (Figure 3). SSH is mostly affected by ocean circulation and may vary by as much as ±1 m. Indeed, just like the rocks inside the Earth, the water in the ocean varies in density. The vertical and horizontal physical structuring of the ocean was extensively discussed by Dawn last November while she was preparing for her PhD Qualifying Exams. Temperature clearly is at the core of the processes. As thermal expansion increases the space between warming water particles, the volume of a given amount of liquid water increases with increasing temperature. Warmer waters therefore take up “more space” than cooler waters, resulting in an elevated SSH.
Figure 3: Overview of the different fields used in altimetry (credit: CLS, https://duacs.cls.fr/)
SSH may therefore be used as an indicator of oceanographic phenomena such as upwellings, where warm surface waters are replaced by deep, cooler, and nutrient-rich waters moving upwards. The California Current that moves southwards along the North American coast is known as one of the world’s major currents affiliated with strong upwelling zones, which often triggers increased biological productivity. Several studies conducted in the California Current system have found a link between the variations in SSH and whale abundance or foraging activity (Abrahms et al. 2019; Pardo et al. 2015; Becker et al. 2016; Hazen et al. 2016).
SSH is measured by altimeter satellites and is made freely available by the European Space Agency and the US National Aeronautics and Space Administration. Lucky me! Numerous variables are derived from SSH, as shown in Figure 3. Among other things, I was able to download the daily maps of Sea Surface Height Anomaly (SSHa, also referred to as Sea Level Anomaly: SLA) over the Oregon coast from February 2019 to December 2020. SSHa is the difference between observed SSH at a specific time and place from the mean SSH field of reference calculated over a long period of time. Negative values of SSHa potentially suggest upwellings of cooler waters that could be associated with higher prey availability. Figure 4 shows an example of environmental data mining as I try to match SSHa with whale observations made during OPAL surveys. Figure 4B suggests increased whale occurrence where/when SSHa is lower.
Figure 4: Preliminary exploration of the relationship between sea surface height anomaly (SSHa) and baleen whales (blue, fin, humpback, unidentified) observed during OPAL surveys off Oregon, USA, between February 2019 and December 2020. A) Example covering 3 months of survey during summer 2019. Sightings were grouped over 5-km segments of surveyed trackline and segments with at least one sighting were mapped with colored circles. Dotted grey lines are the repeated survey tracklines for each of the labeled study areas (NB = North Bend). Sightings are symbolized by area (color) and group size (circle size). Monthly averages of SSHa are represented with a colored gradient. B) Monthly averages of SSHa measured over 5-km segments where whales were detected (presence) or not (absence).
Although encouraging, these preliminary insights are just the tip of the modeling iceberg. Many more testing and modeling steps will be required to determine confounding factors and relevant spatio-temporal scales at which these oceanographic variables may be influencing whale distribution off the Oregon coast. I am only at the start of a long road…
References
Abrahms, Briana, Heather Welch, Stephanie Brodie, Michael G. Jacox, Elizabeth A. Becker, Steven J. Bograd, Ladd M. Irvine, Daniel M. Palacios, Bruce R. Mate, and Elliott L. Hazen. 2019. “Dynamic Ensemble Models to Predict Distributions and Anthropogenic Risk Exposure for Highly Mobile Species.” Diversity and Distributions, no. December 2018: 1–12. https://doi.org/10.1111/ddi.12940.
Becker, Elizabeth, Karin Forney, Paul Fiedler, Jay Barlow, Susan Chivers, Christopher Edwards, Andrew Moore, and Jessica Redfern. 2016. “Moving Towards Dynamic Ocean Management: How Well Do Modeled Ocean Products Predict Species Distributions?” Remote Sensing 8 (2): 149. https://doi.org/10.3390/rs8020149.
Hazen, Elliott L, Daniel M Palacios, Karin A Forney, Evan A Howell, Elizabeth Becker, Aimee L Hoover, Ladd Irvine, et al. 2016. “WhaleWatch : A Dynamic Management Tool for Predicting Blue Whale Density in the California Current.” Journal of Applied Ecology 54 (5): 1415–28. https://doi.org/10.1111/1365-2664.12820.
Pardo, Mario A., Tim Gerrodette, Emilio Beier, Diane Gendron, Karin A. Forney, Susan J. Chivers, Jay Barlow, and Daniel M. Palacios. 2015. “Inferring Cetacean Population Densities from the Absolute Dynamic Topography of the Ocean in a Hierarchical Bayesian Framework.” PLOS One 10 (3): 1–23. https://doi.org/10.1371/journal.pone.0120727.
Clara and I have just returned from ten fruitful days at sea aboard NOAA Ship Bell M. Shimada as part of the Northern California Current (NCC) ecosystem survey. We surveyed between Crescent City, California and La Push, Washington, collecting data on oceanography, phytoplankton, zooplankton, and marine mammals (Fig. 1). This year represents the third year I have participated in these NCC cruises, which I have come to cherish. I have become increasingly confident in my marine mammal observation and species identification skills, and I have become more accepting of the things out of my control – the weather, the sea state, the many sightings of “unidentified whale species”. Careful planning and preparation are critical, and yet out at sea we are ultimately at the whim of the powerful Pacific Ocean. Another aspect of the NCC cruises that I treasure is the time spent with members of the science team from other disciplines. The chatter about water column features, musings about plankton species composition, and discussions about what drives marine mammal distribution present lively learning opportunities throughout the cruise. Our concurrent data collection efforts and ongoing conversations allow us to piece together a comprehensive picture of this dynamic NCC ecosystem, and foster a collaborative research environment.
Figure 1. Data collection effort for the NCC September 2020 cruise, between Crescent City, CA, and La Push, WA. Red points represent oceanographic sampling stations, and black lines show the track of the research vessel during marine mammal survey effort.
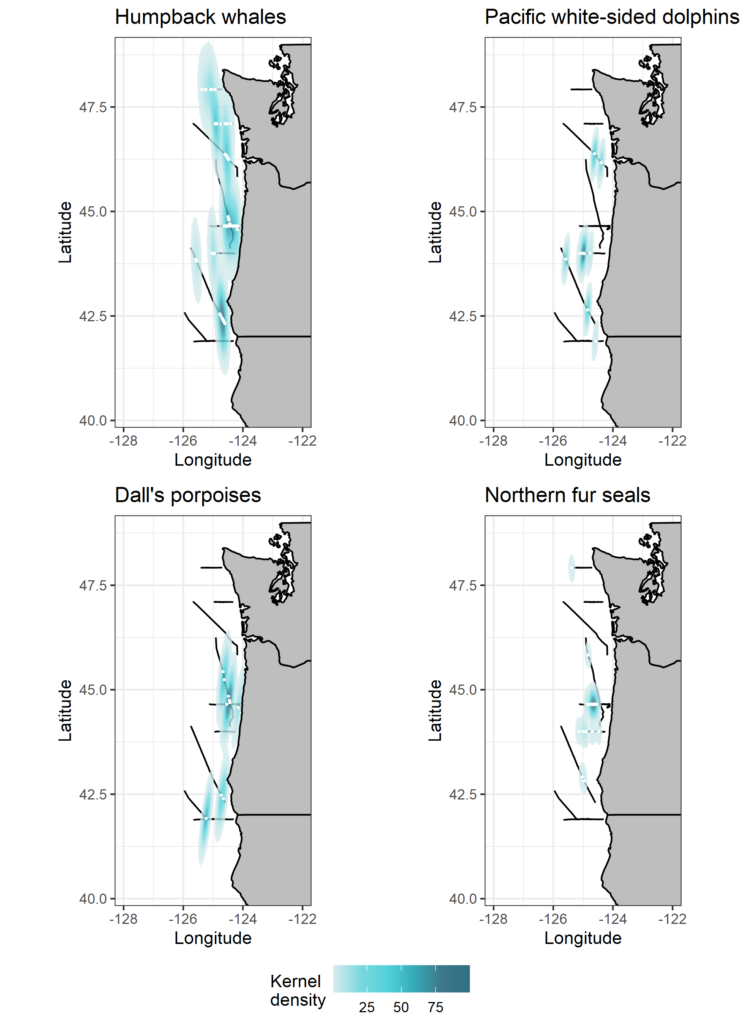
Every time I head to sea, I am reminded of the patchy distribution of resources in the vast and dynamic marine environment. On this recent cruise we documented a stark contrast between expansive stretches of warm, blue, stratified, and seemingly empty ocean and areas that were plankton-rich and supported multi-species feeding frenzies that had marine mammal observers like me scrambling to keep track of everything. This year, we were greeted by dozens of blue and humpback whales in the productive waters off Newport, Oregon. Off Crescent City, California, the water was very warm, the plankton community was dominated by gelatinous species like pyrosomes, salps, and other jellies, and the marine mammals were virtually absent except for a few groups of common dolphins. To the north, the plume of water flowing from the Columbia River created a front between water masses, where we found ourselves in the midst of pacific white-sided dolphins, northern right whale dolphins, and humpback whales. These observations highlight the strength of ecosystem-scale and multi-disciplinary data collection efforts such as the NCC surveys. By drawing together information on physical oceanography, primary productivity, zooplankton community composition and abundance, and marine predator distribution, we can gain a nearly comprehensive picture of the dynamics within the NCC over a broad spatial scale.
Northern fur seal
Humpback whales
Common dolphins
Humpback whale breaching
Blue whale
Risso’s dolphins
Pacific white-sided dolphins
A few images of marine mammals of the NCC. Click a photo to explore gallery. All photos by Dawn Barlow.
This year, the marine mammals delivered and kept us observers busy. We lucked out with good survey conditions and observed many different species throughout the NCC (Table 1, Fig. 2).
Table 1. Summary of all marine mammal sightings from the NCC September 2020 cruise.
Figure 2. Maps showing kernel densities of four frequently observed and widely distributed species seen during the cruise. Black lines show the track of the research vessel during marine mammal survey effort, white points represent sighting locations, and colors show kernel density estimates weighted by group size at each sighting.
This year’s NCC cruise was unique. We went to sea as a global pandemic, wildfires, and political tensions continue to strain this country and our communities. This cruise was the first NOAA Fisheries cruise to set sail since the start of the pandemic. Our team of scientists and the ship’s crew went to great lengths to make it possible, including a seven-day shelter-in-place period and COVID-19 tests prior to cruise departure. As a result of these extra challenges and preparations, I think we were all especially grateful to be on the water, collecting data. At-sea fieldwork is always challenging, but morale was up, spirits were high, and laughs were frequent despite smiles being concealed by our masks. I am grateful for the opportunity to participate in this ongoing valuable data collection effort, and to be part of this team. Thanks to all who made it such a memorable cruise.
Figure 3. The NCC September 2020 science team at the end of a successful research cruise! Fieldwork in the time of COVID-19 presents many logistical challenges, but this team rose to the occasion and completed a safe and fruitful survey despite the circumstances.
Just like that, I have wrapped up year 1 of my PhD in Wildlife Science. For my PhD, I am investigating the ecology and distribution of blue whales in New Zealand across multiple spatial and temporal scales. In a region where blue whales overlap with industrial activity, there is considerable interest from managers to be able to reliably forecast when and where blue whales are most likely to be in the area. In a series of five chapters and utilizing multiple different data sources (dedicated boat surveys, oceanographic data, acoustic recordings, remotely sensed environmental data, opportunistic blue whale sightings information), I will attempt to describe, quantify, and predict where blue whales are found in relation to their environment. Each chapter will evaluate the distribution of blue whales relative to the environment at different scales in space (ranging from 4 km to 25 km resolution) and time (ranging from daily to seasonal resolution). One overarching method I am using throughout my PhD is species distribution modeling. Having just completed my research review with my doctoral committee last week, I’ll share this aspect of my research proposal that I’ve particularly enjoyed reading, writing, and thinking about.
A pair of blue whales surfacing in the South Taranaki Bight region of New Zealand. Drone piloted by Todd Chandler during the 2017 field season.
Species distribution models (SDMs), which are sometimes referred to as habitat models or ecological niche models, are mathematical algorithms that combine observations of a species with environmental conditions at their observed locations, to gain ecological insight and predict spatial distributions of the species (Elith and Leathwick, 2009; Redfern et al., 2006). Any model is just one description of what is occurring in the natural world. Just as there are many ways to describe something with words and many languages to do so, there are many options for modeling frameworks and approaches, with stark and nuanced differences. My labmate and friend Solene Derville has equated the number of choices one has for SDMs to the cracker section in an American grocery store. When navigating all of these choices and considerations, it is important to remember that no model will ever be completely correct—it is our best attempt at describing a complex natural system—and as an analyst we need to do the best that we can with the data available to address the ecological questions at hand. As it turns out, the dividing line between quantitative analysis and philosophy is thin at times. What may seem at first like a purely objective, statistical endeavor requires careful consideration and fundamental decision-making on the part of the analyst.
Ecosystems are multifaceted, complex, and hierarchical. They are comprised of multiple physical and biological components, which operate at multiple scales across space and time. As Dr. Simon Levin stated in at 1989 MacArthur Award lecture on the topic of scale in ecology:
“A good model does not attempt to reproduce every detail of the biological system; the system itself suffices for that purpose as the most detailed model of itself. Rather, the objective of a model should be to ask how much detail can be ignored without producing results that contradict specific sets of observations, on particular scales of interest” (Levin, 1992).
The question of scale is central to ecology. As many biology students learn in their first introductory classes, parsimony is “The principle that the most acceptable explanation of an occurrence, phenomenon, or event is the simplest, involving the fewest entities, assumptions, or changes” (Oxford Dictionary). In other words, the best explanation is the simplest one. One challenge in ecological modeling, including SDMs, is to select spatial and temporal scales as coarse as possible for the most parsimonious—the most straightforward—model, while still being fine enough to capture relevant patterns. Another critical consideration is the scale of the question you are interested in answering. The scale of the analysis must match the scale at which you want to make inferences about the ecology of a species.
Similarly, the issue of complexity is central to distribution modeling. Overly simple models may not be able to adequately describe the relationship between species occurrence and the environment. In contrast, highly complex models may have very high explanatory power, but risk ascribing an ecological pattern to noise in the data (Merow et al., 2014), in other words, finding patterns that aren’t real. Furthermore, highly complex models tend to have poorer predictive capacity than simpler models (Merow et al., 2014). There is a trade-off between descriptive and predictive power in SDMs (Derville et al., 2018). Therefore, a key component in the SDM process is establishing the end goal of the model with respect to the region of interest, scale, explanatory power, predictive capacity, and in many cases management need.
Finally, any model is ultimately limited by the data available and the scale at which it was collected (Elith and Leathwick, 2009; Guillera-Arroita et al., 2015; Redfern et al., 2006). Prior knowledge of what environmental features are important to the species of interest is often limited at the time of the data collection effort, and data collection is constrained by when it is logistically feasible to sample. For example, we collect detailed oceanographic data during the summer months when it is practical to get out on the water, satellite imagery of sea surface temperature might be unavailable during times of cloud cover, and people are more likely to report blue whale sightings in areas where there is more human activity. Therefore, useful SDMs that address both ecological and management needs typically balance the scale of analysis and model complexity with the limitations of the data.
Managers and politicians within the New Zealand government are interested in a tool to predict when and where blue whales are most likely to be, based on sound ecological analysis. This is one of the end-goals of my PhD, but in the meantime, I am grappling with the appropriate scales of analysis, and attempting to balance questions of model complexity, explanatory power, and predictive capacity. There is no single, correct answer, and so my process is in part quantitative analysis, part philosophy, and all with the goal of increased ecological understanding and conservation of a species.
A blue whale breaks the surface. As I grapple with questions of model complexity and scale of analysis, I sometimes need a reminder that behind each data point is a blue whale, and what a privilege it is to study them. Photo by Leigh Torres.
References:
Derville, S., Torres, L. G., Iovan, C., and Garrigue, C. (2018). Finding the right fit: Comparative cetacean distribution models using multiple data sources and statistical approaches. Divers. Distrib. 24, 1657–1673. doi:10.1111/ddi.12782.
Elith, J., and Leathwick, J. R. (2009). Species Distribution Models: Ecological Explanation and Prediction Across Space and Time. Annu. Rev. Ecol. Evol. Syst. 40, 677–697. doi:10.1146/annurev.ecolsys.110308.120159.
Guillera-Arroita, G., Lahoz-Monfort, J. J., Elith, J., Gordon, A., Kujala, H., Lentini, P. E., et al. (2015). Is my species distribution model fit for purpose? Matching data and models to applications. Glob. Ecol. Biogeogr. 24, 276–292. doi:10.1111/geb.12268.
Levin, S. A. (1992). The problem of pattern and scale. Ecology 73, 1943–1967.
Merow, C., Smith, M. J., Edwards, T. C., Guisan, A., Mcmahon, S. M., Normand, S., et al. (2014). What do we gain from simplicity versus complexity in species distribution models? Ecography (Cop.). 37, 1267–1281. doi:10.1111/ecog.00845.
Redfern, J. V., Ferguson, M. C., Becker, E. A., Hyrenbach, K. D., Good, C., Barlow, J., et al. (2006). Techniques for cetacean-habitat modeling. Mar. Ecol. Prog. Ser. 310, 271–295. doi:10.3354/meps310271.
By: Alexa Kownacki, Ph.D. Student, OSU Department of Fisheries and Wildlife, Geospatial Ecology of Marine Megafauna Lab
Data analysis is often about parsing down data into manageable subsets. My project, which spans 34 years and six study sites along the California coast, requires significant data wrangling before full analysis. As part of a data analysis trial, I first refined my dataset to only the San Diego survey location. I chose this dataset for its standardization and large sample size; the bulk of my sightings, over 4,000 of the 6,136, are from the San Diego survey site where the transect methods were highly standardized. In the next step, I selected explanatory variable datasets that covered the sighting data at similar spatial and temporal resolutions. This small endeavor in analyzing my data was the first big leap into understanding what questions are feasible in terms of variable selection and analysis methods. I developed four major hypotheses for this San Diego site.
The study species: common bottlenose dolphin (Tursiops truncatus) seen along the California coastline in 2015. Image source: Alexa Kownacki.
Hypotheses:
H1: I predict that bottlenose dolphin sightings along the San Diego transect throughout the years 1981-2015 exhibit clustered distribution patterns as a result of the patchy distributions of both the species’ preferred habitats, as well as the social nature of bottlenose dolphins.
H2: I predict there would be higher densities of bottlenose dolphin at higher latitudes spanning 1981-2015 due to prey distributions shifting northward and less human activities in the northerly sections of the transect.
H3: I predict that during warm (positive) El Niño Southern Oscillation (ENSO) months, the dolphin sightings in San Diego would be distributed more northerly, predominantly with prey aggregations historically shifting northward into cooler waters, due to (secondarily) increasing sea surface temperatures.
H4: I predict that along the San Diego coastline, bottlenose dolphin sightings are clustered within two kilometers of the six major lagoons, with no specific preference for any lagoon, because the murky, nutrient-rich waters in the estuarine environments are ideal for prey protection and known for their higher densities of schooling fishes.
Data Description:
The common bottlenose dolphin (Tursiops truncatus) sighting data spans 1981-2015 with a few gap years. Sightings cover all months, but not in all years sampled. The same transect in San Diego was surveyed in a small, rigid-hulled inflatable boat with approximately a two-kilometer observation area (one kilometer surveyed 90 degrees to starboard and port of the bow).
I wanted to see if there were changes in dolphin distribution by latitude and, if so, whether those changes had a relationship to ENSO cycles and/or distances to lagoons. For ENSO data, I used the NOAA database that provides positive, neutral, and negative indices (1, 0, and -1, respectively) by each month of each year. I matched these ENSO data to my month-date information of dolphin sighting data. Distance from each lagoon was calculated for each sighting.
Figure 1. Map representing the San Diego transect, represented with a light blue line inside of a one-kilometer buffered “sighting zone” in pale yellow. The dark pink shapes are dolphin sightings from 1981-2015, although some are stacked on each other and cannot be differentiated. The lagoons, ranging in size, are color-coded. The transect line runs from the breakwaters of Mission Bay, CA to Oceanside Harbor, CA.
Results:
H1:True, dolphins are clustered and do not have a uniform distribution across this area. Spatial analysis indicated a less than a 1% likelihood that this clustered pattern could be the result of random chance (Fig. 1, z-score = -127.16, p-value < 0.0001). It is well-known that schooling fishes have a patchy distribution, which could influence the clustered distribution of their dolphin predators. In addition, bottlenose dolphins are highly social and although pods change in composition of individuals, the dolphins do usually transit, feed, and socialize in small groups.
Figure 2. Summary from the Average Nearest Neighbor calculation in ArcMap 10.6 displaying that bottlenose dolphin sightings in San Diego are highly clustered. When the z-score, which corresponds to different colors on the graphic above, is strongly negative (< -2.58), in this case dark blue, it indicates clustering. Because the p-value is very small, in this case, much less than 0.01, these results of clustering are strongly significant.
H2:False, dolphins do not occur at higher densities in the higher latitudes of the San Diego study site. The sightings are more clumped towards the lower latitudes overall (p < 2e-16), possibly due to habitat preference. The sightings are closer to beaches with higher human densities and human-related activities near Mission Bay, CA. It should be noted, that just north of the San Diego transect is the Camp Pendleton Marine Base, which conducts frequent military exercises and could deter animals.
Figure 3. Histogram comparing the latitudes with the frequency of dolphin sightings in San Diego, CA. The x-axis represents the latitudinal difference from the most northern part of the transect to each dolphin sighting. Therefore, a small difference would translate to a sighting being in the northern transect areas whereas large differences would translate to sightings being more southerly. This could be read from left to right as most northern to most southern. The y-axis represents the frequency of which those differences are seen, that is, the number of sightings with that amount of latitudinal difference, or essentially location on the transect line. Therefore, you can see there is a peak in the number of sightings towards the southern part of the transect line.
H3: False, during warm (positive) El Niño Southern Oscillation (ENSO) months, the dolphin sightings in San Diego were more southerly. In colder (negative) ENSO months, the dolphins were more northerly. The differences between sighting latitude and ENSO index was significant (p<0.005). Post-hoc analysis indicates that the north-south distribution of dolphin sightings was different during each ENSO state.
Figure 4. Boxplot visualizing distributions of dolphin sightings latitudinal differences and ENSO index, with -1,0, and 1 representing cold, neutral, and warm years, respectively.
H4:True, dolphins are clustered around particular lagoons. Figure 5 illustrates how dolphin sightings nearest to Lagoon 6 (the San Dieguito Lagoon) are always within 0.03 decimal degrees. Because of how these data are formatted, decimal degrees is the easiest way to measure change in distance (in this case, the difference in latitude). In comparison, dolphins at Lagoon 5 (Los Penasquitos Lagoon) are distributed across distances, with the most sightings further from the lagoon.
Figure 5. Bar plot displaying the different distances from dolphin sighting location to the nearest lagoon in San Diego in decimal degrees. Note: Lagoon 4 is south of the study site and therefore was never the nearest lagoon.
I found a significant difference between distance to nearest lagoon in different ENSO index categories (p < 2.55e-9): there is a significant difference in distance to nearest lagoon between neutral and negative values and positive and neutral years. Therefore, I hypothesize that in neutral ENSO months compared to positive and negative ENSO months, prey distributions are changing. This is one possible hypothesis for the significant difference in lagoon preference based on the monthly ENSO index. Using a violin plot (Fig. 6), it appears that Lagoon 5, Los Penasquitos Lagoon, has the widest variation of sighting distances in all ENSO index conditions. In neutral years, Lagoon 0, the Buena Vista Lagoon has multiple sightings, when in positive and negative years it had either no sightings or a single sighting. The Buena Vista Lagoon is the most northerly lagoon, which may indicate that in neutral ENSO months, dolphin pods are more northerly in their distribution.
Figure 6. Violin plot illustrating the distance from lagoons of dolphin sightings under different ENSO conditions. There are three major groups based on ENSO index: “-1” representing cold years, “0” representing neutral years, and “1” representing warm years. On the x-axis are lagoon IDs and on the y-axis is the distance to the nearest lagoon in decimal degrees. The wider the shapes, the more sightings, therefore Lagoon 6 has many sightings within a very small distance compared to Lagoon 5 where sightings are widely dispersed at greater distances.
Bottlenose dolphins foraging in a small group along the California coast in 2015. Image source: Alexa Kownacki.
Takeaways to science and management:
Bottlenose dolphins have a clustered distribution which seems to be related to ENSO monthly indices, and likely, their social structures. From these data, neutral ENSO months appear to have something different happening compared to positive and negative months, that is impacting the sighting distributions of bottlenose dolphins off the San Diego coastline. More research needs to be conducted to determine what is different about neutral months and how this may impact this dolphin population. On a finer scale, the six lagoons in San Diego appear to have a spatial relationship with dolphin sightings. These lagoons may provide critical habitat for bottlenose dolphins and/or for their preferred prey either by protecting the animals or by providing nutrients. Different lagoons may have different spans of impact, that is, some lagoons may have wider outflows that create larger nutrient plumes.
Other than the Marine Mammal Protection Act and small protected zones, there are no safeguards in place for these dolphins, whose population hovers around 500 individuals. Therefore, specific coastal areas surrounding lagoons that are more vulnerable to habitat loss, habitat degradation, and/or are more frequented by dolphins, may want greater protection added at a local, state, or federal level. For example, the Batiquitos and San Dieguito Lagoons already contain some Marine Conservation Areas with No-Take Zones within their reach. The city of San Diego and the state of California need better ways to assess the coastlines in their jurisdictions and how protecting the marine, estuarine, and terrestrial environments near and encompassing the coastlines impacts the greater ecosystem.
This dive into my data was an excellent lesson in spatial scaling with regards to parsing down my data to a single study site and in matching my existing data sets to other data that could help answer my hypotheses. Originally, I underestimated the robustness of my data. At first, I hesitated when considering reducing the dolphin sighting data to only include San Diego because I was concerned that I would not be able to do the statistical analyses. However, these concerns were unfounded. My results are strongly significant and provide great insight into my questions about my data. Now, I can further apply these preliminary results and explore both finer and broader scale resolutions, such as using the more precise ENSO index values and finding ways to compare offshore bottlenose dolphin sighting distributions.
By Rachael Orben, Assistant Professor (Senior Research), Seabird Oceanography Lab
This February I had the opportunity to spend two weeks at Midway Atoll National Wildlife Refuge in the Papahānaumokuākea Marine National Monument. I was there to GPS track black-footed and Laysan albatross during their short chick-brooding foraging trips. Two weeks is just enough time since the albatross are taking short trips (3-5 days) to feed their rapidly growing chicks.
My first visit to Midway (2016 blog post) occurred right as the black-footed albatross chicks were hatching (quickly followed by the Laysan albatross chicks). This time, we arrived almost exactly when I had left off. The oldest chicks were just about two weeks old. This shift in phenology meant that, though subtle, each day offered new insights for me as I watched chicks transform into large aware and semi-mobile birds. By the time we left, unattended chicks were rapidly multiplying as the adults shifted to the chick-rearing stage. During chick rearing, both parents leave the chick unattended and take longer foraging trips.
Our research goal was to collect tracking data from both species that can be used to address a couple of research questions. First of all, winds can aid, or hinder albatross foraging and flight efficiency (particularly during the short brooding trips). In the North Pacific, the strength and direction of the winds are influenced by the ENSO (El Niño Southern Oscillation) cycles. The day after we left Midway, NOAA issued an El Niño advisory indicating weak El Nino conditions. We know from previous work at Tern Island (farther east and farther south at 23.87 N, -166.28 W) that El Niño improves foraging for Laysan albatrosses during chick brooding, while during La Niña reproductive success is lower (Thorne et al., 2016). However, since Midway is farther north, and farther west the scenario might be different there. Multiple years of GPS tracking data are needed to address this question and we hope to return to collect more data next year (especially if La Niña follows the El Niño as is often the case).
We will also overlap the tracking data with fishing boat locations from the Global Fishing Watch database to assess the potential for birds from Midway to interact with high seas fisheries during this time of year (project description, associated blog post). Finally, many of the tags we deployed incorporated a barometric pressure sensor and the data can be used to estimate flight heights relative to environmental conditions such as wind strength. This type of data is key to assessing the impact of offshore wind energy (Kelsey et al., 2018).
How to track an albatross
To track an albatross we use small GPS tags that we tape to the back feathers. After the bird returns from a foraging trip, we remove the tape from the feathers and take the datalogger off. Then we recharge the battery and download the data!
Catching a tagged albatross that just returned from its foraging trip. Photo V. Ternisien
Recording data. Photo V. Ternisien
Getting ready to remove the GPS datalogger. Photo V. Ternisien
The GPS datalogger was taped to the back feathers of the bird. To remove the tag we simply peel off the tape. Photo V. Ternisien
My previous visit to Midway occurred just after house mice were discovered attacking incubating adult albatrosses. Since then, a lot of thought and effort had gone into developing a plan to eradicate mice from Midway. You can find out more via Island Conservation’s Midway blogs and the USFWS.
Dancing.
There is always something happening in a seabird colony.
Red-tailed tropicbird
From left to right: A Laysan albatross, a hybrid, and a black-footed albatross.
The colony was never quiet. Even at night.
A GPS tagged Laysan albatross.
Habitat restoration.
A short-tailed albatross sitting next to his chick (the large black chick the his right).
References
Kelsey, E. C., Felis, J. J., Czapanskiy, M., Pereksta, D. M., & Adams, J. (2018). Collision and displacement vulnerability to offshore wind energy infrastructure among marine birds of the Pacific Outer Continental Shelf. Journal of Environmental Management, 227, 229–247. http://doi.org/10.1016/j.jenvman.2018.08.051
Thorne, L. H., Conners, M. G., Hazen, E. L., Bograd, S. J., Antolos, M., Costa, D. P., & Shaffer, S. A. (2016). Effects of El Niño-driven changes in wind patterns on North Pacific albatrosses. Journal of the Royal Society Interface, 13(119), 20160196. http://doi.org/10.1098/rsif.2016.0196
I have the privilege of studying the largest animals on the planet: blue whales (Balaenoptera musculus). However, in order to understand the ecology, distribution, and habitat use patterns of these ocean giants, I have dedicated the past several months to studying something much smaller: krill (Nyctiphanes australis). New Zealand’s South Taranaki Bight region (“STB”, Figure 1) is an important foraging ground for a unique population of blue whales [1,2]. A wind-driven upwelling system off of Kahurangi Point (the “X” in Figure 1) generates productivity in the region [3], leading to an abundance of krill [4], the desired blue whale prey [5].
Figure 1. A map of New Zealand, with the South Taranaki Bight (STB) region delineated by the black box. Cape Farewell is denoted by a star, and Kahurangi point is denoted by an X.
Figure 4. A blue whale lunges on an aggregation of krill. UAS piloted by Todd Chandler.
Krill (Nyctiphanes australis), blue whale prey in New Zealand’s South Taranaki Bight region. Image: Te Papa.
Our blue whale research team collected a multitude of datastreams in three different years, including hydroacoustic data to map krill distribution throughout our study region. The summers of 2014 and 2017 were characterized by what could be considered “typical” conditions: A plume of cold, upwelled water curving its way around Cape Farewell (marked with the star in Figure 1) and entering the South Taranaki Bight, spurring a cascade of productivity in the region. The 2016 season, however, was different. The surface water temperatures were hot, and the whales were not where we expected to find them.
Figure 2. Sea surface temperature maps of the South Taranaki Bight region in each of our three study years. The white circles indicate where most blue whale sightings were made in each year. Note the very warm temperatures in 2016, and more westerly location of blue whale sightings.
What happened to the blue whales’ food source under these different conditions in 2016? Before I share some preliminary findings from my recent analyses, it is important to note that there are many possible ways to measure krill availability. For example, the number of krill aggregations, as well as how deep, thick, and dense those aggregations are in an area will all factor into how “desirable” krill patches are to a blue whale. While there may not be “more” or “less” krill from one year to the next, it may be more or less accessible to a blue whale due to energetic costs of capturing it. Here is a taste of what I’ve found so far:
In 2016, when surface waters were warm, the krill aggregations were significantly deeper than in the “typical” years (ANOVA, F=7.94, p <0.001):
Figute 3. Boxplots comparing the median krill aggregation depth in each of our three survey years.
The number of aggregations was not significantly different between years, but as you can see in the plot below (Figure 4) the krill were distributed differently in space:
Figure 4. Map of the South Taranaki Bight region with the number of aggregations per 4 km^2, standardized by vessel survey effort. The darker colors represent areas with a higher density of krill aggregations.
While the bulk of the krill aggregations were located north of Cape Farewell under typical conditions (2014 and 2017), in the warm year (2016) the krill were not in this area. Rather, the area with the most aggregations was offshore, in the western portion of our study region. Now, take a look at the same figure, overlaid with our blue whale sighting locations:
Figure 5. Map of standardized number of krill aggregations, overlaid with blue whale sighting locations in red stars.
Where did we find the whales? In each year, most whale encounters were in the locations where the most krill aggregations were found! Not only that, but in 2016 the whales responded to the difference in krill distribution by shifting their distribution patterns so that they were virtually absent north of Cape Farewell, where most sightings were made in the typical years.
The above figures demonstrate the importance of studying an ecosystem. We could puzzle and speculate over why the blue whales were further west in the warm year, but the story that is emerging in the krill data may be a key link in our understanding of how the ecosystem responds to warm conditions. While the focus of my dissertation research is blue whales, they do not live in isolation. It is through understanding the ecosystem-scale story that we can better understand blue whale ecology in the STB. As I continue modeling the relationships between oceanography, krill, and blue whales in warm and typical years, we are beginning to scratch the surface of how blue whales may be responding to their environment.
Torres LG. 2013 Evidence for an unrecognised blue whale foraging ground in New Zealand. New Zeal. J. Mar. Freshw. Res.47, 235–248. (doi:10.1080/00288330.2013.773919)
Barlow DR et al. 2018 Documentation of a New Zealand blue whale population based on multiple lines of evidence. Endanger. Species Res.36, 27–40. (doi:https://doi.org/10.3354/esr00891)
Shirtcliffe TGL, Moore MI, Cole AG, Viner AB, Baldwin R, Chapman B. 1990 Dynamics of the Cape Farewell upwelling plume, New Zealand. New Zeal. J. Mar. Freshw. Res.24, 555–568. (doi:10.1080/00288330.1990.9516446)
Bradford-Grieve JM, Murdoch RC, Chapman BE. 1993 Composition of macrozooplankton assemblages associated with the formation and decay of pulses within an upwelling plume in greater cook strait, New Zealand. New Zeal. J. Mar. Freshw. Res.27, 1–22. (doi:10.1080/00288330.1993.9516541)
Gill P. 2002 A blue whale (Balaenoptera musculus) feeding ground in a southern Australian coastal upwelling zone. J. Cetacean Res. Manag.4, 179–184.