By Amanda Rose Kent, College of Earth Ocean and Atmospheric Sciences, OSU, GEMM Lab/Krill Seeker undergraduate intern
If you asked me five years ago where I’d thought I’d be today, the answer I would give would not reflect where I am now. Back then, I was a customer service representative for a hazardous waste company, and I believed that going to university and participating in research was a straightforward experience. I learned soon after I left that career and began my journey at OSU in 2020 that I wasn’t even remotely aware of the process. I knew that as part of my oceanography degree I would need to become involved in some form of research, but I had no idea where to start.
I started looking through the Oregon State website and I eventually found an outdated flier from 2018 that advertised a lab that studied plankton in Antarctica, and that was when I first reached out to Dr. Kim Bernard. My journey took off from there. As an undergraduate researcher in the URSA Engage program working with Kim and one of her graduate students, Rachel, I conducted a literature review on the ecosystem services provided by two species of krill off the coast of Oregon, including their value to baleen whales. After learning all I could from the literature about krill and how important they were to the ocean, I knew that there was so much more to learn and that this was the topic I wanted to continue to pursue. After I completed the URSA program, I remained a member of Kim’s zooplankton ecology lab.
While continuing to work with Rachel, I was given the opportunity to join the GEMM Lab’s Project HALO for a daylong cruise conducting a whale survey along the Newport Hydrographic Line. I was initially brought on to learn how to use the echosounder to collect krill data but unfortunately, the device had technical difficulties and Rachel and I were no longer needed. We decided to go on the cruise anyway, and I was able to instead learn how to survey for marine mammals (it’s not as easy as it may seem, but still very fun!).
Figure 1. Enjoying the point of view from the crow’s nest on the R/V Pacific Storm, but also very cold.
Soon, another opportunity arose to apply for a brand-new program called ARC-Learn. This two-year research program focuses on studying the Arctic using publicly available data, and with the support of my mentors, I applied and was accepted. Initially I found that there were no mentors within the program that studied krill, so I found myself becoming immersed in a new topic: harmful algal blooms (HABs). Determined to incorporate krill into this research, I started looking through the literature trying to develop my hypothesis that HABs affected zooplankton in some way. There was evidence to potentially support my hypothesis, but I ended up encountering numerous data gaps in the region I was studying. After months of roadblocks, I eventually started feeling defeated and regretted applying for the program. Rachel was quick to remind me that all experiences are valuable experiences, and that I was still gaining new skills I could use in graduate school or my career.
As my undergraduate degree progressed, I continued supporting Rachel in her graduate research, spending some time during the summer processing krill samples by sorting, sexing, and drying them to crush them into pellets. Our goal was to process them in an instrument called a bomb calorimeter, which is used to quantify the caloric content of prey species and help us better understand the energy flux required for animals higher up the food chain (like whales) and the amount they need to eat. I was only able to do this for a few weeks before heading out on the experience of a lifetime, spending three weeks on a ship traveling around the Bering, Chukchi, and Beaufort Seas with one of my ARC-Learn mentors. It was a great opportunity for me to see the toxic phytoplankton (which can form HABs) I had been studying and learn about methods of sample collection and processing. If I could go back and do it again, I’d go in a heartbeat.
Figure 2. Pulling out all of the animal biomass out of the Arctic sediment.
At the beginning of my bachelor’s degree, I had expected to just work with Kim and conduct research within her lab. Instead, I have had opportunities I would never have expected five years ago. I have learned a vast amount from my graduate mentor, Rachel, which has helped influence my trajectory in my degree. I have had the privilege to not only meet giants in the field I’m interested in, but also work with them and learn from them, and to spend three weeks in the Arctic Ocean. The experiences I have had throughout this roller-coaster helped me develop a project idea with new mentors that I eventually hope to pursue in my master’s degree. I wasn’t prepared for the number of adjustments I would make to find new experiences and start new projects, but all the experiences I had were necessary to learn about what I was interested in and what I wanted to pursue. Looking back on it all today, I have zero regrets.
Figure 3. A picture of the Norseman II, the ship I was on in the Arctic, taken by the Japanese ship JAMSTEC on a short rendezvous between the Chukchi and Beaufort Seas.
I wrote my last blogpost in the midst of winter and feeling overwhelmed as I was trying to fly to the US at the peak of the omicron pandemic… Since then, morale has improved exponentially. I have spent two months in the company of my delightful GEMM lab friends, nerding over statistics, sharing scientific conversations, drinking (good!) beer and enjoying the company of this great group of people. During that stay, I was able to focus on my OPAL project more than I have ever been able to, as I set myself the goal of not getting distracted by anything else during my stay in Newport.
The only one distraction that I do not regret is a post I read one morning on the Cetal Fauna Facebook page, a group of cetacean experts and lovers who share news, opinions, photos… anything cetacean related! Someone was posting a photo of a humpback whale stranded in the 1990s’ on Coolum beach, on the east coast of Australia, which is known as a major humpback whale migratory corridor. The story said that (probably with considerable effort) the whale was refloated by many different individuals and organizations present at the beach on that day, specifically Sea World Research, Rescue & Conservation.
I felt very touched by this story and the photo that illustrated it (Figure 1). Seeing all these people come together in this risky operation to save this sea giant is quite something. And the fact that they succeeded was even more impressive! Indeed, baleen whales strand less commonly than toothed whales but their chances of survival when they do so are minimal. In addition to the actual potential damages that might have caused the whale to strand in the first place (entanglements, collisions, diseases etc.), the beaching itself is likely to hurt the animal in a permanent way as their body collapses under their own weight usually causing a cardiovascular failure (e.g., Fernández et al., 2005). The rescue of baleen whales is also simply impaired by the sheer size and weight of these animals. Compared to smaller toothed whales such as pilot whales and false killer whales that happen to strand quite frequently over some coastlines, baleen whales are almost impossible to move off the beach and getting close to them when beached can be very dangerous for responders. For these reasons, I found very few reports and publications mentioning successful rescues of beached baleen whales (e.g., Priddel and Wheeler, 1997; Neves et al., 2020).
Figure 1: Stranded humpback whale on Coolum Beach, East Australia, in 1996. Look at the size of the fluke compared to the men who are trying to rescue her! Luckily, that risky operation ended well. This image won Australian Time Magazine Cover of the year. Credit: Sea World Research, Rescue and Conservation. Photo posted by P. Garbett on https://www.facebook.com/groups/CetalFauna – February 26, 2022)
Now the story gets even better… the following day I received an email from Ted Cheeseman, director and co-founder of Happywhale, a collaborative citizen science tool to share and match photographes of cetaceans (initially only humpback whales but has extended to other species) to recognize individuals based on the unique patterns of the their fluke or dorsal fin. The fluke of the whale stranded in Australia in 1991 had one and only match within the Happywhale immense dataset… and that match was to a whale seen in New Caledonia (Figure 2). “HNC338” was the one!
Figure 2: Happy whale page showing the match of HNC338 between East Australia and New Caledonia. https://happywhale.com/individual/78069;enc=284364?fbclid=IwAR1QEG_6JkpH_k2UrF-qp-9qrOboHYakKjlTj0lLbDFygjN5JugkkKVeMQw
Since I conducted my PhD on humpback whale spatial ecology in New Caledonia, I have continued working on a number of topics along with my former PhD supervisor, Dr Claire Garrigue, in New Caledonia. Although I do not remember each and every whale from her catalogue (composed of more than 1600 humpback whales as of today), I do love a good “whale tale” and I was eager to know who this HNC338 was. I quickly looked into Claire’s humpback whale database and sure enough I found it there: encountered at the end of the 2006 breeding season on September 12th, at a position of 22°26.283’S and 167°01.991’E and followed for an hour. Field notes reported a shy animal that kept the boat at a distance. But most of all, HNC338 was genetically identified as a female and was accompanied by a calf during that season! The calf was particularly big, as expected at this time of the season. What an inspiring thing to think that this whale, stranded in 1996, was resighted 10 years later in a neighboring breeding ground, apparently healthy and raising a calf of her own.
As genetic paternity analysis have been conducted on many New Caledonia calf biopsy samples as part of the Sexy Singing project conducted with our colleagues from St Andrews University in Scotland, we might be able to identify the calf’s father in this breeding stock. Thanks to the great amount of data shared and collected through Happywhale, we are discovering more and more about whale migratory patterns and behavior. It might as well be that this calf’s father was one of those whales that seem to roam over several different breeding grounds (New Caledonia and East Australia). This story is far from finished…
Figure 3: A (pretty bad!) photo of HNC338’s fluke. Luckily the Happywhale matching algorithm is very efficient and was able to detect the similarities of the fluke’s trailing edge compared to figure 1 (Cheeseman et al., 2021). Also of note, see that small dorsal fin popping out of the waters behind big mama’s fluke? That’s her calf!
From the people who pulled this whale back into the water in 1996, to the scientists and cetacean enthusiasts who shared their data and whale photos online, this story once again shows us that hope lies in cooperation! Happywhale was only created in 2015 but since then it has brought together the general public and the scientists to contribute over 465,000 photos allowing the identification of 75,000 different individuals around the globe. In New Caledonia, in Oregon and elsewhere, I hope that these collective initiatives grow more and more in the future, to the benefit of biodiversity and people.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get weekly updates and more! Just add your name into the subscribe box on the left panel.
References
Cheeseman, T., Southerland, K., Park, J., Olio, M., Flynn, K., Calambokidis, J., et al. (2021). Advanced image recognition: a fully automated, high-accuracy photo-identification matching system for humpback whales. Mamm. Biol. doi:10.1007/s42991-021-00180-9.
Fernández, A., Edwards, J. F., Rodríguez, F., Espinosa De Los Monteros, A., Herráez, P., Castro, P., et al. (2005). “Gas and fat embolic syndrome” involving a mass stranding of beaked whales (Family Ziphiidae) exposed to anthropogenic sonar signals. Vet. Pathol. 42, 446–457. doi:10.1354/vp.42-4-446.
Neves, M. C., Neto, H. G., Cypriano-Souza, A. L., da Silva, B. M. G., de Souza, S. P., Marcondes, M. C. C., et al. (2020). Humpback whale (megaptera novaeangliae) resighted eight years after stranding. Aquat. Mamm. 46, 483–487. doi:10.1578/AM.46.5.2020.483.
Priddel, D., and Wheeler, R. (1997). Rescue of a Bryde’s whale Balaenoptera edeni entrapped in the Manning River, New South Wales: Unmitigated success or unwarranted intervention? Aust. Zool. 30, 261–271. doi:10.7882/AZ.1997.002.
2MS Student, OSU Department of Fisheries, Wildlife, and Conservation Sciences, Seabird Oceanography Lab
The marine environment is dynamic, and mobile animals must respond to the patchy and ephemeral availability of resource in order to make a living (Hyrenbach et al. 2000). Climate change is making ocean ecosystems increasingly unstable, yet these novel conditions can be difficult to document given the vast depth and remoteness of most ocean locations. Marine megafauna species such as marine mammals and seabirds integrate ecological processes that are often difficult to observe directly, by shifting patterns in their distribution, behavior, physiology, and life history in response to changes in their environment (Croll et al. 1998, Hazen et al. 2019). These mobile marine animals now face additional challenges as rising temperatures due to global climate change impact marine ecosystems worldwide (Hazen et al. 2013, Sydeman et al. 2015, Silber et al. 2017, Becker et al. 2019). Given their mobility, visibility, and integration of ocean processes across spatial and temporal scales, these marine predator species have earned the reputation as effective ecosystem sentinels. As sentinels, they have the capacity to shed light on ecosystem function, identify risks to human health, and even predict future changes (Hazen et al. 2019). So, let’s explore a few examples of how studying marine megafauna has revealed important new insights, pointing toward the importance of monitoring these sentinels in a rapidly changing ocean.
Cairns (1988) is often credited as first promoting seabirds as ecosystem sentinels and noted several key reasons why they were perfect for this role: (1) Seabirds are abundant, wide-ranging, and conspicuous, (2) although they feed at sea, they must return to land to nest, allowing easier observation and quantification of demographic responses, often at a fraction of the cost of traditional, ship-based oceanographic surveys, and therefore (3) parameters such as seabird reproductive success or activity budgets may respond to changing environmental conditions and provide researchers with metrics by which to assess the current state of that ecosystem.
The unprecedented 2014-2016 North Pacific marine heatwave (“the Blob”) caused extreme ecosystem disruption over an immense swath of the ocean (Cavole et al. 2016). Seabirds offered an effective and morbid indication of the scale of this disruption: Common murres (Uria aalge), an abundant and widespread fish-eating seabird, experienced widespread breeding failure across the North Pacific. Poor reproductive performance suggested that there may have been fewer small forage fish around and that these changes occurred at a large geographic scale. The Blob reached such an extreme as to kill immense numbers of adult birds, which professional and community scientists found washed up on beach-surveys; researchers estimate that an incredible 1,200,000 murres may have died from starvation during this period (Piatt et al. 2020). While the average person along the Northeast Pacific Coast during this time likely didn’t notice any dramatic difference in the ocean, seabirds were shouting at us that something was terribly wrong.
Happily, living seabirds also act as superb ecosystem sentinels. Long-term research in the Gulf of Maine by U.S. and Canadian scientists monitors the prey species provisioned by adult seabirds to their chicks. Will has spent countless hours over five summers helping to conduct this research by watching terns (Sterna spp.) and Atlantic puffins (Fratercula arctica) bring food to their young on small islands off the Maine coast. After doing this work for multiple years, it’s easy to notice that what adults feed their chicks varies from year to year. It was soon realized that these data could offer insight into oceanographic conditions and could even help managers assess the size of regional fish stocks. One of the dominant prey species in this region is Atlantic herring (Clupea harengus), which also happens to be the focus of an economically important fishery. While the fishery targets four or five-year-old adult herring, the seabirds target smaller, younger herring. By looking at the relative amounts and sizes of young herring collected by these seabirds in the Gulf of Maine, these data can help predict herring recruitment and the relative number of adult herring that may be available to fishers several years in the future (Scopel et al. 2018). With some continued modelling, the work that we do on a seabird colony in Maine with just a pair of binoculars can support or maybe even replace at least some of the expensive ship-based trawl surveys that are now a popular means of assessing fish stocks.
A common tern (Sterna hirundo) with a young Atlantic herring from the Gulf of Maine, ready to feed its chick (Photo courtesy of the National Audubon Society’s Seabird Institute)
For more far-ranging and inaccessible marine predators such as whales, measuring things such as dietary shifts can be more challenging than it is for seabirds. Nevertheless, whales are valuable ecosystem sentinels as well. Changes in the distribution and migration phenology of specialist foragers such as blue whales (Balaenoptera musculus) and North Atlantic right whales (Eubalaena glacialis) can indicate relative changes in the distribution and abundance of their zooplankton prey and underlying ocean conditions (Hazen et al. 2019). In the case of the critically endangered North Atlantic right whale, their recent declines in reproductive success reflect a broader regime shift in climate and ocean conditions. Reduced copepod prey has resulted in fewer foraging opportunities and changing foraging grounds, which may be insufficient for whales to obtain necessary energetic stores to support calving (Gavrilchuk et al. 2021, Meyer-Gutbrod et al. 2021). These whales assimilate and showcase the broad-scale impacts of climate change on the ecosystem they inhabit.
Blue whales that feed in the rich upwelling system off the coast of California rely on the availability of their krill prey to support the population (Croll et al. 2005). A recent study used acoustic monitoring of blue whale song to examine the timing of annual population-level transition from foraging to breeding migration compared to oceanographic variation, and found that flexibility in timing may be a key adaptation to persistence of this endangered population facing pressures of rapid environmental change (Oestreich et al. 2022). Specifically, blue whales delayed the transition from foraging to breeding migration in years of the highest and most persistent biological productivity from upwelling, and therefore listening to the vocalizations of these whales may be valuable indicator of the state of productivity in the ecosystem.
Figure reproduced from Oestreich et al. 2022, showing relationships between blue whale life-history transition and oceanographic phenology of foraging habitat. Timing of the behavioral transition from foraging to migration (day of year on the y-axis) is compared to (a) the date of upwelling onset; (b) the date of peak upwelling; and (c) total upwelling accumulated from the spring transition to the end of the upwelling season.
In a similar vein, research by the GEMM Lab on blue whale ecology in New Zealand has linked their vocalizations known as D calls to upwelling conditions, demonstrating that these calls likely reflect blue whale foraging opportunities (Barlow et al. 2021). In ongoing analyses, we are finding that these foraging-related calls were drastically reduced during marine heatwave conditions, which we know altered blue whale distribution in the region (Barlow et al. 2020). Now, for the final component of Dawn’s PhD, she is linking year-round environmental conditions to the occurrence patterns of different blue whale vocalization types, hoping to shed light on ecosystem processes by listening to the signals of these ecosystem sentinels.
A blue whale comes up for air in the South Taranaki Bight of New Zealand. photo by L. Torres.
It is important to understand the widespread implications of the rapidly warming climate and changing ocean conditions on valuable and vulnerable marine ecosystems. The cases explored here in this blog exemplify the importance of monitoring these marine megafauna sentinel species, both now and into the future, as they reflect the health of the ecosystems they inhabit.
Did you enjoy this blog? Want to learn more about marine life, research and conservation? Subscribe to our blog and get a weekly email when we make a new post! Just add your name into the subscribe box on the left panel.
References:
Barlow DR, Bernard KS, Escobar-Flores P, Palacios DM, Torres LG (2020) Links in the trophic chain: Modeling functional relationships between in situ oceanography, krill, and blue whale distribution under different oceanographic regimes. Mar Ecol Prog Ser 642:207–225.
Barlow DR, Klinck H, Ponirakis D, Garvey C, Torres LG (2021) Temporal and spatial lags between wind, coastal upwelling, and blue whale occurrence. Sci Rep 11:1–10.
Becker EA, Forney KA, Redfern J V., Barlow J, Jacox MG, Roberts JJ, Palacios DM (2019) Predicting cetacean abundance and distribution in a changing climate. Divers Distrib 25:626–643.
Cairns DK (1988) Seabirds as indicators of marine food supplies. Biol Oceanogr 5:261–271.
Cavole LM, Demko AM, Diner RE, Giddings A, Koester I, Pagniello CMLS, Paulsen ML, Ramirez-Valdez A, Schwenck SM, Yen NK, Zill ME, Franks PJS (2016) Biological impacts of the 2013–2015 warm-water anomaly in the northeast Pacific: Winners, losers, and the future. Oceanography 29:273–285.
Croll DA, Marinovic B, Benson S, Chavez FP, Black N, Ternullo R, Tershy BR (2005) From wind to whales: Trophic links in a coastal upwelling system. Mar Ecol Prog Ser 289:117–130.
Croll DA, Tershy BR, Hewitt RP, Demer DA, Fiedler PC, Smith SE, Armstrong W, Popp JM, Kiekhefer T, Lopez VR, Urban J, Gendron D (1998) An integrated approch to the foraging ecology of marine birds and mammals. Deep Res Part II Top Stud Oceanogr.
Gavrilchuk K, Lesage V, Fortune SME, Trites AW, Plourde S (2021) Foraging habitat of North Atlantic right whales has declined in the Gulf of St. Lawrence, Canada, and may be insufficient for successful reproduction. Endanger Species Res 44:113–136.
Hazen EL, Abrahms B, Brodie S, Carroll G, Jacox MG, Savoca MS, Scales KL, Sydeman WJ, Bograd SJ (2019) Marine top predators as climate and ecosystem sentinels. Front Ecol Environ 17:565–574.
Hazen EL, Jorgensen S, Rykaczewski RR, Bograd SJ, Foley DG, Jonsen ID, Shaffer SA, Dunne JP, Costa DP, Crowder LB, Block BA (2013) Predicted habitat shifts of Pacific top predators in a changing climate. Nat Clim Chang 3:234–238.
Hyrenbach KD, Forney KA, Dayton PK (2000) Marine protected areas and ocean basin management. Aquat Conserv Mar Freshw Ecosyst 10:437–458.
Meyer-Gutbrod EL, Greene CH, Davies KTA, Johns DG (2021) Ocean regime shift is driving collapse of the north atlantic right whale population. Oceanography 34:22–31.
Oestreich WK, Abrahms B, Mckenna MF, Goldbogen JA, Crowder LB, Ryan JP (2022) Acoustic signature reveals blue whales tune life history transitions to oceanographic conditions. Funct Ecol.
Piatt JF, Parrish JK, Renner HM, Schoen SK, Jones TT, Arimitsu ML, Kuletz KJ, Bodenstein B, Garcia-Reyes M, Duerr RS, Corcoran RM, Kaler RSA, McChesney J, Golightly RT, Coletti HA, Suryan RM, Burgess HK, Lindsey J, Lindquist K, Warzybok PM, Jahncke J, Roletto J, Sydeman WJ (2020) Extreme mortality and reproductive failure of common murres resulting from the northeast Pacific marine heatwave of 2014-2016. PLoS One 15:e0226087.
Scopel LC, Diamond AW, Kress SW, Hards AR, Shannon P (2018) Seabird diets as bioindicators of atlantic herring recruitment and stock size: A new tool for ecosystem-based fisheries management. Can J Fish Aquat Sci.
Silber GK, Lettrich MD, Thomas PO, Baker JD, Baumgartner M, Becker EA, Boveng P, Dick DM, Fiechter J, Forcada J, Forney KA, Griffis RB, Hare JA, Hobday AJ, Howell D, Laidre KL, Mantua N, Quakenbush L, Santora JA, Stafford KM, Spencer P, Stock C, Sydeman W, Van Houtan K, Waples RS (2017) Projecting marine mammal distribution in a changing climate. Front Mar Sci 4:413.
Sydeman WJ, Poloczanska E, Reed TE, Thompson SA (2015) Climate change and marine vertebrates. Science 350:772–777.
Part of what makes being a graduate student so exciting is the way that learning can flip the world around: you learn a new framework or method, and suddenly everything looks a little different. I am experiencing this fabulous phenomenon lately as I learn to collect and process active acoustic data, which can reveal the distribution and biomass of animals in the ocean – including those favored by foraging whales off of Oregon, like the tiny shrimp-like krill.
We know that whales seek out the dense, energy-rich swarms that krill form, and that knowing where to expect krill can give us a leg up in anticipating whale distributions. Project OPAL (Overlap Predictions About Large whales) seeks to model and provide robust predictions of whale distributions off the coast of Oregon, so that managers can make spatially discrete decisions about potential fishery closures, minimizing burdens to fishermen while also maximizing protection of whales. We hope that including prey in our ecosystem models will help this effort, and working on this aim is one of the big tasks of my PhD.
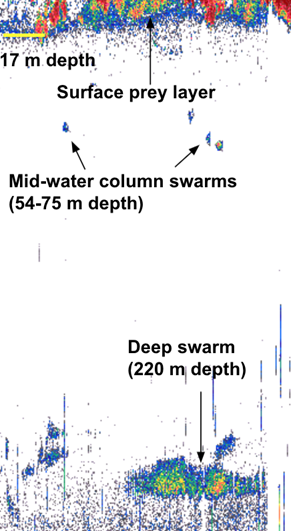
So, how do we know where to expect krill to be off the coast of Oregon? Acoustic tools give us the opportunity to flip the world upside down: we use a tool called an echosounder to eavesdrop on the ocean, yielding visual outputs like the ones below that let us “see” and interpret sound.
Echograms like these reveal features in the ocean that scatter “pings” of sound, and interpreting these signals can show life in the water column.
This is how it works. The echosounder emits pulses of sound at a known frequency, and then it listens for their return after it bounces of the sea floor or things in the water column. Based on sound experiments in the laboratory, we know to expect our krill species, Euphausia pacifica and Thysanoessa spinifera, to return those echoes at a characteristic decibel level. By constantly “pinging” the water column with this sound, we can record a continuous soundscape along the cruise track of a vessel, and analyze it to identify the animals and features recorded.
I had the opportunity to use an echosounder for the first time recently, on the first HALO cruise. We deployed the echosounder soon after sunrise, 65 miles offshore from Newport. After a little fiddling and troubleshooting, I was thrilled to start “listening” to the water; I was able to see the frothy noise at its surface, the contours of the seafloor, and the pixelated patches that indicate prey in between. Although it’s difficult to definitively identify animals only based on the raw output, we saw swarms that looked like our beloved krill, and other aggregations that suggested hake. Sometimes, at the same time that the team of visual observers on the flying bridge of the vessel sighted whales, I also saw potential prey on the echogram.
I spent much of the HALO cruise monitoring incoming data from the transducer on the SIMRAD EK60. Photo: Marissa Garcia.
I’m excited to keep collecting these data, and grateful that I can also access acoustic data collected by others. Many research vessels use echosounders while they are underway, including the NOAA Ship Bell M. Shimada, which conducts cruises in the Northern California Current several times a year. Starting in 2018, GEMM Lab members have joined these cruises to conduct marine mammal surveys.
This awesome pairing of data types means that we can analyze the prey that was available at the time of marine mammal sightings. I’ve been starting to process acoustic data from past Northern California Current cruises, eavesdropping on the preyscape in places that were jam-packed with whales, such as this echogram from the September 2020 cruise, below.
An echogram from the September 2020 NCC cruise shows a great deal of prey at different depths.
Like a lot of science, listening to animals in the sea comes down to occasional bursts of fieldwork followed by a lot of clicking on a computer screen during data analysis. This analysis can be some pretty fun clicking, though – it’s amazing to watch the echogram unfurl, revealing the preyscape in a swath of ocean. I’m excited to keep clicking, and learn what it can tell us about whale distributions off of Oregon.
Clara Bird, PhD Student, OSU Department of Fisheries, Wildlife, and Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
The start of a new school year is always an exciting time. Like high school, it means seeing friends again and the anticipation of preparing to learn something new. Even now, as a grad student less focused on coursework, the start of the academic year involves setting project timelines and goals, most of which include learning. As I’ve been reflecting on these goals, one of my dad’s favorite sayings has been at the forefront of my mind. As an overachieving and perfectionist kid, I often got caught up in the pursuit of perfect grades, so the phrase “just learn the stuff” was my dad’s reminder to focus on what matters. Getting good grades didn’t matter if I wasn’t learning. While my younger self found the phrase rather frustrating, I have come to appreciate and find comfort in it.
Given that my research is focused on behavioral ecology, I’ve also spent a lot of time thinking about how gray whales learn. Learning is important, but also costly. It involves an investment of energy (a physiological cost, Christie & Schrater, 2015; Jaumann et al., 2013), and an investment of time (an opportunity cost). Understanding the costs and benefits of learning can help inform conservation efforts because how an individual learns today affects the knowledge and tactics that the individual will use in the future.
Like humans, individual animals can learn a variety of tactics in a variety of ways. In behavioral ecology we classify the different types of learning based on the teacher’s role (even though they may not be consciously teaching). For example, vertical transmission is a calf learning from its mom, and horizontal transmission is an individual learning from other conspecifics (individuals of the same species) (Sargeant & Mann, 2009). An individual must be careful when choosing who to learn from because not all strategies will be equally efficient. So, it stands to reason than an individual should choose to learn from a successful individual. Signals of success can include factors such as size and age. An individual’s parent is an example of success because they were able to reproduce (Barrett et al., 2017). Learning in a population can be studied by assessing which individuals are learning, who they are learning from, and which learned behaviors become the most common.
An example of such a study is Barrett et al. (2017) where researchers conducted an experiment on capuchin monkeys in Costa Rica. This study centered around the Panama ́fruit, which is extremely difficult to open and there are several documented capuchin foraging tactics for processing and consuming the fruit (Figure 1). For this study, the researchers worked with a group of monkeys who lived in a habitat where the fruit was not found, but the group included several older members who had learned Panamá fruit foraging tactics prior to joining this group. During a 75-day experiment, the researchers placed fruits near the group (while they weren’t looking) and then recorded the tactics used to process the fruit and who used each tactic. Their results showed that the most efficient tactic became the most common tactic over time, and that age-bias was a contributing factor, meaning that individuals were more like to copy older members of the group.
Figure 1. Figure from Barrett et al. (2017) showing a capuchin monkey eating a Panamá fruit using the canine seam technique.
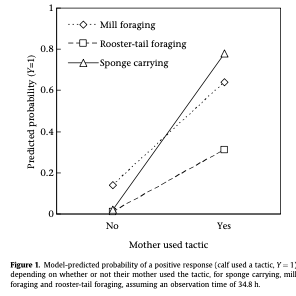
Social learning has also been documented in dolphin societies. A long-term study on wild bottlenose dolphins in Shark Bay, Australia assessed how habitat characteristics and the foraging behaviors used by moms and other conspecifics affected the foraging tactics used by calves (Sargeant & Mann, 2009). Interestingly, although various factors predicted what foraging tactic was used, the dominant factor was vertical transmission where the calf used the tactic learned from its mom (Figure 2). Overall, this study highlights the importance of considering a variety of factors because behavioral diversity and learning are context dependent.
Figure 2. Figure from Sargeant & Mann (2009) showing that the probability of a calf using a tactic was higher if the mother used that tactic.
Social learning is something that I am extremely interested in studying in our study population of gray whales in Oregon. While studies on social learning for such long-lived animals require a longer study period than of the span of our current dataset, I still find it important to consider the role learning may play. One day I would love to delve into the different factors of learning by these gray whales and answer questions such as those addressed in the studies I described above. Which foraging tactics are learned? How much of a factor is vertical transmission? Considering that gray whale calves spend the first few months of the foraging season with their mothers I would expect that there is at least some degree of vertical transmission present. Furthermore, how do environmental conditions affect learning? What tactics are learned in good vs. poor years of prey availability? Does it matter which tactic is learned first? While the chances that I’ll get to address these questions in the next few years are low, I do think that investigating how tactic diversity changes across age groups could be a good place to start. As I’ve discussed in a previous blog, my first dissertation chapter will focus on quantifying the degree of individual specialization present in my study group. After reading about age-biased learning, I am curious to see if older whales, as a group, use fewer tactics and if those tactics are the most energetically efficient.
The importance of understanding learning is related to that of studying individual specialization, which can allows us to estimate how behavioral tactics might change in popularity over time and space. We could then combine this with knowledge of how tactics are related to morphology and habitat and the associated energetic costs of each tactic. This knowledge would allow us to estimate the impacts of environmental change on individuals and the population. While my dissertation research only aims to provide a few puzzle pieces in this very large and complicated gray whale ecology puzzle, I am excited to see what I find. Writing this blog has both inspired new questions and served as a good reminder to be more patient with myself because I am still, “just learning the stuff”.
Returning to a place you once lived always shows how much you and the world around you have changed, offering a new perspective on the time away and where you are now. I’m writing this from my old office at Bigelow Laboratory for Ocean Sciences in East Boothbay, Maine, where I worked before moving out to Oregon to join the GEMM Lab and start graduate school at OSU. Being back in Maine has made me reflect on how much I’ve learned over the last year, and given me the opportunity to think about what’s ahead.
As a science communications specialist at Bigelow for three years, much of my work involved quickly getting up to speed on new research and writing articles for a general audience about important ocean processes. My first year of grad school has both deepened and broadened my perspective on the ocean, prodding me to think at telescoping temporal and spatial scales. I can tell that I think about the ocean differently now.
In 2019, writing this feature-length article about impacts of changing climate on North Atlantic right whales and their prey was my first introduction to research using environmental models to help mitigate entanglement issues. Now, I’m excited to be pursuing research with these themes as part of the GEMM Lab’s Project OPAL.
Over the last year, my coursework in ocean ecology and biogeochemistry surveyed the physical and chemical workings of the ocean, marine ecosystem dynamics, and the global cycles that control much of life on earth. Through lab activities and fieldwork, I began learning about whales and the marine system off the coast of Oregon, and how to ask questions that occupy the intersection between whales and their environment.
This work and learning have made me think in a new way about whales as agents of biogeochemical cycling: how do they shuttle nutrients across large distances and affect global cycles? In what ways is the biogeography of whales an expression of the global patterns of light availability and nutrient fluxes that support their prey? How is it possible to detangle and encapsulate all of the relevant variability of a natural system into a mathematical model?
All these questions were churning in my mind at the start of this trip, as I spent the bus ride from Boston to Maine reading papers for our monthly GEMM lab meeting. I also remembered the first meeting that I joined, when I was so intimidated that I couldn’t imagine discussing research with this impressive group. This time, I was just as in awe as ever of the lab, but a bit more confident in wielding acronyms and sharing ideas.
I actually attended my first GEMM Lab meeting while still working in Maine, in July 2020. I was joined by my friends’ one-year-old daughter, who alternately tried to chime in on the meeting and shut my laptop. Now, she is a chatty two-year-old kid and newly a big sister. The new baby became part of my PhD this week too, snoozing in my lap as I edited an abstract.
Only 16 days old and already helping write an abstract!
Often, it’s only seeing my friends’ children grow that shows me how much time has passed. This time, I can feel it in myself, as well. I’m excited to have made it through the first year of coursework and to be learning to formulate research questions and think about ocean systems in new ways. I’m happy to be back in this place that inspired me to pursue a PhD, and to be able to share my own work and knowledge with former colleagues.
I gained so much during my time here at Bigelow: the communication and outreach skills in my job, inspiration from the scientific curiosity and passion of my colleagues, and the support of all these people who reassured me that I would get into grad school and that doing a PhD is a good idea. I’m so happy to be able to carry this support and momentum forward with me through the rest of grad school, and excited to return to Oregon and keep going.
About 10 months have passed since I started working on OPAL, a project that aims to identify the co-occurrence between whales and fishing effort in Oregon to reduce entanglement risk. During this period, you would be surprised to know how little ecology I have actually done and how much time has been devoted to data processing! I compiled several million GPS trackline positions, processed hundreds of marine mammal observations, wrote several thousand lines of R code, downloaded and extracted a couple Gb of environmental data… before finally reaching the modeling phase of the OPAL project. And with it, finally comes the time to look more closely at the ecology and behavior of my species of interest. While the previous steps of the project were pretty much devoid of ecological reasoning, the literature homework now comes in handy to guide my choices regarding habitat use models, such as selecting environmental predictors of whale occurrence, deciding on what seasons should be modeled, and choosing the spatio-temporal scale at which the data should be aggregated.
Whale diversity on the US west coast
The productive waters off the US west coast host a great diversity of cetaceans. Eight species of baleen whales are reported to occur there by NOAA fisheries: blue whales, Bryde’s whales, fin whales, gray whales, humpback whales, minke whales, North Pacific right whales and sei whales. Among them, no less than five are listed as Endangered under the Endangered Species Act. Whether they are only passing by or spending months feeding in the region, the timing and location where these animals are observed varies greatly by species and by population.
During the 113 hours of aerial survey effort and 264 hours of boat-based search conducted for the OPAL project, 563 groups of baleen whales have been observed to-date (up to mid-May 2021 to be exact… more data coming soon!). Among the observations where animals could be identified to the species level, humpback whales are preponderant, as they represent about half of the whale groups observed (n = 293). Blue (n = 41) and gray whales (n = 46) come next, the latter being observed in more nearshore waters. Finally, a few fin whale groups were observed (n = 28). The other baleen whale species reported by NOAA in the US west coast species list were very rarely or not observed at all during OPAL surveys.
The OPAL aerial surveys conducted in partnership with the United States Coast Guard (USCG) were specifically designed to study whales occurring on the continental shelf along the coast of Oregon. Hence, most of this survey effort is located in waters from 800 m to 30 m deep, which may explain the relatively low number of gray whales detected. Indeed, gray whales observed in Oregon may either be migrating along the coast to and from their breeding grounds in Baja California, or be part of the small Pacific Coast Feeding Group that forage in Oregon nearshore and shallow waters during the summer. This group of whales is one the main GEMM lab’s research focus, being at the core of no less than three ongoing research projects: AMBER, GRANITE, and TOPAZ.
So today, let’s turn our eyes to the sea horizon and talk about some other members of the baleen whale community: rorquals. Conveniently, the three species of baleen whales (gray whales aside) most commonly observed during OPAL surveys are all part of the rorqual family, a.k.a Balaenopteridae: humpback whales, blue whales and fin whales (Figure 1). They are morphologically characterized by the pleated throat grooves that allow them to engulf large quantities of food and water, for instance when lunge-feeding. Known cases of hybridization between these three species demonstrate their close relatedness (Jefferson et al., 2021). They all have worldwide distributions and display unequally understood migratory behaviors, seasonally traveling between warm tropical breeding grounds and temperate-polar feeding grounds. They occur in great numbers in productive waters such as the upwelling system of the California Current.
The three accomplices
Figure 1: Aerial view of three rorquals species: a humpback whale (left), a fin whale (center), and a blue whale (right). Photo credit: Leigh Torres and Craig Hayslip. Photos taken off the Oregon coast under NOAA/NMFS permit during USCG helicopter flights conducted as part of the OPAL project
Humpback whales (Megaptera novaeangliae) are easily differentiated from other rorquals because of their long pectoral fins (up to one third of their body length!), which inspired their scientific name, Megaptera, « big-winged » (Figure 1). Individuals observed in Oregon mostly belong to a mix of two Distinct Population Segments (DPS): the threatened Mexico and endangered Central American DPS. Although humpback whales from different DPS do not show any morphological differences, they are genetically distinct because they have been mating separately in distinct breeding grounds for generations and generations. This genetic differentiation has great implications in terms of conservation since the Central American DPS is recovering at a lesser rate than the Mexican and is therefore subject to different management measures (recovery plan, monitoring plan, designated critical habitats). Humpback whales migrate and feed off the US west coast, with a peak in abundance in the mid to late summer. Compared to other rorquals that are found in the open ocean, humpback whales are mostly observed on the continental shelf (Becker et al., 2019). They are considered to have a relatively generalist diet, as they feed on a mix of krill (Euphausiids) and fishes (e.g. anchovy, sardines) and are capable of switching their feeding behavior depending on relative prey availability (Fleming, Clark, Calambokidis, & Barlow, 2016; Fossette et al., 2017).
Blue whales (Balaenoptera musculus) are the largest animals ever known (max length 33 m, Jefferson et al., 2008), and sadly the most at risk of global extinction among our three species of interest (listed as « endangered » in the IUCN red list). They have a distinctive mottled blue and light gray skin, a slender body and a broad U-shaped head (or as some say « like a gothic arch », Figure 1). Blue whales tend to be open ocean animals, but they regroup seasonally to feed in highly productive nearshore areas such as the Southern California Bight (Becker et al. 2019, Abrahms et al. 2019). Blue whales migrating or feeding along the US west coast belong to the Eastern North Pacific stock and are subject to great research and conservation efforts. Contrary to their other rorqual counterparts, blue whales are quite picky eaters, as they exclusively feed on krill. This difference in diet leads to resource partitioning facilitating rorqual coexistence in the California Current (Fossette et al., 2017). These differences in feeding strategies have important implications for designing predictive models of habitat use.
Fin whales (Balaenoptera physalus) are nicknamed « greyhounds of the sea » due to their exceptional swim speed (max 46 km/h). They are a little smaller than blue whales (max length 27 m, Jefferson, Webber, & Pitman, 2008) but share a similar sleek and streamlined shape. Their coloration is their most distinctive feature: the left lower jaw being mostly dark while the right is white. V-shaped light-gray « chevrons » color their back, behind the head (Figure 1). The California/Oregon/Washington is one of the three stocks recognized in the North Pacific (NOAA Fisheries, 2018). Within this region, there is genetic evidence for a geographic separation north and south of Point Conception, CA (Archer et al., 2013). Like other rorquals, they are migratory, but their seasonal distribution is relatively less well understood as they appear to spend a lot of time in open oceans. For instance, a meta-analysis for the North Pacific found little evidence for fin whales using distinct calving areas (Mizroch, Rice, Zwiefelhofer, Waite, & Perryman, 2009). In the California Current System, satellite tracking has provided great insights into their space-use patterns. In the Southern California Bight, fin whales show year-round residency and seasonal shifts in habitat use as they move further offshore and north during the spring/summer (Scales et al., 2017). The Northern California Current offshore waters appeared to be used during the summer months by the whales tagged in the Southern California Bight. Yet, fin whales are observed year-round in Oregon (NOAA Fisheries, 2018).
Towards predictive models of rorqual distribution
Enough observations have now been collected as part of the OPAL project to be able to model the habitat use of some of these rorqual species. Based on 12 topographic (i.e., depth, slope, distance to canyons) and physical variables (temperature, chlorophyll-a, water column stratification, etc.), I have made my first attempt at predicting seasonal distribution patterns of humpback whales and blue whales in Oregon. These models will be improved in the coming months, with more data pouring in and refined parametrizations, but they already bring insights into the shared habitat use patterns of these species, as well as their specificities.
Across multiple cross-validations of the species-specific models, sea surface temperature, sea surface height and depth were recurrently selected among the most important variables influencing both humpback and blue whale distributions. Predicted densities of blue whales were relatively higher at less than 40 fathoms compared to humpback whales, although both species’ hotspots were located outside this newly implemented seasonal fishing limit (Figure 2). Higher densities were generally predicted off Newport and Port Orford, and north of North Bend.
Figure 2: Predicted densities of humpback and blue whales during the month of September 2018, 2019, and 2020 in Oregon waters (OPAL project). Core areas of use (predicted densities in the top 25%) are represented, with darker shades of blue and orange showing higher predicted densities. Dashed lines represent the tracklines followed by USCG monthly aerial surveys. The black line represents the 40 fathom isobath. Grey boxes overlayed on predictions delineate the areas of extrapolation where environmental conditions are non-analogous to the conditions in which the models were trained. Disclaimer: these model outputs are preliminary and should be interpreted with caution.
Once our rorqual models are finalized, we will work with our partners at the Oregon Department of Fisheries and Wildlife to overlay predicted whale hotspots with areas of high crab pot densities. This overlap analysis will help us understand the times and places where co-occurrence of suitable whale habitat and fishing activities put whales at risk of entanglement.
Becker, E. A., Forney, K. A., Redfern, J. V, Barlow, J., Jacox, M. G., Roberts, J. J., & Palacios, D. M. (2019). Predicting cetacean abundance and distribution in a changing climate. Diversity and Distributions, 25(4), 626–643. https://doi.org/10.1111/ddi.12867
Fleming, A. H., Clark, C. T., Calambokidis, J., & Barlow, J. (2016). Humpback whale diets respond to variance in ocean climate and ecosystem conditions in the California Current. Global Change Biology, 22, 1214–1224. https://doi.org/10.1111/gcb.13171
Fossette, S., Abrahms, B., Hazen, E. L., Bograd, S. J., Zilliacus, K. M., Calambokidis, J., … Croll, D. A. (2017). Resource partitioning facilitates coexistence in sympatric cetaceans in the California Current. Ecology and Evolution, 7, 9085–9097. https://doi.org/10.1002/ece3.3409
Jefferson, T. A., Palacios, D. M., Clambokidis, J., Baker, S. C., Hayslip, C. E., Jones, P. A., … Schulman-Janiger, A. (2021). Sightings and Satellite Tracking of a Blue / Fin Whale Hybrid in its Wintering and Summering Ranges in the Eastern North Pacific. Advances in Oceanography & Marine Biology, 2(4), 1–9. https://doi.org/10.33552/AOMB.2021.02.000545
Jefferson, T. A., Webber, M. A., & Pitman, R. L. (2008). Marine Mammals of the World. A comprehensive guide to their identification. Elsevier, London, UK.
Mizroch, S. A., Rice, D. W., Zwiefelhofer, D., Waite, J., & Perryman, W. L. (2009). Distribution and movements of fin whales in the North Pacific Ocean. Mammal Review, 39(3), 193–227. https://doi.org/10.1111/j.1365-2907.2009.00147.x
NOAA Fisheries. (2018). Fin whale stock assessment report ( Balaenoptera physalus physalus ): California / Oregon / Washington Stock.
Scales, K. L., Schorr, G. S., Hazen, E. L., Bograd, S. J., Miller, P. I., Andrews, R. D., … Falcone, E. A. (2017). Should I stay or should I go? Modelling year-round habitat suitability and drivers of residency for fin whales in the California Current. Diversity and Distributions, 23(10), 1204–1215. https://doi.org/10.1111/ddi.12611
Hello from the RV Bell M. Shimada! We are currently sampling at an inshore station on the Heceta Head Line, which begins just south of Newport and heads out 45 nautical miles west into the Pacific Ocean. We’ll spend 10 days total at sea, which have so far been full of great weather, long days of observing, and lots of whales.
Dawn and Rachel in matching, many-layered outfits, 125 miles offshore on the flying bridge of the RV Bell M. Shimada.
Run by NOAA, this Northern California Current (NCC) cruise takes place three times per year. It is fabulously interdisciplinary, with teams concurrently conducting research on phytoplankton, zooplankton, seabirds, and more. The GEMM Lab will use the whale survey, krill, and oceanographic data to fuel species distribution models as part of Project OPAL. I’ll be working with this data for my PhD, and it’s great to be getting to know the region, study system, and sampling processes.
I’ve been to sea a number of times and always really enjoyed it, but this is my first time as part of a marine mammal survey. The type and timing of this work is so different from the many other types of oceanographic science that take place on a typical research cruise. While everyone else is scurrying around, deploying instruments and collecting samples at a “station” (a geographic waypoint in the ocean that is sampled repeatedly over time), we – the marine mammal team – are taking a break because we can only survey when the boat is moving. While everyone else is sleeping or relaxing during a long transit between stations, we’re hard at work up on the flying bridge of the ship, scanning the horizon for animals.
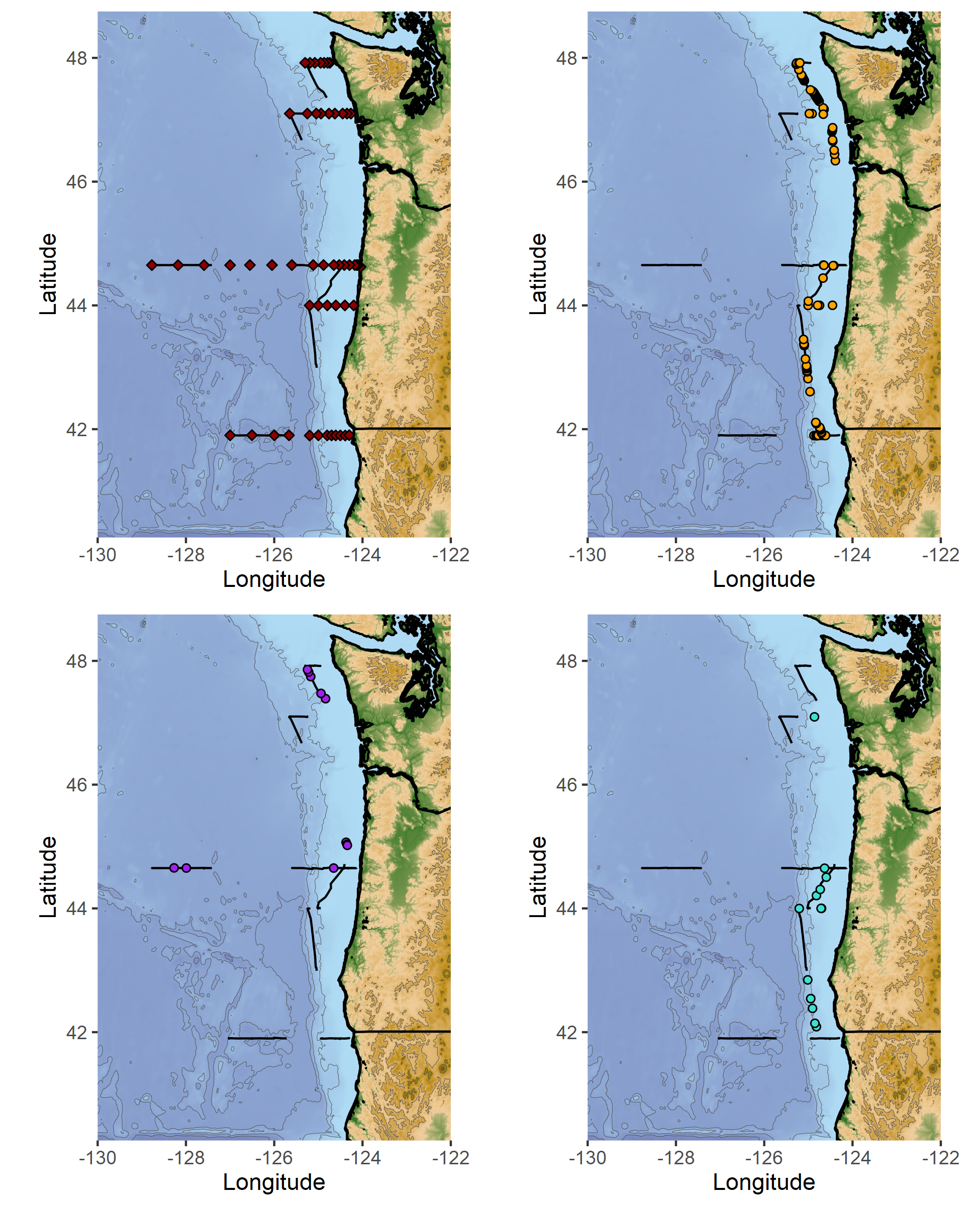
Top left: marine mammal survey effort (black lines), and oceanographic sampling stations (red diamonds). Top right: humpback whale sighting locations. Bottom left: fin whale sighting locations. Bottom right: pacific white-sided dolphin sighting locations.
During each “on effort” survey period, Dawn and I cover separate quadrants of ocean, each manning either the port or starboard side. We continuously scan the horizon for signs of whale blows or bodies, alternating between our eyes and binoculars. During long transits, we work in chunks – forty minutes on effort, and twenty minutes off effort. Staring at the sea all day is surprisingly tiring, and so our breaks often involve “going to the eye spa,” which entails pulling a neck gaiter or hat over your eyes and basking in the darkness.
Dawn has been joining these NCC cruises for the last four years, and her wealth of knowledge has been a great resource as I learn how to survey and identify marine mammals. Beyond learning the telltale signs of separate species, one of the biggest challenges has been learning how to read the sea better, to judge the difference between a frothy whitecap and a whale blow, or a distant dark wavelet and a dorsal fin. Other times, when conditions are amazing and it feels like we’re surrounded by whales, the trick is to try to predict the positions and trajectory of each whale so we don’t double-count them.
Over the last week, all our scanning has been amply rewarded. We’ve seen pods of dolphins play in our wake, and spotted Dall’s porpoises bounding alongside the ship. Here on the Heceta Line, we’ve seen a diversity of pinnipeds, including Northern fur seals, Stellar sea lions, and California sea lions. We’ve been surprised by several groups of fin whales, farther offshore than expected, and traveled alongside a pod of about 12 orcas for several minutes, which is exactly as magical as it sounds.
Killer whales traveling alongside the Bell M. Shimada, putting on a show for the NCC science team and ship crew. Photo by Dawn Barlow.
Notably, we’ve also seen dozens of humpbacks, including along what Dawn termed “the humpback highway” during our transit offshore of southern Oregon. One humpback put on a huge show just 200 meters from the ship, demonstrating fluke slapping behavior for several minutes. We wanted to be sure that everyone onboard could see the spectacle, so we radioed the news to the bridge, where the officers control the ship. They responded with my new favorite radio call ever: “Roger that, we are currently enamored.”
A group of humpbacks traveling along the humpback highway. Photo by Dawn Barlow.A humpback whale fluke slapping. Photo by Dawn Barlow.
Even with long days and tired eyes, we are still constantly enamored as well. It has been such a rewarding cruise so far, and it’s hard to think of returning back to “real life” next week. For now, we’re wishing you the same things we’re enjoying – great weather, unlimited coffee, and lots of whales!
By Alejandro Fernandez Ajo, PhD student in the Department of Biology, Northern Arizona University, visiting scientist in the GEMM Lab working on the gray whale physiology and ecology project
When a cetacean (whales and dolphins) is ashore or trapped in nearshore waters and cannot return to the open waters, it is considered stranded. Frequently, the stranded animal is in distress, dying, or dead. Although rare, the stranded cetacean can be a healthy animal trapped due to changes in tide or disorientation. Every year many cetacean strandings are reported from along the coasts around the world, and likely many more stranding events go unnoticed when they occur in remote areas. In all cases, the question is: why do cetaceans beach?
Southern right whales stranded at the coast of Peninsula Valdés, Patagonia-Argentina. Photo: Matias DiMartino / Southern Right Whale Health Monitoring Program.
There may be different causes for whales and dolphins to strand on beaches, either dead or alive. Understanding and investigating the causes of cetaceans strandings is critical because they can be indicators of ocean health, can help identify anthropogenic sources of disturbance, and can give insights into larger environmental issues that may also have implications for human health (NOAA). In this context, when scientists are analyzing a stranding event, they consider both possibilities that the event was natural or human-caused and classify strandings according to specific characteristics to study the causes of these events.
Types of cetacean strandings:
Live or Dead Stranding:
A stranding can involve live animals or dead animals if the death occurs in the sea and the body is thrown ashore by wind or currents. In live strandings, when they occur near urbanized areas, usually significant efforts are made to rescue and return the animals to the water; with small odontocetes, sometimes there is success, and animals can be rescued. However, when large whales are beached alive, their own weight out of the water can compress their organs and can cause irreversible internal damage. Although not externally visible, such damage can sometimes cause the death of the animal even after returning to the sea.
According to the number of individuals:
Single strandings occur when only a single specimen is affected at the time. The cetaceans that most frequently strand individually are the baleen (or mysticete) whales, such as right and humpback whales, due to their often solitary habits.
Mass strandings comprise two or more specimens, and in some cases, it can involve tens or even a few hundred animals. The mass strandings are more frequently observed for the odontocetes, such as pilot whales, false killer whales, and sperm whales with more complex social structures and gregarious habits.
Left: Single southern right whale calf stranded at the coast of Peninsula Valdés, Patagonia-Argentina. Ph.: Mariano Sironi / ICB. Right: Mass stranding of common dolphins in Patagonia-Argentina. Photo: www.elpais.com
Unusual Mortality Events
The Marine Mammal Protection Act defines an unusual mortality event (UME) as a stranding event that is unexpected, involves a significant die-off of any marine mammal population, and demands immediate response. Seven criteria make a mortality event “unusual.” Source: https://www.fisheries.noaa.gov.
A marked increase in the magnitude or a marked change in morbidity, mortality, or strandings when compared with prior records.
A temporal change in morbidity, mortality, or strandings is occurring.
A spatial change in morbidity, mortality, or strandings is occurring.
The species, age, or sex composition of the affected animals is different than that of animals that are normally affected.
Affected animals exhibit similar or unusual pathologic findings, behavior patterns, clinical signs, or general physical condition (e.g., blubber thickness).
Potentially significant morbidity, mortality, or stranding is observed in species, stocks, or populations that are particularly vulnerable (e.g., listed as depleted, threatened, or endangered, or declining). For example, stranding of three or four right whales may be cause for great concern, whereas stranding of a similar number of fin whales may not.
Morbidity is observed concurrent with or as part of an unexplained continual decline of a marine mammal population, stock, or species.
The purpose of the classification of a mortality event as a UME is to activate an emergency response that aims to minimize deaths, determine the event cause, or causes, determine the effect of the event on the population, and identify the role of environmental parameters in the event. Such classification authorizes a federal investigation that is led by the expertise of the Working Group on Marine Mammal Unusual Mortality Events to investigate the event. This working group is comprised of experts from scientific and academic institutions, conservation organizations, and state and federal agencies, all of whom work closely with stranding networks and have a wide variety of experience in biology, toxicology, pathology, ecology, and epidemiology.
Southern right whale necropsy and external measurements. Source: Southern Right Whale Health Monitoring Program / ICB.
What can be learned from strandings and UMEs?
Examining stranded marine mammals can provide valuable insight into marine mammal health and identify environmental factors leading to strandings. Through forensic examinations, the aim is to identify possible risks to whales’ health and evaluate their susceptibility to diseases, pollutants, and other stressors. This information can contribute to cetacean conservation through informed management strategies. However, the quality of the data derived from a necropsy (the postmortem examination of carcasses) is highly contingent upon how early the stranding event is reported. As soon as the animal is deceased, decomposition starts, hindering the possibilities of detailed investigations of the cause of death.
Therefore, a solid network that can report and respond quickly to a stranding event is fundamental; this includes trained personnel, infrastructure, funding, and expertise to respond in a manner that provides for animal welfare (in the case of live strandings) and obtains data on marine mammal health and causes of death. Moreover, a coordinated international organization that integrates national marine mammal stranding networks has also been identifying as a critical aspect to enable adequate response to such mortality events. In many locations and countries around the world, funding, logistical support, and training remain challenging to stranding response.
In response to these concerns and needs, at the last World Marine Mammal Conference, which took place in Barcelona in December of 2019, The Global Stranding Network was founded to “enhance and strengthen international collaboration to (1) ensure consistent, high-quality response to stranded marine mammals globally, and (2) support conservation efforts for species under threat of extinction.” Monitoring marine mammal health worldwide can guide conservation and help identify priority areas for management (Gulland and Stockin, 2020).
What to do in case of finding a whale or dolphin on the beach?
When strandings occur, it is essential to know how to act. Unfortunately, untrained people, often with good intentions, can worsen the situation of stress and injury to the animal or can put themselves at risk of injury or exposure to pathogens. If you find a cetacean alive or dead on the beach, the most important things to do are:
Record information about the location and the animal´s characteristics (the species, if known; the animal’s approximate size; and status (alive or dead)).
Keep at a safe distance: the animal may appear dead to the naked eye and not be. It is important to remember that cetaceans are wild animals and that in stressful situations such as strandings, they can try to defend themselves.
Do not touch the animal: one of the causes of strandings is diseases; therefore, it is advisable not to contact the individuals to avoid exposure to potential pathogens.
If the animal is alive, keep a distance from the animal, especially from its head and tail. Prevent children or dogs from approaching the animal.
Keep calm and do not make noise that could disturb the stranded animal.
Do not take the animal out of the water if it is on the shore or return it to the sea if it is on the beach: Such movement could cause serious injuries, or even death.
Do not feed the animal or give it water: keep the blowhole clear because it is where they breathe.
Proceedings of the workshop “Harmonizing Global Stranding Response.” (2020) World marine mammal Conference Barcelona, Catalonia, Spain. Editors: Gulland F and Stockin K; Ecs Special Publication Series No. 62.
Mazzariol S., Siebert U., Scheinin A., Deaville R., Brownlow A., Uhart M.., Marcondes M., Hernandez G., Stimmelmayr R., Rowles T., Moore K., Gulland F., Meyer M., Grover D., Lindsay P., Chansue N., Stockin K. (2020). Summary of Unusual Cetaceans Strandings Events worldwide (2018-2020). SC-68B/E/09 Rev1.
What I mean is that the vastness of the ocean is very hard to mentally visualize. When facing a conservation issue such as increased whale entanglement along the US West Coast (see OPAL project ), a tempting solution may be to suggest « let’s go see where the whales are and report their location to the fishermen?! ». But, it only takes a little calculation to realize how impractical this idea is.
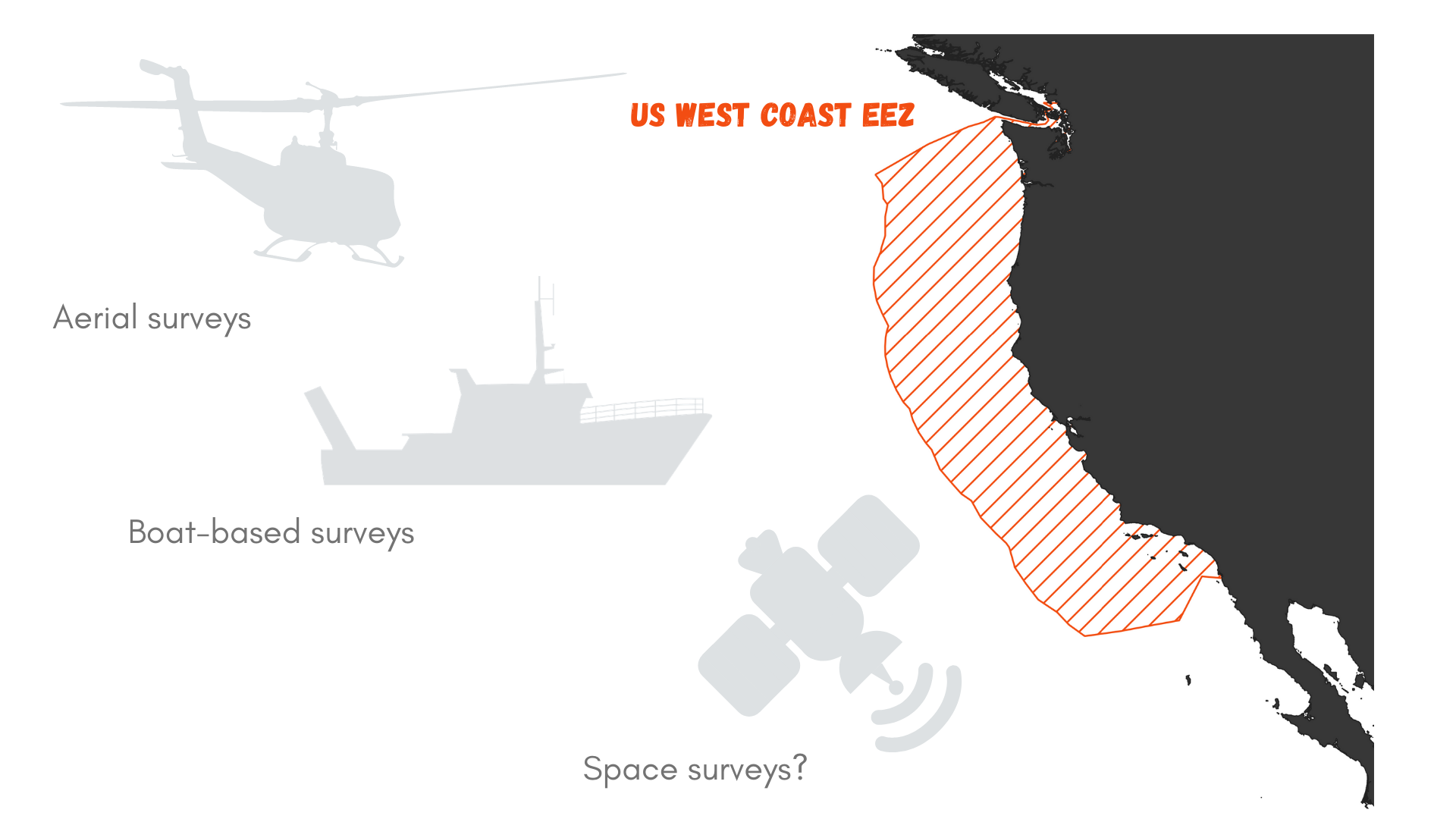
Let’s roll out the numbers. The US West Coast exclusive economic zone (EEZ) stretches from the coast out to 200 nautical miles offshore, as prescribed by the 1982 United Nations Convention on the Law of the Sea. It covers an area of 825,549 km² (Figure 1). Now, imagine that you wish to survey this area for marine mammals. Using a vessel such as the R/V Bell M. Shimada that is used for the Northern California Current Ecosystem surveys cruises (NCC cruises, see Dawn and Rachel’s last blog), we may detect whales at a distance of roughly 6 km (based on my preliminary results). This distance of detection depends on the height of the observer, hence the height of the flying bridge where she/he is standing (the observer’s height may also be accounted for, but unless she/he is a professional basket-ball player, I think it can be neglected here). The Shimada is quite a large ship and it’s flying bridge is 13 meters above the water. Two observers may survey the water on each side of the trackline.
Considering that the vessel is moving at 8 knots (~15 km/h), we may expect to be effectively surveying 180 km² per hour (6x2x15). That’s not too bad, right?
Again, perspective is the key. If we divide the West Coast EEZ surface by 180 km² we can estimate that it would take 2,752 hours to survey this entire region. With an average of 12 hours of daylight, this takes us to…
382 DAYS OF SURVEY, searching for marine mammals over the US West Coast. Considering that observations cannot be undertaken on days with bad weather (fog, heavy rain, strong winds…), it might take more than a year and a half to complete the survey! And what would the marine mammals have done in the meantime? Move…
This little math exercise proves that exhaustively searching for the needle in the haystack from a vessel is not the way to go if we are to describe whale distribution and help mitigate the risk of entanglement. And using another platform of observation is not necessarily the solution. The OPAL project has relied on a great collaboration with the United States Coast Guard to survey Oregon waters. The USCG helicopters travel fast compared to a vessel, about 90 knots (167 km/h). As a result, more ground is covered but the speed at which it is traveling prevents the observer from detecting whales that are very far away. Based on the last analysis I ran for the OPAL project, whales are usually detected up to 3 km from the helicopter (only 5 % of sightings exceed that distance). In addition, the helicopter generally only has capacity for one observer at a time.
If we replicate the survey time calculation from above for the USCG helicopter, we realize that even with a fast-moving aerial survey platform it would still take 137 days to cover the West Coast EEZ.
Figure 1. What is the best survey method to document marine mammal occurrence in the US West Coast Exclusive Economic zone (EEZ)?
First, we can model and extrapolate. This approach is the path we are taking with the OPAL project: we survey Oregon waters in 4 different areas along the coast each month, then model observed whale densities as a function of topographic and oceanographic variables, and then predict whale probability of presence over the entire region. These predictions are based on the assumption that our survey design effectively sampled the variety of environmental conditions experienced by whales over the study region, which it certainly did considering that all sites are surveyed year-round.
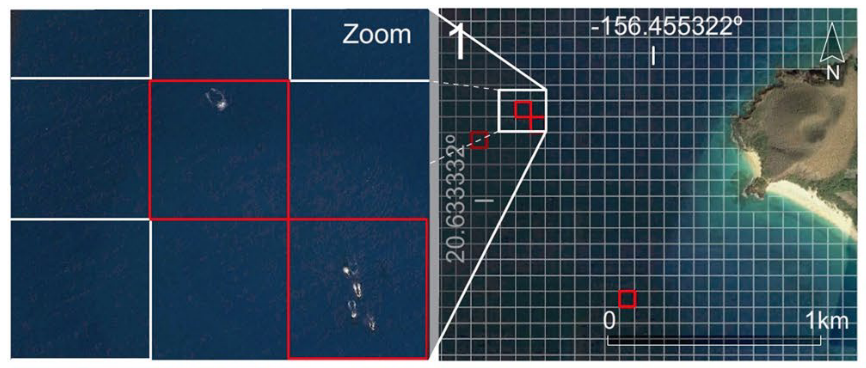
An alternative approach that has been recently discussed in the GEMM Llab, is the use of satellite images to detect whales along the coast. A communication entitled « The Potential of Satellite Imagery for Surveying Whales » was published last month in the Sensors Journal (Höschle et al., 2021) and presents the opportunities offered by this relatively new technology. The WorldView-3 satellite, owned by the company Digitalglobe and launched in 2016, has made it possible to commercialize imagery with a resolution never reached before, of the order of 30 cm per pixel. These very high resolution (VHR) satellite images make it possible to identify several species of large whales (Cubaynes et al. al., 2019) and to estimate their density (Bamford et al., 2020). Furthermore, machine learning algorithms, such as Neural Networks, have proved quite efficient at automatically detecting whales in satellite images (Guirado et al., 2019, Figure 2). While several new ultra-high resolution imaging satellites are expected to be launched in 2021 (by Maxar Technologies and Airbus), this “remote” approach looks like a promising avenue to detect whales over vast regions while drinking a cup of coffee at the office.
Figure 2. Illustration of a whale detection algorithm working on a gridded satellite image (DigitalGlobe). Source: Guirado et al., 2019.
But like any other data collection method, satellites have their drawbacks. We recently discovered that these VHR satellites are routinely switched off while passing above the ocean. Specific inquiries would need to be made to acquire data over our study areas, which would be at great expense. One of the cheapest provider I found is the Soar platform, that provides images at 50 cm resolution in partnership with the Chinese Aerospace Science and Technology Corporation. They advertise daily images anywhere on earth at $10 USD per km². This might sound cheap at first glance, but circling back to our US West Coast EEZ area calculations, we estimate that surveying this region entirely with satellite imagery would cost more than $8 million USD.
Yet, we have to look forward. The use of satellite imagery is likely to broaden and increase in the coming years, with a possible decrease in cost. Quoting Höschle et al. (2021) ‘To protect our world’s oceans, we need a global effort and we need to create opportunities for that to happen’.
Will satellites soon save whales?
References
Bamford, C. C. G. et al. A comparison of baleen whale density estimates derived from overlapping satellite imagery and a shipborne survey. Sci. Rep. 10, 1–12 (2020).
Cubaynes, H. C., Fretwell, P. T., Bamford, C., Gerrish, L. & Jackson, J. A. Whales from space: Four mysticete species described using new VHR satellite imagery. Mar. Mammal Sci. 35, 466–491 (2019).
Guirado, E., Tabik, S., Rivas, M. L., Alcaraz-Segura, D. & Herrera, F. Whale counting in satellite and aerial images with deep learning. Sci. Rep. 9, 1–12 (2019).
Höschle, C., Cubaynes, H. C., Clarke, P. J., Humphries, G. & Borowicz, A. The potential of satellite imagery for surveying whales. Sensors 21, 1–6 (2021).