Eden Van Maren, Homeschool Student from Brookings, TOPAZ/JASPER High School Intern
Hey! I’m Eden Van Maren, an upcoming high school senior from Brookings. I am homeschooled and am taking electives at Brookings Harbor High School.
Growing up in rural Oregon, the outdoors have always been more than just my backyard. It’s been both my classroom and my playground. When Oceana (the other high school intern) and I were homeschooled together as children, Fridays meant her mom would take us up the Chetco River. One Friday, we took our snorkels to observe mature salmon migrating upstream. I remember being so amazed by the size and quantity of the salmon, my young brain could not understand why such large fish would want to swim up to such a small area to lay eggs. The next year when we returned to try and see if the salmon would swim upstream again, we found only one salmon swimming around. This river became my classroom, planting my initial interest in science.
However. Let’s be clear: Being outside in nature was never “all work, no play” – Definitely lots of play! Summers were filled with sunsets on the beach, some foggy day hikes, but most importantly kayaking on the river. I have many fun memories of waking up early on a weekend to pack food for a long day of kayaking in a tandem with my dad and a bunch of other friends. As I’ve gotten older, my passion for both the environment and science have only grown.

Fig 1: My dad and I kayaking with my dog on the Chetco River.
After going on a college tour at the University of Oregon in January, I suddenly started thinking that I should begin planning for college and future career options. On that tour, I met Ma’yet, the Youth Program Education Coordinator at Curry Watershed Partnership who had worked with Allison. Ma’yet was familiar with the TOPAZ/JASPER program run by the GEMM Lab and, while we were discussing possible summer opportunities in science, they suggested that I would be a great fit.
In early March, when I discovered there would be someone from the program coming to present at school, I had already been scheduled to work a shift at Dutch Bros. I managed last minute to have one of my coworkers cover the last few hours of my shift so I would be able to get there. He arrived late, so I ran to get there on time, but I made it! Upon arriving, I sat down for the presentation, and, within minutes, Allison confirmed my desire to be a part of this program. I always knew that science is where I wanted to focus my studies. When I came across this program, I was very interested because it involved exciting outdoor activities while learning and experiencing scientific field work. I was thrilled to meet Allison in person to ask questions and share my enthusiasm about the project.
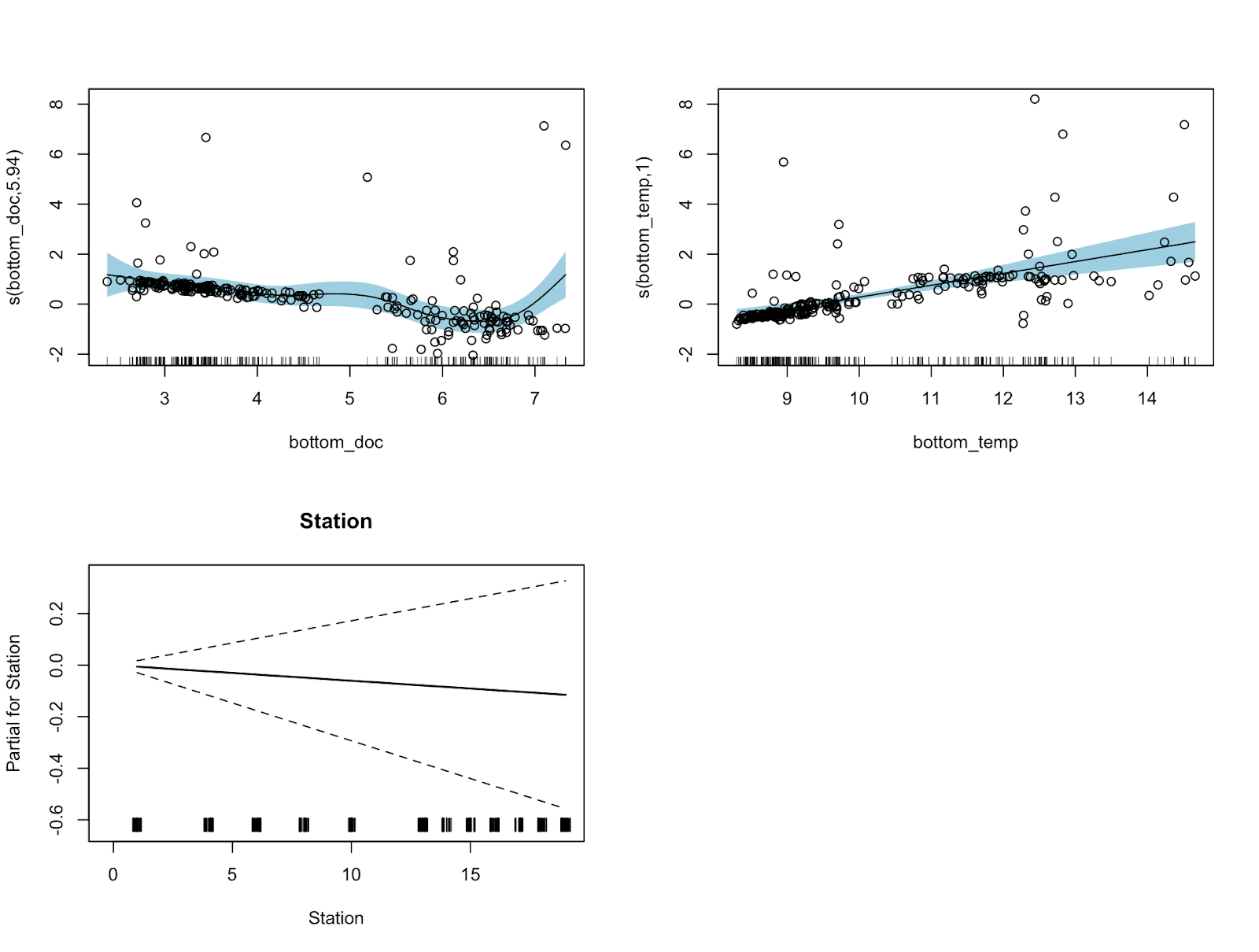
Before working at the Port Orford field station, I had never given much thought to zooplankton. I had known they were the primary prey for whales, but other than that, I hadn’t considered that there was much else to think about. After starting the work associated with zooplankton on this project, I learned through Sophia how zooplankton can be affected by water temperature and kelp abundance, among other things. Along with learning more about zooplankton ecology, part of the program includes collecting zooplankton samples from 12 different stations (using a kayak) out along the Port Orford coastal area. On my very first training day of zooplankton sampling, I pulled up a ridiculous number of zooplankton in the net (much more than the last few seasons).

Fig 2: Me pulling up my first net of zoops! Look at all that zoop!
Once we return to the lab after a morning of zoop collection, we observe these samples under a microscope, identify their species, and count how many species we collected from each station. Just two weeks into our data collection, we have collected 4291 individual zoops, which already surpasses the total amount of zooplankton collected in 2023 and 2022 combined! That’s a lot of zoops! But how do we do it?
In our team, I am considered the zoop expert, but I couldn’t do it without a handful of Welch’s fruit snacks and my playlist full of bangers. Zoop processing can be very tedious, but I really enjoy the peace that comes with finishing a giant sample by myself. I love being able to blast AJR in the background while ID’ing each zooplankton even though my team loves to tease me for it (but really, I’m totally putting them on). As I’ve gotten better at ID’ing zooplankton, I started brainstorming about what could help teach other interns in the future. Allison and Lisa, the previous TOPAZ/JASPER leaders, created very useful guides used to train me but I felt that there could be other interactive methods to help interns learn about zoop. Having used Quizlet in the past, I thought it would be a great resource to introduce the zooplankton basics to new interns, so I created an online Zooplankton Identification quiz!

Fig 3A & 3B: Me processing a giant sample of Atylus Tridens.
Despite having only completed three weeks of our data collection season so far, I have already learned so much! From waking up at 5:30 ten days in a row, to kayaking for four hours straight, to even counting 995 (not 1000!) zooplankton in one sitting, this internship has been amazing. It’s been a great introduction to working in the scientific field as many of the responsibilities we have been taught are completely new to me. I am excited to share this internship experience as I apply to colleges and add to my list of skills “Zoop expert.”

Fig 4: My favorite zooplankton! A Dungeness Crab Larvae.