By Mattea Holt Colberg, GEMM Lab summer intern, OSU junior
Science is about asking new questions in order to make new discoveries. Starting every investigation with a question, sparked by an observation, is enshrined in the scientific method and pursued by researchers everywhere. Asking questions goes beyond scientific research though; it is the best way to learn new things in any setting.
When I first arrived in Port Orford, I did not know much about gray whales. The extent of my knowledge was that they are large baleen whales that migrate every year and feed on plankton. I did, however, know quite a bit about killer whales. I have been interested in killer whales since I was 5 years old, so I have spent years reading about, watching, and listening to them (my current favorite book about them is Of Orcas and Men, by David Neiwert and I highly recommend it!). I have also had opportunities to research them in the Salish Sea, both on a sailing trip and through the dual-enrollment program Ocean Research College Academy, where I explored how killer whales respond to ambient underwater noise for a small independent project. Knowing more about killer whales than other species has caused killer whales to be the lens through which I approach learning and asking questions about other whales.
Figure 1a. Killer whales traveling in a group in the Salish Sea. Source: ISTOCK.
Figure 1b. A gray whale traveling solo in Tichenor Cove. Source: GEMM Lab.
At first, I was not sure how to apply what I know about killer whales specifically to research on gray whales, since killer whales are toothed whales, while gray whales are baleen whales. There are several differences between toothed whales and baleen whales; toothed whales tend to be more social, occurring in pods or groups, eat larger prey like fish, squid, and seals, and they echolocate. In comparison, baleen whales are less social, eat mostly tiny zooplankton prey, and do not echolocate. Because of these differences, I wanted to learn more about gray whales, so I started asking Lisa questions. Killer whales only sleep with half of their brain at a time, so I asked if gray whales do the same. They do. Killer whales typically travel in stable, long-term matriarchal groups, and I recently learned that gray whales frequently travel alone (though not exclusively). This new knowledge to me led me to ask if gray whales vocalize while traveling. They typically do not. Through asking these questions, and others, I have begun to learn more about gray whales.
Figure 2. Mattea on the tandem research kayak taking a break in between prey sampling. Source: L. Hildebrand.
I am still learning about marine mammal research, and from what I have experienced so far, marine mammal acoustics intrigues me the most. As a child, I developed a general interest in whale vocalizations after hearing recordings of them in museums and aquariums. Then, two years ago, I heard orcas vocalizing in the wild, and I decided I wanted to learn more about their vocalizations as a long-term career goal.
To pursue a career studying marine mammal acoustics, I will need scientific and communication skills that this internship is helping me develop. Sitting on the cliff for hours at a time, sometimes with gray whales swimming in our view-scape and sometimes without, is teaching me the patience and attention needed to review hours of sound recordings with or without vocalizations. Identifying and counting zooplankton most days is teaching me the importance of processing data regularly, so it does not build up or get too confusing, as well as attention to detail and keeping focused. Collecting data from a kayak is teaching me how to assess ocean conditions, keep track of gear, and stay calm when things go wrong. I am also practicing the skill of taking and identifying whale photos, which can be applied to many whale research topics I hope to pursue. Through writing this blog post and discussing the project with Lisa and my fellow interns, I am improving my science communication skills.
Figure 3. Mattea manning the theodolite watching and waiting for a gray whale to show up in our study area. Source: L. Hildebrand.
As an undergraduate student, it can sometimes be difficult to find opportunities to research marine mammals, so I am very grateful for and excited about this internship, both because of the skills it is helping me build and the field work experiences that I enjoy participating in. Another aspect of research this internship is helping me learn about is to ask engaging questions. As I mentioned at the beginning of this post, asking questions is a key element of conducting research. By asking questions about gray whales based on both prior knowledge and new observations, I am practicing this skill, as well as thinking of topics I am curious about and might want to explore in the future. While watching for whales, I have thought of questions such as: How is whale behavior affected by surface conditions? Do gray whales prefer feeding at certain times of the day? Questions like these help me learn about whales, and they keep me excited about research. Thanks to this internship, I can continue working towards my dreams of pursuing similar questions about whales as a career.
Yodel-Ay-Ee-Ooooo! Hello from the Theyodelers, this year’s Port Orford gray whale foraging ecology field team. In case you were wondering, no, we aren’t hobby yodelers and we don’t plan on becoming them. The team name this year actually has to be attributed to a parent of one of my interns. Shout out to Scott Holt who during the first week of the field season asked his daughter Mattea (our OSU undergraduate intern) whether using a theodolite (the instrument we use to track gray whales from our cliff site) is anything like yodeling. The name was an immediate hit with the team and so the team name discussion was closed fairly early on in the season. Now that I have explained our slightly unconventional team name, let me tell you a little about this year’s team and what has been going on down here on the Oregon south coast so far.
As you can tell from the byline, I (Lisa) am back as the project’s team lead in this, the 6th year of the Port Orford gray whale research and internship project. Going into this year’s field season with two years of experience under my belt has made me feel more confident and comfortable with diving straight back into our fine-scale research with a new team of interns. Yet, I am beginning to realize that no matter how much experience I have, there will always be unforeseeable curve balls thrown at me that I can’t anticipate no matter how prepared or experienced I am. However, my knowledge and experience now certainly inform how I tackle these curve balls and hopefully allow my problem-solving to be better and quicker. I am so thrilled that Leigh and I were able to get the field season approved here in Port Orford despite the ongoing pandemic. There were many steps we had to take and protocols to write and get approved, but it was worth the work. It certainly is strange living in a place that is meant to be your home for six weeks but having to wear a face covering everywhere except your own bedroom. However, mask wearing, frequent hand washing, and disinfecting is a very small price to pay to avoid having a lapse in our gray whale data collected here in Port Orford (and minimize transmission). Doing field research amidst COVID has certainly been a big curve ball this year but, so far, I have been able to handle these added challenges pretty well, especially with a lot of help from my team. Speaking of which, time to introduce the other Theyodelers…
Figure 1. Noah watching and waiting for whales on the cliff. When we are outside in the wind and are able to maintain a minimum 6-ft distance, we are able to remove our face coverings. Source: T. McCambridge.
First up, we have Noah Dolinajec. Noah is a fellow graduate student who is currently doing a Master’s in Marine & Lacustrine Science and Management at the Vrije Universiteit Brussel in Brussels, Belgium. While he is attending graduate school in Belgium, Noah is not actually from this European country. In fact, he is a Portlandian! As an Oregonian with a passion for the marine environment, Noah is no stranger to the Oregon coast and has spent quite some time exploring it in the past. Some other things about Noah: before going to college he played semi-professional ice hockey, he is a bit of a birder, and he likes to cook (he and I have been tag-teaming the team cooking this year).
Figure 2. Mattea outside the field station holding local fisher-pup Jim.Source: L. Hildebrand.
Next, we have Mattea Holt Colberg. As I mentioned before, Mattea is the team’s OSU undergraduate intern this year. By participating in a running-start program at her high school where she took two years of college classes, Mattea entered OSU as a junior at just 18 years old! However, she has decided to somewhat extend her undergraduate career at OSU by completing a dual major in Biology and Music. She plays the piano and the violin (which she brought to Port Orford, but we have yet to be serenaded by her). Mattea has previously conducted field research on killer whales in the Salish Sea and I can tell that she is hoping for killer whales to show up in Port Orford (while not entirely ludicrous, the chance of this happening is probably very, very slim).
Figure 3. Liz in the bow of the kayak in Tichenor Cove.Source: L. Hildebrand.
Last but certainly not least, is Liz Kelly, our Pacific High School intern from Port Orford. Liz has lived in several different states across the country (I’m talking Kentucky to Florida) and so I am really excited that she currently lives here in Oregon because she has been an absolute joy to have on the team so far. Liz brings a lot of energy and humor to the team, which we have certainly needed whenever those curve balls come flying. Besides her positivity, Liz brings a lot of determination and perseverance and seeing her work through tough situations here already has made me very proud. I really hope this internship provides Liz with the life, STEM, and communication skills she needs to help her succeed in pursuing her goals of doing wildlife research after college. As you may have read in my last blog, our previous high school interns have had successes in being admitted to various colleges to follow their goals, and I feel confident that Liz will be no different. When she is not here at the field station, she can probably be found taking care of and riding one of her four horses (Millie, Maricja, Miera, and Jeanie).
Now that I have introduced the 2020 field team, here is a short play-by-play of what we have been seeing, or perhaps more aptly, not seeing. Our whale sighting numbers have been pretty low so far and when we do see them, they seem to be foraging a little further away from our study site than I am used to seeing in past years. However, this shift in behavior is not entirely surprising to me since our zooplankton net has been coming up pretty empty at our sampling stations. While there are mysids and amphipods scattered here and there, their numbers are in the low 10s when we do our zooplankton ID lab work in the afternoons. These low counts are also reflected by the low densities I am anecdotally seeing on our GoPro drops (Fig 4).
Figure 4. Comparison of zooplankton density from our GoPro videos. Both images were taken at the same sampling station (Tichenor Cove 8), however the image on the left that contains a lot of little critters is from 2018, whereas the image on the right is from last week. This year our drops have been looking more like the image on the right, though typically with even fewer zooplankton.
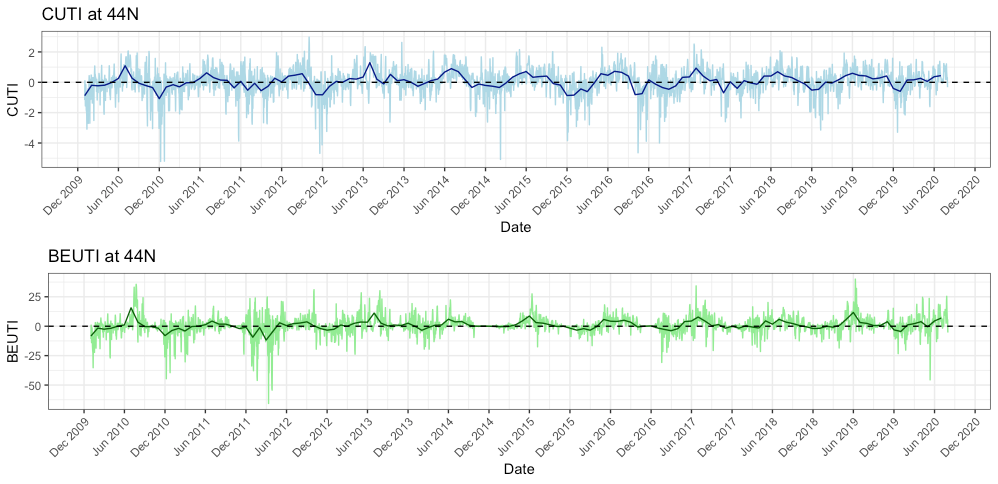
While I am not entirely certain why we are seeing this low prey abundance, I do have some hypotheses. The most likely reason is that this year we experienced some delayed upwelling on our coast. Dawn wrote a great blog about upwelling and wind a few weeks ago and I suggest checking it out to better understand what upwelling is and how it can affect whales (and the whole ecosystem). Typically, we see our peak upwelling occur here in Oregon in May-June. However, if you look at Figure 5 you will see that both the indices remained low at that time this year, whereas in previous years, they were already increasing by May/June.
Figure 5. 10 year time series of the Coastal Upwelling Transport Index (CUTI; top plot) and Biologically Effective Upwelling Transport Index (BEUTI; bottom plot) at 44ºN. CUTI represents the amount of upwelling (positive numbers) or downwelling (negative numbers) while BEUTI estimates the amount of nitrate (i.e. nutrients) upwelled (positive numbers) and downwelled (negative numbers). The light-colored lines representthe CUTI and BEUTI at that point in time while the dark, bold lines represent the long-term average.
A delayed upwelling means that there was likely less nutrients in the water to support little critters like zooplankton to start reproducing and increasing their abundances. Simply put, it means our coastal waters appear to be less productive than they usually are at this time of the year. If there is not much prey around (as we have been finding in our two study sites – Mill Rocks and Tichenor Cove), then it makes sense to me why gray whales are not hanging around since there is not much to feed on. Fortunately, the tail of the trend line in Figure 5 is angling upward, which means that the upwelling finally started in June so hopefully the nutrients, zooplankton and whales will follow soon too. In fact, since I wrote the draft of this blog at the end of last week, we have actually seen an increase in the numbers of mysids in our zooplankton net and on our GoPro videos.
We are almost halfway done with the field season already and I cannot believe how quickly it goes by! During the first two weeks we were busy getting familiar with all of our gear and completing First Aid/CPR and kayak paddle & rescue courses. This week the team started the real data collection. We have had some hiccups (we lost our GoPro stick and our backup GoPro stick, but thankfully have already recovered one of them) but overall, we are off to a pretty good start. Now we just need the upwelling to really kick in, for there to be thick layers of mysids, and for the whales to come in close. Over the next three weeks, you will be hearing from Noah, Mattea and Liz as they share their experiences and viewpoints with all of you!
By: Alexa Kownacki, Ph.D. Student, OSU Department of Fisheries and Wildlife, Geospatial Ecology of Marine Megafauna Lab
Humans are fascinated by food. We want to know its source, its nutrient content, when it was harvested and by whom, and so much more. Since childhood, I was the nagging child who interrogated wait staff about the seafood menu because I cared about the sustainability aspect as well as consuming ethically-sourced seafood. Decades later I still do the same: ask a myriad of questions from restaurants and stores in order to eat as sustainably as possible. But in addition to asking these questions about my food, I also question what my study species eats and why. My study populations, common bottlenose dolphins, are described as top opportunistic predators (Norris and Prescott 1961, Shane et al. 1986, Barros and Odell 1990). In my study area off of California, this species exists in two ecotypes. The coastal ecotype off of California, USA are generalist predators, feeding on many different species of fish using different foraging techniques (Ballance 1992, Shane 1990). The offshore ecotype, on the other hand, is less well-studied, but is frequently observed in association with sperm whales, although the reason is still unknown (Díaz-Gamboa et al. 2018). Stable isotope analysis from skin samples from the two ecotypes indicates that the ecotypes exhibit different foraging strategies based on different isotopic carbon and nitrogen levels (Díaz-Gamboa et al. 2018).
Growing up, I kept the Monterey Bay Aquarium’s Seafood Watch Guide with me to choose the most sustainably-sourced seafood at restaurants. Today there is an easy-to-use application for mobile phones that replaced the paper guide. (Image Source: https://www.seafoodwatch.org/)
Preliminary and historical data on common bottlenose dolphins (Tursiops truncatus) suggest that the coastal ecotype spend more time near estuary mouths than offshore dolphins (Ballance 1992, Kownacki et al. unpublished data). Estuaries contain large concentrations of nutrients from runoff, which support zooplankton and fishes. It is for this reason that these estuaries are thought to be hotspots for bottlenose dolphin foraging. Some scientists hypothesize that these dolphins are estuarine-based prey specialists (Barros and Odell 1990), or that the dolphins simply aggregate in estuaries due to higher prey abundance (Ballance 1992).
Coastal bottlenose dolphins traveling near an estuary mouth in San Diego, CA. (Photographed under NOAA NMFS Permit # 19091).
In an effort to understand diet compositions of bottlenose dolphins, during coastal surveys seabirds were recorded in association with feeding groups of dolphins. Therefore, it is reasonable to believe that dolphins were feeding on the same fishes as Brown pelicans, blue-footed and brown boobies, double-crested cormorants, and magnificent frigatebirds, seeing as they were the most common species associated with bottlenose dolphin feeding groups (Ballance 1992). A shore-based study by Hanson and Defran (1993) found that coastal dolphins fed more often in the early morning and late afternoon, as well as during periods of high tide current. These patterns may have to do with the temporal and spatial distribution of prey fish species. From the few diet studies conducted on these bottlenose dolphins in this area, 75% of the prey were species from the families Ebiotocidae (surf perches) and Sciaendae (croakers) (Norris and Prescott 1961, Walker 1981). These studies, in addition to optimal foraging models, suggest this coastal ecotype may not be as much of a generalist as originally suggested (Defran et al. 1999).
A redtail surfperch caught by a fisherman from a beach in San Diego, CA. These fish are thought to be common prey of coastal bottlenose dolphins. (Image Source: FishwithJD)
Diet studies on the offshore ecotype of bottlenose dolphins worldwide show a preference for cephalopods, similar to other toothed cetaceans who occupy similar regions, such as Risso’s dolphin, sperm whales, and pilot whales (Clarke 1986, Cockcroft and Ross 1990, Gonzalez et al. 1994, Barros et al. 2000, Walker et al. 1999). Because these animals seldom strand on accessible beaches, stomach contents analyses are limited to few studies and isotope analysis is more widely available from biopsies. We know these dolphins are sighted in deeper waters than the habitat of coastal dolphins where there are fewer nutrient plumes, so it is reasonable to hypothesize that the offshore ecotype consumes different species and may be more specialized than the coastal ecotype.
An bottlenose dolphin forages on an octopus. (Image source: Mandurah Cruises)
For a species that is so often observed from shore and boats, and is known for its charisma, it may be surprising that the diets of both the coastal and offshore bottlenose dolphins are still largely unknown. Such is the challenge of studying animals that live and feed underwater. I wish I could simply ask a dolphin, much like I would ask staff at restaurants: what is on the menu today? But, unfortunately, that is not possible. Instead, we must make educated hypotheses about the diets of both ecotypes based on necropsies and stable isotope studies, and behavioral and spatial surveys. And, I will continue to look to new technologies and creative thinking to provide the answers we are seeking.
Literature cited:
Ballance, L. T. (1992). Habitat use patterns and ranges of the bottlenose dolphin in the Gulf of California, Mexico. Marine Mammal Science, 8(3), 262-274.
Barros, N.B., and D. K. Odell. (1990). Food habits of bottlenose dolphins in the southeastern United States. Pages 309-328 in S. Leatherwood and R. R. Reeves, eds. The bottlenose dolphin. Academic Press, San Diego, CA.
Barros, N., E. Parsons and T. Jefferson. (2000). Prey of bottlenose dolphins from the South China Sea. Aquatic Mammals 26:2–6.
Clarke, M. 1986. Cephalopods in the diet of odontocetes. Pages 281–321 in M. Bryden and R. Harrison, eds. Research on dolphins. Clarendon Press, Oxford, NY.
Cockcroft, V., and G. Ross. (1990). Food and feeding of the Indian Ocean bottlenose dolphin off southern Natal, South Africa. Pages 295–308 in S. Leatherwood and R. R. Reeves, eds. The bottlenose dolphin. Academic Press, San Diego, CA.
Defran, R. H., Weller, D. W., Kelly, D. L., & Espinosa, M. A. (1999). Range characteristics of Pacific coast bottlenose dolphins (Tursiops truncatus) in the Southern California Bight. Marine Mammal Science, 15(2), 381-393.
Díaz‐Gamboa, R. E., Gendron, D., & Busquets‐Vass, G. (2018). Isotopic niche width differentiation between common bottlenose dolphin ecotypes and sperm whales in the Gulf of California. Marine Mammal Science, 34(2), 440-457.
Gonzalez, A., A. Lopez, A. Guerra and A. Barreiro. (1994). Diets of marine mammals stranded on the northwestern Spanish Atlantic coast with special reference to Cephalopoda. Fisheries Research 21:179–191.
Hanson, M. T., and Defran, R. H. (1993). The behavior and feeding ecology of the Pacific coast bottlenose dolphin, Tursiops truncatus. Aquatic Mammals, 19, 127-127.
Norris, K. S., and J. H. Prescott. (1961). Observations on Pacific cetaceans of Californian and Mexican waters. University of California Publications of Zoology 63:29, 1-402.
Shane, S. H. (1990). Comparison of bottlenose dolphin behavior in Texas and Florida, with a critique of methods for studying dolphin behavior. Pages 541-558 in S. Leatherwood and R. R. Reeves, eds. The bottlenose dolphin. Academic Press, San Diego, CA.
Shane, S., R. Wells and B. Wursig. (1986). Ecology, behavior and social organization of bottlenose dolphin: A review. Marine Mammal Science 2:34–63.
Walker, W.A. (1981). Geographical variation in morphology and biology of the bottlenose dolphins (Tursiops) in the eastern North Pacific. NMFS/SWFC Administrative Report. No, LJ-91-03C.
Walker, J., C. Potter and S. Macko. (1999). The diets of modern and historic bottlenose dolphin populations reflected through stable isotopes. Marine Mammal Science 15:335–350.
What do I mean by impact? There are different ways to measure the impact of science and I bet that the readers of this blog had different ideas pop into their heads when they read the title. My guess is that most ideas were related to the impact factor (IF) of a journal, which acts as a measure of a journal’s impact within its discipline and allows journals to be compared. Recent GEMM Lab graduate and newly minted Dr. Leila Lemos wrote a blog about this topic and I suggest reading it for more detail. In a nutshell though, the higher the IF, the more prestigious and impactful the journal. It is unsurprising that scientists found a way to measure our impact on the broader scientific community quantitatively.
However, IFs are not the impact I was referring to in my title. The impact I am talking about is arguably much harder to measure because you can’t easily put a number on it. I am talking about the impact we have on communities and individuals through outreach and engagement. The GEMM Lab’s Port Orford gray whale ecology project, which I lead, is going into its 6th consecutive year of summer field work this year. Outreach and engagement are two core components of the project that I have become very invested in since I started in 2018. And so, since we are only one week away from the field season commencing (yes, somehow it’s mid-July already…), for this week’s blog I have decided to reflect on what scientific outreach and engagement is, how we have tried to do both in Port Orford, and some of the associated highs and lows.
2018 team member Dylan presenting at the Port Orford community presentation. Source: T. Calvanese.
I think almost everyone in the scientific community would agree that outreach and engagement are important and that we should strive to interact frequently with the public to be transparent and build public trust, as well as to enable mutual learning. However, in my opinion, most scientists rarely put in the work needed to actually reach out to, and engage with, the community. Outreach and engagement have become buzzwords that are often thrown around, and with some hand-waving, can create the illusion that scientists are doing solid outreach and engagement work. For some, the words are probably even used interchangeably, which isn’t correct as they mean two different things.
Outreach and engagement should be thought of as occurring on two different ends of a spectrum. Outreach occurs in a one-way direction. Examples of outreach are public seminars delivered by a scientist (like Hatfield’s monthly Science on Tap) or fairs where the public is invited to come and talk to different scientific entities at their respective booths (like Hatfield’s annual Marine Science Day). Outreach is a way for scientists to disseminate their research to the public and often do not warrant the umbrella term engagement, as these “conversations” are not two-way. Engagement is collaborative and refers to intentional interactions where both sides (public and scientist) share and receive. It goes beyond a scientist telling the public about what they have been doing, but also requires the scientist to listen, absorb, and implement what the views from the ‘other side’ are.
2015 team tracking a whale on Graveyard Point above the port of Port Orford. Source: F. Sullivan.
Now that I have (hopefully) clarified the distinction between the two terms, I am going to shift the focus to specifically talk about the Port Orford project. Before I do, I would like to emphasize that I do not think our outreach and engagement is the be-all and end-all. There is definitely room for improvement and growth, but I do believe that we actively work hard to do both and to center these aspects within the project, rather than doing it as an afterthought to tick a box.
In talking about outreach and engagement, I have been using the words ‘public’ and ‘community’. I think these words conjure an image of a big group of people, an entire town, county, state or even nation. While this can be the case, it can also refer to smaller groups of people, even individuals. The outreach we conduct for the Port Orford project certainly occurs at the town-level. At the end of every field season, we give a community presentation where the field team and Leigh present new findings and give a recount of the field season. In the past, various teams have also given talks at the Humbug Mountain Campground and at Redfish Rocks Community Team events. These events, especially the community presentation, have been packed to the brim every year, which shows the community’s interest for the gray whales and our research. In fact, Tom Calvanese, the OSU Port Orford Field Station manager, has shared with me that now in early summer, Port Orford residents ask him when the ‘whale team’ is returning. I believe that our project has perhaps shifted the perception the local community has of scientists a little bit. Although in our first year or two of the project we may have been viewed as nosy outsiders, I feel that now we are almost honorary members within the community.
A packed room at the 2017 Port Orford community presentation. Photo: GEMM Lab.
Our outreach is not just isolated to one or two public talks per field season though. We have been close collaborators with South Coast Tours (SCT), an adventure tour company headed by Dave Lacey, since the start of the project. During the summer, SCT has almost daily kayak and fishing tours (this year, boat tours too!) out of Port Orford. The paddle routes of SCT and our kayak team will typically intersect in Tichenor’s Cove around mid-morning. When this happens, we form a little kayak fleet with the tour and research kayaks and our kayak team gives a short, informal talk about our research. We often pass around samples of zooplankton we just collected and answer questions that many of the paddlers have. These casual interactions are a highlight to the guests on SCT’s tours (Dave’s words, not mine) and they also provide an opportunity for the project’s interns to practice their science communication skills in a ‘low-stakes’ setting.
The nature of our engagement is more at the individual-level. Since the project’s conception in 2015, the team has been composed of some combination of 4-5 students, be it high school, undergraduate or graduate students. Aside from Florence Sullivan and myself as the GEMM Lab graduate student project leads, in total, we have had 16 students participate in the program, of which 4 were high school students (two from Port Orford’s Pacific High School and two from Astoria High School), 11 OSU and Lawrence University undergraduates, and 1 Duke University graduate student. This year we will be adding 3 more to the total tally (1 Pacific High School student, 1 OSU undergrad, and 1 graduate student from the Vrije Universiteit Brussel in Belgium). I am the first to admit that our yearly (and total) numbers of ‘impacted’ students is small. Limitations of funding and also general logistics of coordinating a large group of interns to participate in field work prevent us from having a larger cohort participate in the field season every summer. However, the impact on each of these students is huge.
The 2019 team with Dave Lacey who instructed our kayak paddle & safety course. Photo: L. Hildebrand.
If I had to pick one word to describe the 6-week Port Orford field season, it would be ‘intense’. The word is perfect because it can simultaneously describe something positive and negative, and the Port Orford field season definitely has elements of both. Both as a team and as individuals we experience incredible high points (an example being last year when we saw Port Orford’s favorite whale ‘Buttons’ breach multiple times on several different days), but we also have pretty low points (I’m thinking of a day in 2018 when two of my interns tried incredibly hard to get our GoPro stick dislodged from a rocky crevice for over 1-hour before radioing me to tell me they couldn’t retrieve it). These highs and lows occur on top of the team’s slowly depleting levels of energy as the field season goes on; with every day we get up at 5:30 am and we get a little more exhausted. The work requires a lot of brain power, a lot of muscle, and a lot of teamwork. Like I said, it’s intense and that’s coming from someone who had several years of marine mammal field work experience before running this project for the first time in 2018. The majority of the interns who have participated in our project have had no marine mammal field experience, some have had no field experience at all. It’s double, if not triple, intense for the interns!
I ask a lot of my interns. I am aware of that. It has been a steep learning curve for me since I took on the project in 2018. I’ve had to adjust my expectations and remember not to measure the performance of my interns against my own. I can always give 110% during the field season, even when I’m exhausted, because the stakes are high for me. After all, the data that is being collected feeds straight into my thesis. However, it took me a while to realize that the stakes, and therefore the motivation, aren’t the same for my interns as they are for me. And so, expecting them to perform at the same level I am, is unfair. I believe I have grown a lot since running that first field season. I have taken the feedback from interns to heart and tried to make adjustments accordingly. While those adjustments were hard because it ultimately meant making compromises that affected the amount of data collected, I recognize and respect the need to make those adjustments. I am incredibly grateful to all of the interns, including the ones that participated before my leadership of the project, who really gave it their all to collect the data that I now get to dig into and draw conclusions from.
2016 interns Kelli and Catherine paddling to a kayak sampling station. Photo: F. Sullivan.
But, as I said before, engagement is not one-sided, and I am not the only one who benefits from having interns participate in the project. The interns themselves learn a wealth of skills that are valuable for the future. Some of these skills are very STEM (Science, Technology, Engineering & Mathematics) specific (e.g. identifying zooplankton with a microscope, tracking whales with a theodolite), but a lot of them are transferrable to non-STEM futures (e.g. attention to detail and concentration required for identifying zooplankton, team work, effective communication). Our reach may be small with this project but the impact that participating in our internship has on each intern is a big one. Three of our four high school interns have gone on to start college. One plans to major in Marine Studies (in part a result of participating in this internship) while another decided to go to college to study Biology because of this internship. Several of the undergraduate students that participated in the 2015, 2016, 2017 & 2018 field seasons have gone on to start Master’s degrees at graduate schools around the country (3 of which have already graduated from their programs). A 2015 intern now teaches middle school in Washington and a 2016 intern is working with Oceans Initiative on their southern resident killer whale project this summer. Leigh, Florence and I have written many letters of recommendations for our interns, and these letters were not written out of duty, but out of conviction.
I love working closely with students and watching them grow. For the last two years, my proudest moment has always been watching my interns present our research at the annual community presentation we give at the end of the field season in Port Orford. No matter the amount of lows and struggles I experienced throughout the season, I watch my interns and my face almost hurts because of the huge smile on my face. The interns truly undergo a transformation where at the start of the season they are shy or feel inadequate and awkward when talking to the public about gray whales and the methods we employ to study them. But on that final day, there is so much confidence and eloquence with which the interns talk about their internship, that they are oftentimes even comfortable enough to crack jokes and share personal stories with the audience. As I said before, engagement of this nature is hard to measure and put a number on. Our statistic (engaging with 16 students) makes it sound like a small impact, but when you dig into what these engagements have meant for each student, the impact is enormous.
All of the past PO gray whale ecology teams, from left to right: 2015 (Sarah, Florence, Cricket, Justin), 2016 (Florence, Kelli, Catherine, Cathryn), 2017 (Nathan, Quince, Florence, Morgan), 2018 (Haley, Robyn, Hayleigh, Dylan, Lisa), and 2019 (Anthony, Donovan, Lisa, Mia). Bottom left: Florence and Leigh; bottom right: Lisa and Leigh.
I treasure my 6 weeks in Port Orford. Even though they are intense and there are new challenges every year, they bring me a lot of happiness. And it’s only in part because I get to see gray whales and kayak on an (almost) daily basis. A large part is because of the bonds I have formed and continue to cultivate with Port Orford locals, the leaps and bounds I know the interns will make, and the fact that the gray whales, completely unknowingly, bring together a small group of students and a community every year.
If you feel like taking a trip down memory lane, below are the links of the blogs written by previous PO interns:
Knowing what and how much prey a predator feeds on are key components to better understanding and conserving that predator. Prey abundance and availability are frequently predictors for marine predator reproductive success and population dynamics. It is the reason why the GEMM Lab makes a concerted effort to not only track our main taxa of interest (marine mammals) but to simultaneously measure their prey. However, over the last decade or two, there has been increased recognition that prey quality is also highly important in understanding a predator’s ecology (Spitz et al. 2012). Optimal foraging theory is a widely accepted framework that posits that predators should attempt to maximize energy gained and minimize energy spent during a foraging event (Charnov 1976, Krebs 1978, Pyke 1984). Thus, knowledge of how valuable a prey item is in terms of its energetic content is an important part of the equation when applying optimal foraging theory to a predator of interest.
Ideally, the prey species with the highest energetic value would also be the easiest, most ubiquitous and least energetically expensive prey item to capture and consume, such that a predator truly could expend very little energy to get very high energetic rewards. However, it rarely is this straightforward. The caloric content of several marine prey species has been shown to increase with increasing size (e.g. Benoit-Bird 2004; Fig. 1), both length and weight. Yet, increasing size often also means increased mobility and, as a result, ability to evade and escape predation. Furthermore, increasing size also inherently means decreasing abundances – there will always be billions more krill in the ocean than whales based solely on cost of reproduction. Therefore, just based on sheer numbers, there are fewer big prey items, which increases the time between, and decreases the likelihood of, a predator encountering big prey items. So, there are clear trade-offs here. It may take longer to locate and capture a high value prey item, which costs more energy to capture, but the payout could potentially be much bigger. However, if a predator gambles too much, then their net energy expenditure to obtain high value prey may be higher than the net energy gained. Instead, it may be worth pursuing smaller prey items with lower energetic values, where discovery and capture success are higher and more frequent. However, in this case, many, many more pursuits are likely needed, thus costing more energy to meet daily energetic demands.
Figure 1. Increasing caloric content with increasing length (a) and wet weight (b). Figures and caption reproduced from Benoit-Bird 2004.
Is your head spinning as much as mine? Let me try and simplify this complex web of interactions with a tangible example. Bowen et al. (2002) investigated foraging of harbor seals in Nova Scotia to assess prey profitability of different species. By attaching camera systems to the backs of 39 adult male harbor seals, the authors identified sand lance and flounder to be the most targeted prey species. However, there were significant differences in pursuit/handling cost per prey type (kJ/min) with sand lance only requiring 14.8 ± 2.7, whereas flounder required significantly more at 30.3 ± 7.9. Therefore, based solely on energy required to capture prey, the sand lance would seem to be the better option. In fact, to a certain degree, this hypothesis is actually true when we compare the energetic content of the two prey types. Sand lance have a higher energetic value at lengths of 10 and 15 cm (53.6 and 95.8 kJ, respectively) compared to flounder (22.6 and 88.6 kJ, respectively). So, the net gain of a harbor seal foraging on a 15 cm sand lance (assuming that it only takes 1 minute to catch the fish – this is more for explanatory purposes as it likely takes much longer for a harbor seal to capture a fish) would be 81 kJ. This gain is larger than that of a 15 cm flounder (58.3 kJ). However, once we compare these fish at 20 and 25 cm lengths, the flounder actually becomes the more beneficial prey item at 232.6 and 492.3 kJ, respectively, over the sand lance (158.1 and 233.8 kJ). Now, assuming once again that it only takes 1 minute to catch the fish, the harbor seal enjoys a net energetic gain of a whopping 462 kJ when capturing a 25 cm flounder compared to 219 kJ for a sand lance of the same size – that makes the flounder more than twice as profitable!
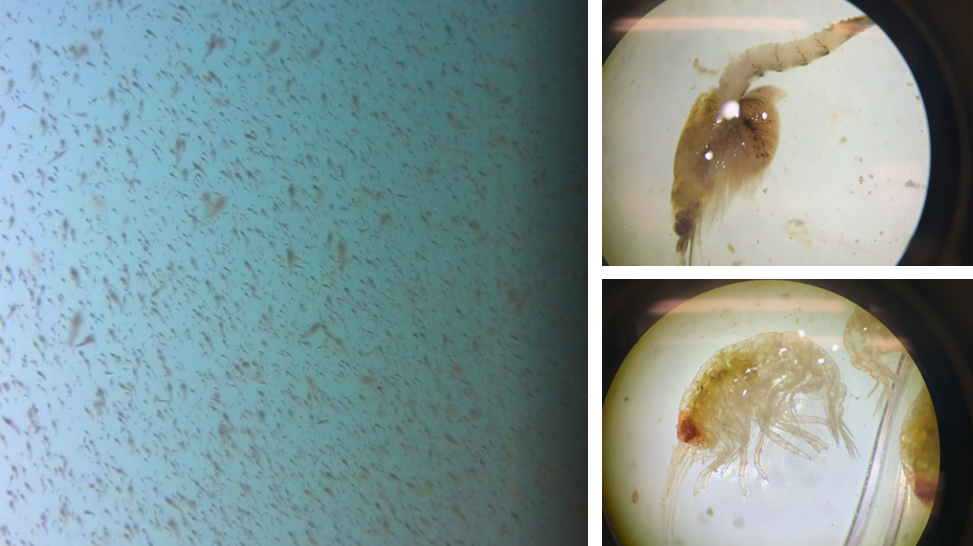
The Bowen et al. study is an excellent demonstration of the importance of considering the quality of prey items when studying the ecology of marine predators. However, the authors did not assess the relative availability of sand lance and flounder. Ideally, foraging ecology studies aimed at understanding prey choice would try to address both important prey metrics – quality and quantity. This goal is the exact aim of my second Master’s thesis chapter where I am investigating whether prey quality (determined through community composition and caloric content) or prey quantity (measured as relative density) is more important in driving fine-scale gray whale foraging behavior in Port Orford, Oregon (Fig. 2). This question can be simplified by asking does it matter more what prey is in an area, or how much prey there is in an area? Or we can relate it back to the title of this post by asking whether individual gray whales would rather attend a cheap all-you-can-eat buffet or an expensive fine-dining restaurant. I am unfortunately not quite done with my analyses yet (but I’m getting closer!) and therefore am not ready to answer these questions. However, I have done extensive research on this topic and therefore am in a position to briefly mention a few other studies that have investigated these questions for other marine predators.
Figure 2. A question of what or how much. Left image: example of the screenshots we take to estimate relative prey density in Port Orford. Right images: two examples of the main prey species we find (top: mysid shrimp Neomysis rayii with a full brood pouch; bottom: amphipod Polycheria osborni).
Ludynia et al. (2010) explored reasons why African penguin (Spehniscus demersus) numbers have declined in Namibia. They found that after the collapse of pelagic fish stocks in the 1970s (including the principal penguin prey item, sardine), African penguins switched to feeding on bearded goby, which are considered a low-energy prey species. Bearded goby are relatively abundant along Namibia’s southern coast and as such, limited prey availability is not the reason for declining African penguin numbers. Therefore, the authors concluded that the low quality of bearded goby (compared to sardine) appears to be the reason for declining population trends of the penguins. This study demonstrates that African penguins do better when eating at a fine-dining restaurant, rather than loading up a whole plate of junk food.
Grémillet et al. (2004) studied the foraging effort and number of successful prey captures per foraging trip (yield) of great cormorants (Phalacrocorax carbo) in Greenland in relation to prey abundance and quality within their foraging areas. The authors radio-tracked 11 great cormorants during a total of 163 foraging trips to estimate foraging effort and yield. The study found that contrary to the authors’ hypothesis, great cormorants foraged in areas of low prey abundance where the average caloric value was also relatively low. Therefore, in this example, it would seem that the predator of interest prioritizes neither high quality nor quantity when foraging.
Haug et al. (2002) investigated the variations in minke whale (Balaenoptera acutorostrata) diet and body condition in response to ecosystem changes in the Barents Sea. The main prey item of minke whales in the Barents Sea is immature herring. However, when recruitment failure and subsequent weak cohorts leads to reduced availability of immature herring, minke whales switched their diet to other prey items such as krill, capelin, and sometimes other gadoid fish species. The authors found a correlation between body condition of minke whales and immature herring abundances, such that minke whales displayed a poor body condition during low immature herring abundances. However, in the years of low immature herring abundance, abundances of krill and capelin were not low. Therefore, similar to the Ludynia et al. (2010) study, it seems that minke whales in the Barents Sea also do better in years when the prey type of highest caloric value is the most abundant. However, decreases in high quality prey has not led to population declines in minke whales in the Barents Sea, indicating that they likely take advantage of high quantities of low quality prey, unlike the African penguins.
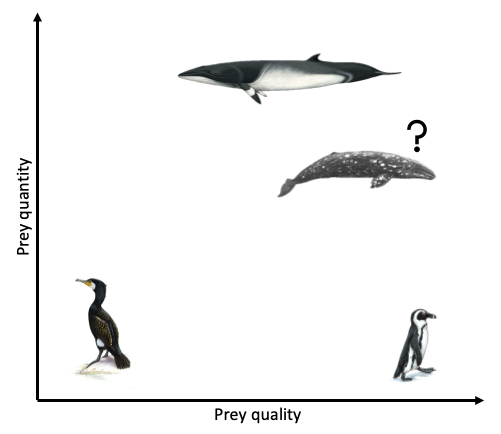
Clearly, the answer as to whether marine predators prefer quality over quantity is not simple and constant. Rather, prey preference varies based on predator needs and ecology, falling anywhere on a broad spectrum from low to high prey quality and low to high prey quantity (Fig. 3). To a certain extent, it probably also is not solely predator choice that determines what they eat but many other factors, such as climate, disturbance, and health. As a result, these preferences and choices will likely be fluid, rather than fixed. While I anticipate that individual gray whales will be flexible foragers, I do hypothesize that when there is a prey patch of a higher energetic value in the area, whales will preferentially consume these patches over areas where there is less energetically rich prey, even if it is more abundant.
Figure 3. A spectrum of prey quantity and quality. Giant cormorants forage on low prey quality & quantity (Grémillet et al. 2004). African penguin populations are declining despite high abundances of low quality prey, suggesting that high prey quality is important for their survival (Ludynia et al. 2010). Body condition of Barents Sea minke whales decreases when high quality prey is less abundant, however their populations have not declined, suggesting they instead exploit high abundances of low quality prey (Haug et al. 2002). What will the gray whales do?
Literature cited
Benoit-Bird, K. J. 2004. Prey caloric value and predator energy needs: foraging predictions for wild spinner dolphins. Marine Biology 145:435-444.
Bowen, W. D., D. Tuley, D. J. Boness, B. M. Bulheier, and G. J. Marshall. 2002. Prey-dependent foraging tactics and prey profitability in a marine mammal. Marine Ecology Progress Series 244:235-245.
Charnov, E. L. 1976. Optimal foraging, the marginal value theorem. Theoretical Population Biology 9(2):129-136.
Grémillet D., G. Kuntz, F. Delbart, M. Mellet, A. Kato, J-P. Robin, P-E. Chaillon, J-P. Gendner, S-H. Lorentsen, and Y. Le Maho. 2004. Linking the foraging performance of a marine predator to local prey abundance. Functional Ecology 18(6):793-801.
Haug, T., U. Lindstrøm, and K. T. Nilssen. 2002. Variations in minke whale (Balaenoptera acutorostrata) diet and body condition in response to ecosystem changes in the Barents Sea. Sarsia 87(6):409-422.
Krebs, J. R. 1978. Optimal foraging: decision rules for predators. Behvaioral Ecology: An Evolutionary Approach, eds. Krebs, J. R., and N. B. Davies. Oxford: Blackwell.
Ludynia, J., J-P. Roux, R. Jones, J. Kemper, and L. G. Underhill. 2010. Surviving off junk: low-energy prey dominates the diet of African penguins Spheniscus demersus at Mercury Island, Namibia, between 1996 and 2009. African Journal of Marine Science 32(3):563-572.
Pyke, G. H. 1984. Optimal foraging theory: a critical review. Annual Reviews of Ecology and Systematics 15:523-575.
Spitz, J., A. W. Trites, V. Becquet, A. Brind’Amour, Y. Cherel, R. Galois, and V. Ridoux. 2012. Cost of living dictates what whales, dolphins and porpoises eat: the importance of prey quality on predator foraging strategies. PLoS ONE 7(11):e50096.
Young, J. K., B. A. Black, J. T. Clarke, S. V. Schonberg, and K. H. Dunton. 2017. Abundance, biomass and caloric content of Chukchi Sea bivalves and association with Pacific walrus (Odobenus rosmarus divergens) relative density and distribution in the northeastern Chukchi Sea. Deep-Sea Research Part II 144:125-141.
By: Alexa Kownacki, Ph.D. Student, OSU Department of Fisheries and Wildlife, Geospatial Ecology of Marine Megafauna Lab
While sitting on my porch and watching the bees pollinate the blooming spring flowers, I intermittently pause to scratch the hives along my shoulders and chest. In the middle of my many Zoom calls, I mute myself and stop my video because a wave of pollen hits my face and I immediately have to sneeze. With this, I’m reminded: Welcome to prime allergy season in the Northern Hemisphere. As I was scratching my chronic idiopathic urticaria (hives caused by an overactive immune system), I asked myself “Do dolphins get hives?” I had no idea. I know most terrestrial mammals can and do—just yesterday, one of the horses in the nearby pasture was suffering from a flare of hives. But, what about aquatic and marine mammals?
Springtime flowers blooming on the Central California Coast 2017. (Image Source: A. Kownacki)
As with most research on marine mammal health, knowledge is scare and is frequently limited to studies conducted on captive and stranded animals. Additionally, most of the current theories on allergic reactions in marine mammals are based on studies from terrestrial wildlife and humans. Because nearly all research on histamine pathways centers on terrestrial animals, I wanted to see what information exists the presence of skin allergies in marine mammals.
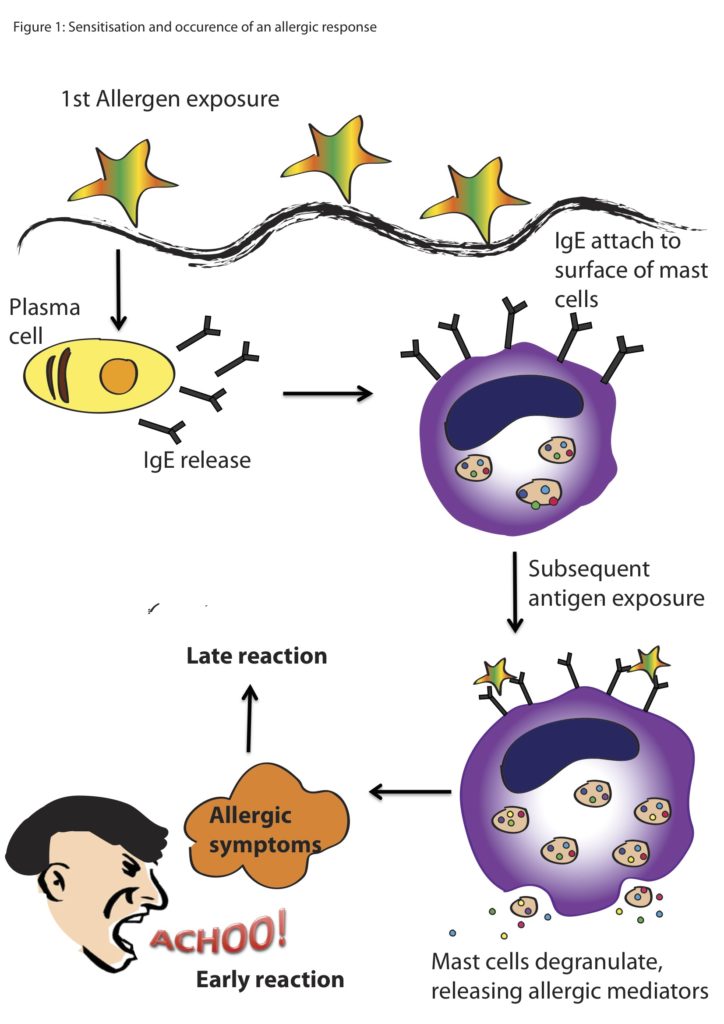
Allergic reactions trigger a cascade within the body, beginning with the introduction of a foreign body, which for many people is pollen. The allergen binds to antibodies that are produced to fight potentially harmful substances. Once this allergen binds to different types of cells, including mast cells, chemicals like histamines are released. Histamines cause the production of mucus and constriction of blood vessels, and thus are the reason your eyes water, your nose runs, or you start coughing.
Basic cartoon of an allergic reaction from exposure to the allergen to the reaction from the animal. (Image Source: Scientific Malaysian)
As you probably can tell just by looking at a marine mammal, they have thicker skin and fewer mucus membranes that humans, due to the fact that they live in the water. However, mast cells or mast cell-like cells have been described in most vertebrate lineages including mammals, birds, reptiles, amphibians, and bony fishes (Hellman et al. 2017, Reite and Evenson 2006). Mast cell-like cells have also been described in an early ancestor of the vertebrates, the tunicate, or sea squirt (Wong et al. 2014). Therefore, allergic-reaction cascades that may present as hives, red and itchy eyes or nose in humans, also exist in marine mammals, but perhaps cause different or less visible symptoms.
Skin conditions in cetaceans are gathering interest within the marine mammal health community. Even our very own Dawn Barlow, Dr. Leigh Torres, and Acacia Pepperassessed the skin conditions in New Zealand blue whales in their recent publication. Most visible skin lesions or markings on cetaceans are caused by parasites, shark bits, fungal infections, and fishery or boat interactions (Leone et al. 2019, Sweeney and Ridgway 1985). However, there is very little scientific literature about allergic reactions in marine mammals, let alone cetaceans. That being said, I managed to find a few critical pieces of information supporting the theory that marine mammals do in fact have allergies that can produce dermal reactions similar to hives in humans.
In one study, three captive bottlenose dolphins developed reddened skin, sloughing, macules, and wheals on their ventral surfaces (Monreal-Pawlowsky et al. 2017). The medical staff first noticed this atopic dermatitis in 2005 and observed the process escalate over the next decade. Small biopsy samples from the affected areas on the three dolphins coincided with the appearance of four pollens in the air within the geographic region: Betula, Pistacia, Celtis, and Fagus (Monreal-Pawlowsky et al. 2017). Topical prednisone treatments were applied to the affected areas at various dosages that slowly resolved the skin irritations. Researchers manufactured an allergy vaccine using a combination of the four pollens in hopes that it would prevent further seasonal outbreaks, but it was unsuccessful. In the coming years, the facility intends to adjust the dosages to create a successful vaccine.
In the three top images, visible skin irritation including redness, macules, wheals, and sloughing are present. In the image below, the above animal was treated with methylprednisolone and the skin irritation subsides. (Monreal-Pawlowsky et al. 2017)
In addition to the above study, there is an unpublished case of suspected allergic reaction to another pollen that produces a pruritic reaction on the ventral areas of dolphins on a seasonal basis (Vicente Arribes, personal communication). Although there are only a few documented cases of environmentally-triggered allergic reactions that are visible on the dermal layer of cetaceans, I believe this evidence makes the case that some cetaceans suffer from allergies much like us. So, next time you’re enjoying the beautiful blooms and annoyingly scratch your eyes, know that you are not alone.
Image Source: FurEver Family
Citations:
Barlow DR, Pepper AL and Torres LG (2019) Skin Deep: An Assessment of New Zealand Blue Whale Skin Condition. Front. Mar. Sci. 6:757.doi: 10.3389/fmars.2019.00757
Hellman LT, Akula S, Thorpe M and Fu Z (2017) Tracing the Origins of IgE, Mast Cells, and Allergies by Studies of Wild Animals. Front. Immunol. 8:1749. doi: 10.3389/fimmu.2017.01749
Leone AB, Bonanno Ferraro G, Boitani L, Blasi MF. Skin marks in bottlenose dolphins (Tursiops truncatus) interacting with artisanal fishery in the central Mediterranean Sea. PLoS One. 2019;14(2):e0211767. Published 2019 Feb 5. doi:10.1371/journal.pone.0211767
Monreal-Pawlowsky T, Fernández-Bellon H, Puigdemont A (2017) Suspected Allergic Reaction in Bottlenose Dolphins (Tursiops truncatus). J Vet Sci Ani Husb 5(1): 108. doi: 10.15744/2348-9790.5.108
Reite OB, Evensen O. Inflammatory cells of teleostean fish: a review focusing on mast cells/eosinophilic granule cells and rodlet cells. Fish Shellfish Immunol (2006) 20:192–208. doi:10.1016/j.fsi.2005.01.012
Sweeney, J. C., & Ridgway, S. H. (1975). Common diseases of small cetaceans. J. Am. Vet. Med. Assoc, 167(7), 533-540.
Wong GW, Zhuo L, Kimata K, Lam BK, Satoh N, Stevens RL. Ancient originof mast cells. Biochem Biophys Res Commun (2014) 451:314–8. doi:10.1016/j.bbrc.2014.07.124
The last two months have been challenging for everyone across the world. While I have also experienced lows and disappointments during this time, I always try to see the positives and to appreciate the good things every day, even if they are small. One thing that I have been extremely grateful and excited about every week is when the clock strikes 9:58 am every Thursday. At that time, I click a Zoom link and after a few seconds of waiting, I am greeted by the smiling faces of the GEMM Lab. This spring term, our Principal Investigator Dr. Leigh Torres is teaching a reading and conference class entitled ‘Cetacean Behavioral Ecology’. Every week there are 2-3 readings (a mix of book chapters and scientific papers) focused on a particular aspect of behavioral ecology in cetaceans. During the first week we took a deep dive into the foundations of behavioral ecology (much of which is terrestrial-based) and we have now transitioned into applying the theories to more cetacean-centric literature, with a different branch of behavior and ecology addressed each week.
Leigh dedicated four weeks of the class to discussing foraging behavior, which is particularly relevant (and exciting) to me since my Master’s thesis focuses on the fine-scale foraging ecology of gray whales. Trying to understand the foraging behavior of cetaceans is not an easy feat since there are so many variables that influence the decisions made by an individual on where and when to forage, and what to forage on. While we can attempt to measure these variables (e.g., prey, environment, disturbance, competition, an individual’s health), it is almost impossible to quantify all of them at the same time while also tracking the behavior of the individual of interest. Time, money, and unworkable weather conditions are the typical culprits of making such work difficult. However, on top of these barriers is the added complication of scale. We still know so little about the scales at which cetaceans operate on, or, more importantly, the scales at which the aforementioned variables have an effect on and drive the behavior of cetaceans. For instance, does it matter if a predator is 10 km away, or just when it is 1 km away? Is a whale able to sense a patch of prey 100 m away, or just 10 m away? The same questions can be asked in terms of temporal scale too.
What is that gray whale doing in the kelp? Source: F. Sullivan.
As such, cetacean field work will always involve some compromise in data collection between these factors. A project might address cetacean movements across large swaths of the ocean (e.g., the entire U.S. west coast) to locate foraging hotspots, but it would be logistically complicated to simultaneously collect data on prey distribution and abundance, disturbance and competitors across this same scale at the same time. Alternatively, a project could focus on a small, fixed area, making simultaneous measurements of multiple variables more feasible, but this means that only individuals using the study area are studied. My field work in Port Orford falls into the latter category. The project is unique in that we have high-resolution data on prey (zooplankton) and predators (gray whales), and that these datasets have high spatial and temporal overlap (collected at nearly the same time and place). However, once a whale leaves the study area, I do not know where it goes and what it does once it leaves. As I said, it is a game of compromises and trade-offs.
Ironically, the species and systems that we study also live a life of compromises and trade-offs. In one of this week’s readings, Mridula Srinivasan very eloquently starts her chapter entitled ‘Predator/Prey Decisions and the Ecology of Fear’ in Bernd Würsig’s ‘Ethology and Behavioral Ecology of Odontocetes’ with the following two sentences: “Animal behaviors are governed by the intrinsic need to survive and reproduce. Even when sophisticated predators and prey are involved, these tenets of behavioral ecology hold.”. Every day, animals must walk the tightrope of finding and consuming enough food to survive and ensure a level of fitness required to reproduce, while concurrently making sure that they do not fall prey to a predator themselves. Krebs & Davies (2012) very ingeniously use the idea of economic analysis of costs and benefits to understand foraging behavior (but also behavior in general). While foraging, individuals not only have to assess potential risk (Fig. 1) but also decide whether a certain prey patch or item is profitable enough to invest energy into obtaining it (Fig. 2).
Leigh’s class has been great, not only to learn about foundational theories but to then also apply them to each of our study species and systems. It has been exciting to construct hypotheses based on the readings and then dissect them as a group. As an example, Sih’s 1984 paper on the behavioral response race of predators and prey prompted a discussion on responses of predators and prey to one another and how this affects their spatial distributions. Sih posits that since predators target areas with high prey densities, and prey will therefore avoid areas that predators frequent, their responses are in conflict with one another. Resultantly, there will be different outcomes depending on whichever response dominates. If the predator’s response dominates (i.e. predators are able to seek out areas of high prey density before prey can respond), then predators and prey will have positively correlated spatial distributions. However, if the prey responses dominate, then the spatial distributions of the two should be negatively correlated, as predators will essentially always be ‘one step behind’ the prey. Movement is most often the determinant factor to describe the strength of these relationships.
Video 1. Zooplankton closest to the camera will jump or dart away from it. Source: GEMM Lab.
So, let us think about this for gray whales and their zooplankton prey. The latter are relatively immobile. Even though they dart around in the water column (I have seen them ‘jump’ away from the GoPro when we lower it from the kayak on several occasions; Video 1), they do not have the ability to maneuver away fast or far enough to evade a gray whale predator moving much faster. As such, the predator response will most likely always be the strongest since gray whales operate at a scale that is several orders of magnitude greater than the zooplankton. However, the zooplankton may not be as helpless as I have made them seem. Based on our field observations, it seems that zooplankton often aggregate beneath or around kelp. This behavior could potentially be an attempt to evade predators as the kelp and reef crevices may serve as a refuge. So, in areas with a lot of refuges, the prey response may in fact dominate the relationship between gray whales and zooplankton. This example demonstrates the importance of habitat in shaping predator-prey interactions and behavior. However, we have often observed gray whales perform “bubble blasts” in or near kelp (Video 2). We hypothesize that this behavior could be a foraging tactic to tip the see-saw of predator-prey response strength back into their favor. If this is the case, then I would imagine that gray whales must decide whether the energetic benefit of eating zooplankton hidden in kelp refuges outweighs the energy required to pursue them (Fig. 2). On top of all these choices, are the potential risks and threats of boat traffic, fishing gear, noise, and potential killer whale predation (Fig. 1). Bringing us back to the analogy of economic analysis of costs and benefits to predator-prey relationships. I never realized it so clearly before, but gray whales sure do have a lot of decisions to make in a day!
Video 2. Drone footage of a gray whale foraging in kelp and performing a “bubble blast” at 00:40. Footage captured under NMFS permit #21678. Source: GEMM Lab.
Trying to tease apart these nuanced dynamics is not easy when I am unable to simply ask my study subjects (gray whales) why they decided to abandon a patch of zooplankton (Were the zooplankton too hard to obtain because they sought refuge in kelp, or was the patch unprofitable because there were too few or the wrong kind of zooplankton?). Or, why do gray whales in Oregon risk foraging in such nearshore coastal reefs where there is high boat traffic (Does their need for food near the reefs outweigh this risk, or do they not perceive the boats as a risk?). So, instead, we must set up specific hypotheses and use these to construct a thought-out and informed study design to best answer our questions (Mann 2000). For the past few weeks, I have spent a lot of time familiarizing myself with spatial packages and functions in R to start investigating the relationships between zooplankton and kelp hidden in the data we have collected over 4 years, to ultimately relate these patterns to gray whale foraging. I still have a long and steep journey before I reach the peak but once I do, I hope to have answers to some of the questions that the Cetacean Behavioral Ecology class has inspired.
Literature cited
Krebs, J. R., and N. B. Davies. 2012. Economic decisions and the individual in Davies, N. B. et al., eds. An introduction to behavioral ecology. John Wiley & Sons, Oxford.
Mann, J. 2000. Unraveling the dynamics of social life: long-term studies and observational methods in Mann, J., ed. Cetacean societies: field studies of dolphins and whales. University of Chicago Press, Chicago.
Sih, A. 1984. The behavioral response race between predator and prey. The American Naturalist 123:143-150.
Srinivasan, M. 2019. Predator/prey decisions and the ecology of fear in Würsig, B., ed. Ethology and ecology of odontocetes. Springer Nature, Switzerland.
By: Alexa Kownacki, Ph.D. Student, OSU Department of Fisheries and Wildlife,
Geospatial Ecology of Marine Megafauna Lab
We live in an interesting time. Many of us academic
scientists sit in the confines of our homes, reading scientific papers,
analyzing years-worth of data, working through a years-worth of house projects,
or simply watching Netflix. While we are confined to a much smaller area,
wildlife is not.
During this challenging situation we have unique
opportunities to study what happens when people are not outside for recreation.
All of us who feel trapped inside our homes are not only saving human lives, we
are changing ecosystems. Humans are constantly molding our ecosystems on fine
and grand scales, from xeriscaping our lawns with native, drought-resistant
plants to developing large plots of land for new homes. We manipulate nature,
for better or for worse.
So, what happens when we change our behavior? Rather than
driving, we’re gardening, instead of playing at parks, we’re playing board
games at our kitchen tables; we as a society are completely changing our
habitat-use patterns. When any top predator changes its habitat-use, switches
niches, or drastically changes its behaviors, there are top-down ecosystem
effects. When one species changes its behavior, there are major downstream
impacts on predation, foraging, diet, and habitat use. For example, when
bluegill sunfish underwent large shifts in both diet and habitat, major
predator-mediated habitat use changes in other species occurred (Mittelbach
1986). There are multiple studies describing the impacts of human-mediated
drivers on ecosystems worldwide. In coastal environments, anthropogenic
activities, specifically shipping, industry, and urban development, dramatically
change both the coastal and marine ecosystems (Mead et al. 2013).
The highly developed coastline along Los Angeles, CA is a prime example of urban development. (Image source: LA Magazine.)
By far the most pronounced example of how an international halt on travel can alter ecosystems comes from the tragic terrorist attacks on September 11, 2001. Prior to this current, viral pandemic, the events following 9/11 were the first time that nearly all major transit stopped in the USA—including airplanes and major shipping traffic. This halt created a unique opportunity to study some of the secondary impacts, such as a reduction in shipping traffic noise, on cetaceans. Following 9/11, there was a six decibel decrease in underwater noise that co-occurred with a decrease in stress hormones of endangered North Atlantic right whales (Rolland et al. 2012). When I first read about this study, my first thought was “leave it to scientists to make the best out of a terrible situation.” Truly, learning from nature, even in the darkest of days, is an incredible skillset. Research like this inspires me to ask questions about what changes are happening in ecosystems now because of recent events. For example, the entire port of San Diego, its beaches and bays, are closed for all recreational activity and I wonder how this reduction in traffic is similar to the post-9/11 study but on bottlenose dolphins, gray whales, and pinnipeds that are coast-associated. Are urban and suburban neighborhoods slowly becoming more rural and making space for wildlife again?
My dad lives in a suburban neighborhood of San Diego, CA. In the past few weeks, his “Ring doorbell camera” captured a bobcat walking along the raised brick path multiple times. (Media source: Eric Kownacki)
There is increasing news coverage on wild animals “taking over” cities. Dr. Leila Lemos touched on this earlier with her blog post centering on how academics are changing their means of teaching, conferencing, and learning. There are photos of wild goats running through the streets of Wales, UK, coyotes roaming the streets of San Francisco, CA, USA, monkeys swarming the streets in Thailand, pumas wandering the streets of Santiago, Chile, and Sika deer peering into empty restaurants in Nara, Japan (Colarossi 2020). In reality, this wildlife was likely part of the ecosystem prior to the formation of these cities but was forced out of the more urban centers. As we sit in our homes, rather than looking bleakly onto empty streets, we can search for wildlife, create a backyard birding competition with your friends, guess which flowers will bloom first, and ask questions of our changing ecosystems.
Coyote at a park in northern California with the San Francisco Golden Gate Bridge in the background. (Image source: u/beccatravels via Reddit)
Mead, A., Griffiths, C.L., Branch, G.M.,
McQuaid, C.D., Blamey, L.K., Bolton, J.J., Anderson, R.J., Dufois, F., Rouault,
M., Froneman, P.W. and Whitfield, A.K., 2013. Human-mediated drivers of
change—impacts on coastal ecosystems and marine biota of South Africa. African
Journal of Marine Science, 35(3), pp.403-425.
Mittelbach, Gary. 1986. Predator-mediated
habitat use: some consequences for species interactions. Environ Biol
Fish16, 159–169. https://doi.org/10.1007/BF00005168
Rolland, R.M., Parks, S.E., Hunt, K.E.,
Castellote, M., Corkeron, P.J., Nowacek, D.P., Wasser, S.K. and Kraus, S.D.,
2012. Evidence that ship noise increases stress in right whales. Proceedings
of the Royal Society B: Biological Sciences, 279(1737),
pp.2363-2368.
I want to start my post this week with a disclaimer – I am not a virologist or an epidemiologist. My knowledge and understanding on what a virus is, how it changes and spreads, and predicting its trajectory, is very limited (though it has definitely improved in recent weeks). Nevertheless, I did not want that to stop me from shifting my focus and time currently spent reading about a certain virus in humans, to thinking about viruses in marine mammals. So, after several hours of reading papers and reports, I believe I have a good enough grasp on viruses in marine mammals to write a blog post on this topic.
To answer the question in my title – yes, marine mammals can get coronavirus! Coronaviruses have been detected in several marine mammals – mostly in captive ones (harbor seal, beluga whale, Indo-Pacific bottlenose dolphin), but it was also detected in a wild harbor seal1. It is at this point where I am going to step back from marine mammals for a moment and give a very short ‘lesson’ on viruses.
Viruses are microscopic infectious agents that replicate inside living cells of organisms. They have the ability to infect all forms of life – anything from bacteria to plants to animals to humans. Nothing is excluded. Viruses are classified similarly to how living organisms are classified. Try to think back to middle school science when your teacher used mnemonic devices like, “Kids prefer candy over fancy green salad” or “Kings play chess on fine glass surfaces”, to get you to remember the Kingdom-Phylum-Class-Order-Family-Genus-Species classification. Well, viruses have almost the same classification tree. The only difference is that instead of Kingdom at the top, viruses have a Realm. As of 2019, the International Committee on Taxonomy of Viruses (ICTV) has defined 5,560 species of viruses in over 1,000 genera and 150 families. Different species of virus are classified based on their genomic material and key elements of structure and replication. That is as far as I am going to go with virus background – back to marine mammals!
Grey seal hauled out along the west coast of the U.K. Source: L. Hildebrand.
So, yes, coronaviruses have been detected in marine mammals before. But, no, they were not the same species of coronavirus that is currently spreading across the globe in humans. Coronavirus, or Coronaviridae, is a family of viruses that contains around 40 species. However, coronavirus is not the family that has plagued marine mammals the most since research on marine mammal diseases began. The infectious disease that I have found to be the most common and recurring in marine mammals is morbillivirus and I will therefore focus on that virus for the rest of this post.
Morbillivirus is a genus of viruses in the family Paramyxoviridae and hosts of this genus include humans, dogs, cats, cattle, seals, and cetaceans. There are seven described species of morbillivirus, three of which have been detected in marine mammals, namely canine distemper virus (CDV), cetacean morbillivirus (CeMV), and phocine distemper virus (PDV). The earliest, traceable case of morbillivirus in a marine mammal occurred in 1982 in bottlenose dolphins in the Indian and Banana Rivers in Florida2. This case was followed by hundreds of others in subsequent years all along the Atlantic U.S. coast and resulted in the first Unusual Mortality Event (UME; 1987-1988) that was concluded to have been caused by morbillivirus (Table 1).
Table 1. Unusual Mortality Events (UMEs) of marine mammals in the U.S. where the cause was determined to be or is suspected to be morbillivirus. Data obtained from NOAA Fisheries.
Interestingly, at the same time as this 1980s morbillivirus in the US, the first documented marine mammal morbillivirus epidemic occurred in Europe in the North Sea. This outbreak led to the death of more than 23,000 harbor seals, which accounted for roughly 60% of all North Sea harbor seals at the time3. The virus that was isolated from the stranded seals in the North Sea was similar to CDV but not exactly the same. Resultantly, it was described as a new species of morbillivirus and it was therefore the first outbreak of PDV. Another interesting thing about this case in Europe is that while the infection originated at the Danish island of Anholt, new centers of infection appeared quite far from this first epicenter within a relatively short amount of time (~3-4 weeks) from the initial outbreak, some as far as the Irish Sea (~2,000 km away; Figure 1). Harbor seals typically have a limited home range and do not travel such distances, leading scientists to speculate that grey seals may have been a carrier of the virus and transported it from Anholt to haul-out sites in the Irish Sea. Mixed species haul-out sites of harbor and grey seals are very common across the North Sea and is the most logical explanation for the rapid spread of the virus across such distances.
Figure 1. Map of the North Sea showing Anholt island (red marker) and the Irish Sea (white circle).
Harbor seals seem to be the most susceptible to PDV based on all documented cases of PDV outbreaks, however the reason for this pattern remains unknown1. While CDV has only been detected in Baikal and Caspian seals, CeMV has occurred in a larger number of cetaceans including harbor porpoises, striped, bottlenose, Guiana and Fraser’s dolphins, pilot whales, and a minke whale. This list is not extensive as morbillivirus has been found in 23 of the 90 cetacean species. In fact, it has been suggested that CeMV should be divided into more than one species as the morbilliviruses detected in the Northern Hemisphere show significant divergence from those found in the Southern Hemisphere.
Transmission is believed to mostly occur horizontally, meaning that the morbillivirus is passed from one individual to another. This transfer happens when one individual inhales the aerosolized virus breathed out by an infected individual. This is likely the reason why odontocete and pinniped groups are most affected due to their social group behavior and/or high density of individuals within groups4. However, vertical transmission has also been suggested as a possible transmission route as morbillivirus antigens have been detected in the mammary glands of bottlenose dolphins along the U.S. Atlantic Coast5 and striped dolphins in the Mediterranean Sea affected by CeMV6. Thus, it has been postulated that CeMV infected females could transmit the infection to their fetuses and neonates in utero, as well as to their calves during lactation.
Bottlenose dolphins populations have been involved in several UME events related to morbillivirus along the U.S. coasts (Table 1). Source: L. Hildebrand. Image captured under NMFS permit #19116.
Morbilliviruses mostly affect the respiratory and neurologic systems in marine mammals, wherein affected individuals may display ocular and naval discharge, erratic swimming, respiratory distress, raised body temperature, and/or cachexia (weakness and wasting away of the body due to severe illness). However, most diagnoses occur post-mortem. Some individuals may survive the initial acute infection of morbillivirus, yet the general weakening of the immune system will make individuals more susceptible to other infections, diseases, and disturbance events7.
It is impossible to know whether marine mammals take precautions when a virus has taken grip of a group or population, or if marine mammals even have an awareness of such things occurring. There obviously is no such thing as an emergency room or a doctor in the lives of marine mammals, but do individuals perhaps demonstrate social distancing by increasing the space between each other when traveling in groups? Do groups decrease their traveling distances or foraging ranges to isolate themselves in a smaller area? Are sick individuals ‘quarantined’ by being forced out of a group? These are just some of the questions I have been asking myself while working from home (day 16 for me now). I hope you are all staying safe and healthy and have enjoyed distracting yourselves from thinking about one virus to learn about another in a different kind of mammal.
Literature cited
1 Bossart, G. D., and P. J. Duignan. 2018. Emerging viruses in marine mammals. CAB Reviews 13(52): doi:10.1079/PAVSNNR201913052.
2 Duignan, P. J., C. House, D. K. Odell, R. S. Wells, L. J. Hansen, M. T. Walsh, D. J. St. Aubin, B. K. Rima, and J. R. Geraci. 1996. Morbillivirus infection in bottlenose dolphins: evidence for recurrent epizootics in the western Atlantic and Gulf of Mexico. Marine Mammal Science 12(4):499-515.
3 Härkönen, T., R. Dietz, P. Reijnders, J. Teilmann, K. Harding, A. Hall, S. Brasseur, U. Siebert, S. J. Goodman, P. D. Jepson, T. D. Rasmussen, and P. Thompson. 2006. A review of the 1988 and 2002 phocine distemper virus epidemics in European harbor seals. Diseases of Aquatic Organisms 68:115-130.
4 Van Bressem, M-F., P. J. Duignan, A. Banyard, M. Barbieri, K. M. Colegrove, S. De Guise, G. Di Guardo, A. Dobson, M. Domingo, D. Fauquier, A. Fernandez, T. Goldstein, B. Grenfell, K. R. Groch, F. Gulland, B. A. Jensen, P. D. Jepson, A. Hall, T. Kuiken, S. Mazzariol, S. E. Morris, O. Nielsen, J. A. Raga, T. K. Rowles, J. Saliki, E. Sierra, N. Stephens, B. Stone, I. Tomo, J. Wang, T. Waltzek, and J. F. X. Wellehan. 2014. Cetacean morbillivirus: current knowledge and future directions. Viruses 6(12):5145-5181.
5 Schulman, F. Y., T. P. Lipscomb, D. Moffett, A. E. Krafft, J. H. Lichy, M. M. Tsai, J. K. Taubenberger, and S. Kennedy. 1997. Histologic, immunohistochemical, and polymerase chain reaction studies of bottlenose dolphins from the 1987-1988 United States Atlantic coast epizootic. Veterinary Pathology 34(4):288-295.
6 Domingo, M., J. Visa, M. Pumarola, A. J. Marco, L. Ferrer, R. Rabanal, and S. Kennedy. 1992. Pathologic and immunocytochemical studies of morbillivirus infection in striped dolphins (Stenella coeruleoalba). Veterinary Pathology 29(1):1-10.
7 Wellehan, J., and G. Cortes-Hinojosa. 2019. Marine Mammal Viruses. Fowler’s Zoo and Wild Animal Medicine Current Therapy 9:597-602.
By: Alexa Kownacki, Ph.D. Student, OSU Department of Fisheries and Wildlife, Geospatial Ecology of Marine Megafauna Lab
Human-wildlife interactions have occurred since people first inhabited the Earth. However, today, when describing human-wildlife interactions specifically in relation to dolphins, frequently we hear about ‘conflicts’. Interactions between fisheries and dolphins that lead to bycatch or depredation (stealing bait/catching from gear) are particularly common. But, symbiotic relationships with dolphin species and certain human groups can also be mutualistic, with both groups benefitting. These symbiotic relationships have been around for hundreds, if not thousands of years.
A depiction of Aboriginal Australians using nets to catch fish in a small inlet with the assistance of coastal dolphins. (Image source: Our Pacific Ocean)
In eastern Australia, cooperative fishing interactions occur
between Aboriginal Australians and dolphins—both bottlenose dolphins and orcas.
In Burleigh Heads National Park, Queensland, AUS, the dolphins are thought to
help the local indigenous Kombemerri (saltwater) people hunt for fish. Indigenous
stories recall men wading into the water with their spears and nets. Then, many
of the men would hit the surface waters to make noises with the splashes.
Underwater, this sound was amplified and then the dolphins would begin chasing
the fish toward the men and their nets (Neil 2002). Aboriginal Australians,
especially those in eastern Australia have an emotional and spiritual
connection to both dolphins and orcas. There are widespread accounts of cooperation
between indigenous people and small cetaceans on the eastern Australian
coastline, which create both context and precedent for the economic and
emotional objectives to contemporary human-dolphin interactions such as dolphin
provisioning (Neil 2002).
Dolphins and fishermen work together in Laguna, Brazil, to catch mullet. (Image Source: Fábio Daura-Jorge)
In the coasts off of Laguna, Brazil, bottlenose dolphins and local fishermen cooperatively fish while tourists gather to watch. Previously, PhD candidate Leila Lemos wrote about these interactions in a blog post. Like many groups of socializing dolphins, these dolphins have a unique whistle to recognize each other. The waters surrounding Laguna, Brazil are murky, turbid and dark green to the point where the fisherman cannot see any of the fish in the water. As the fishermen wade into the murky waters, bottlenose dolphins chase shoals of mullet toward the shore. Then the dolphins tail slap or abruptly dive, “signaling” the fishermen to cast their nets. Research has shown that when the fishermen “work with” the dolphins, both the dolphins and the people catch more, larger fish (Roman 2013). One fisherman claims it is not worth fishing unless the dolphins are around (Roman 2013). Here, the fishermen know the dolphins based on their markings. They know which dolphins participate in the different parts of hunting as well—which dolphin initiates the tail slap, which dolphin usually circles the fish, and which drive the fish towards the coastline. After the dolphins round up and chase the fish for the fishermen and themselves, there is no “reward” from the fishermen for the dolphins—no fish tossed their way. Scientists also found there is a difference in whistle structure between cooperative and non-cooperative dolphin groups (Preston 2017).
A fisherman in Brazil throws a net after dolphins chase mullet into the shore. (Image Source: Leo Francini:Alamy Stock Photo)
Along most coastlines worldwide, humans and dolphins are
competitors. Dolphins are seen as thieves who steal fish out of nets, or get
caught in their gear and ruin fishing opportunities. Thus, dolphins are often unwelcome
near fishing communities. Such negative interactions sometimes lead to
human-caused fatalities of dolphin from gunshots or stabbings, thought to be
from angry fishermen. Yet, in this same
world, fishermen thank the dolphins for bringing their catch to them. Clearly,
both humans and dolphins share high intelligence levels and skills in fishing.
If it is a matter of two minds are better than one, then I think indigenous
communities figured this equation out first: working with the dolphins, and not
against, can better feed their people.
Citations:
Neil, David.
(2002). Cooperative fishing interactions between Aboriginal Australians and
dolphins in eastern Australia. Anthrozoos: A Multidisciplinary Journal of The
Interactions of People & Animals. 15. 10.2752/089279302786992694.
Preston, Elizabeth.
“Dolphins That Work with Humans to Catch Fish Have Unique Accent.” New
Scientist, 2 Oct. 2017,
www.newscientist.com/article/2149139-dolphins-that-work-with-humans-to-catch-fish-have-unique-accent/.
Roman, Joe. “Fishing with
Dolphins: An astonishing cooperative venture in which every species wins but
the fish.” Slate Magazine, 31 Jan. 2013,
slate.com/technology/2013/01/fishing-with-dolphins-symbiosis-between-humans-and-marine-mammals-to-catch-more-fish.html.