“Why don’t you just automate it?” This is a question I am frequently asked when I tell someone about my work. My thesis involves watching many hours of drone footage of gray whales and meticulously coding behaviors, and there are plenty of days when I have asked myself that very same question. Streamlining my process is certainly appealing and given how wide-spread and effective machine learning methods have become, it is a tempting option to pursue. That said, machine learning is only appropriate for certain research questions and scales, and it’s important to consider these before investing in using a new tool.
The application of machine learning methods to behavioral ecology is called computational ethology (Anderson & Perona, 2014). To identify behaviors from videos, the model tracks individuals across video frames and identifies patterns of movement that form a behavior. This concept is similar to the way we identify a whale as traveling if it’s moving in a straight line and as foraging if it’s swimming in circles within a small area (Mayo & Marx, 1990, check out this blog to learn more). The level of behavioral detail that the model is able to track depends on the chosen method (Figure 1, Pereira et al., 2020). These methods range from tracking each animal as a simple single point (called a centroid) to tracking the animal’s body positioning in 3D (this method is called pose estimation), which range from providing less detailed to more detailed behavior definitions. For example, tracking an individual as a centroid could be used to classify traveling and foraging behaviors, while pose estimation could identify specific foraging tactics.
Figure 1. Figure from Pereira et al. (2020) illustrating the different methods of animal behavior tracking that are possible using machine learning.
Pose estimation involves training the machine learning algorithm to track individual anatomical features of an individual (e.g., the head, legs, and tail of a rat), meaning that it can define behaviors in great detail. A behavior state could be defined as a combination of the angle between the tail and the head, and the stride length.
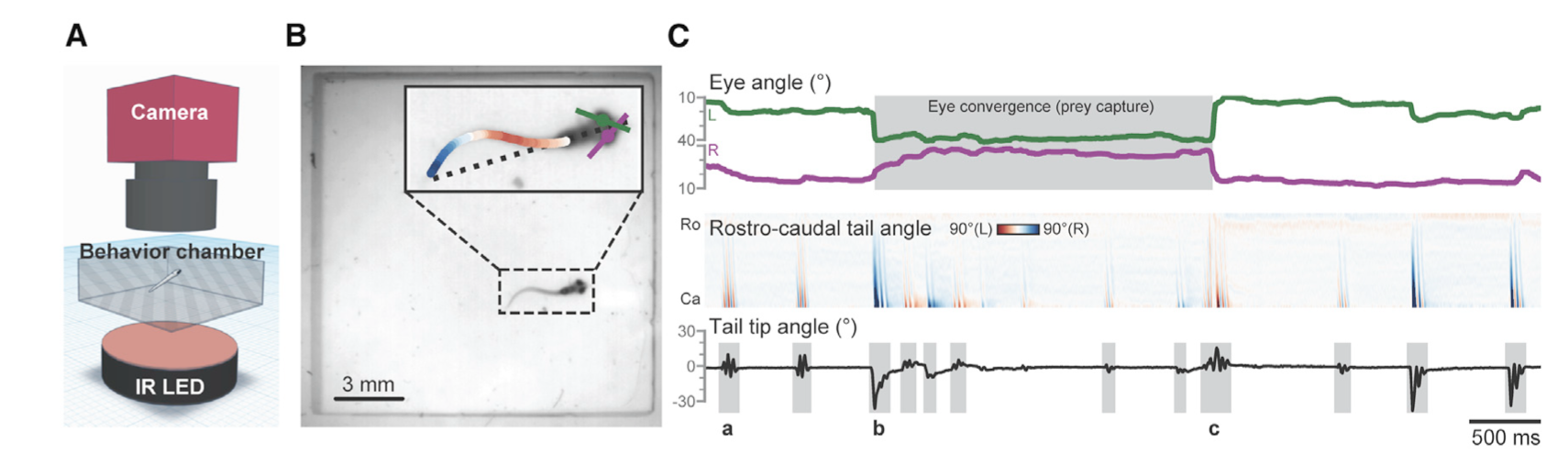
For example, Mearns et al. (2020) used pose estimation to study how zebrafish larvae in a lab captured their prey. They tracked the tail movements of individual larvae when presented with prey and classified these movements into separate behaviors that allowed them to associate specific behaviors with prey capture (Figure 2). The authors found that these behaviors occurred in a specific sequence, that the behaviors kept the prey within the larvae’s line of sight, and that the sequence was triggered by visual cues. In fact, when they removed the visual cue of the prey, the larvae terminated the behavior sequence, meaning that the larvae are continually choosing to do each behavior in the sequence, rather than the sequence being one long behavior event that is triggered only by the initial visual cue. This study is a good example of the applicability of machine learning models for questions aimed at kinematics and fine-scale movements. Pose estimation has also been used to study the role of facial expression and body language in rat social communication (Ebbesen & Froemke, 2021).
Figure 2. Excerpt from figure 1 of Mearns et al. (2020) illustrating (A) the camera set up for their experiment, (B) how the model tracked the eye angles and tail of the larvae fish, (C) the kinematics extracted from the footage. In panel (C) the top plot shows how the eyes converged on the same object (the prey) during prey capture event, the middle plot shows when the tail was curved to the left or the right, and the bottom plot shows the angle of the tail tip relative to the body.
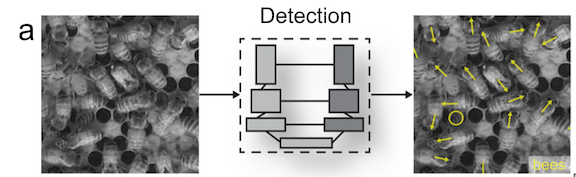
While previous machine learning methods to track animal movements required individuals to be physically marked, the current methods can perform markerless tracking (Pereira et al., 2020). This improvement has broadened the kinds of studies that are possible. For example, Bozek et al., (2021) developed a model that tracked individuals throughout an entire honeybee colony and showed that certain individual behaviors were spatially distributed within the colony (Figure 3). Machine learning enabled the researchers to track over 1000 individual bees over several months, a task that would be infeasible for someone to do by hand.
Figure 3. Excerpt from figure 1 of Bozek et al., (2021) showing how individual bees and their trajectories were tracked.
These studies highlight that the potential benefits of using machine learning when studying fine scale behaviors (like kinematics) or when tracking large groups of individuals. Furthermore, once it’s trained, the model can process large quantities of data in a standardized way to free up time for the scientists to focus on other tasks.
While machine learning is an exciting and enticing tool, automating behavior detection via machine learning could be its own PhD dissertation. Like most things in life, there are costs and benefits to using this technique. It is a technically difficult tool, and while applications exist to make it more accessible, knowledge of the computer science behind it is necessary to apply it effectively and correctly. Secondly, it can be tedious and time consuming to create a training dataset for the model to “learn” what each behavior looks like, as this step involves manually labeling examples for the model to use.
As I’ve mentioned in a previous blog, I came quite close to trying to study the kinematics of gray whale foraging behaviors but ultimately decided that counting fluke beats wasn’t necessary to answer my behavioral research questions. It was important to consider the scale of my questions (as described in Allison’s blog) and I think that diving into more fine-scale kinematics questions could be a fascinating follow-up to the questions I’m asking in my PhD.
For instance, it would be interesting to quantify how gray whales use their flukes for different behavior tactics. Do gray whales in better body condition beat their flukes more frequently while headstanding? Does the size of the fluke affect how efficiently they can perform certain tactics? While these analyses would help quantify the energetic costs of different behaviors in better detail, they aren’t necessary for my broad scale questions. Consequently, taking the time to develop and train a pose estimation machine learning model is not the best use of my time.
That being said, I am interested in applying machine learning methods to a specific subset of my dataset. In social behavior, it is not only useful to quantify the behaviors exhibited by each individual but also the distance between them. For example, the distance between a mom and her calf can be indicative of the calves’ dependence on its mom (Nielsen et al., 2019). However, continuously measuring the distance between two individuals throughout a video is tedious and time intensive, so training a machine learning model could be an effective use of time. I plan to work with an intern this summer to develop a machine learning model to track the distance between pairs of gray whales in our drone footage and then relate this distance data with the manually coded behaviors to examine patterns in social behavior (Figure 4). Stay tuned to learn more about our progress!
Figure 4. A mom and calf pair surfacing together. Image collected under NOAA/NMFS permit #21678
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly alert when we make a new post! Just add your name into the subscribe box on the left panel.
References
Anderson, D. J., & Perona, P. (2014). Toward a Science of Computational Ethology. Neuron, 84(1), 18–31. https://doi.org/10.1016/j.neuron.2014.09.005
Bozek, K., Hebert, L., Portugal, Y., Mikheyev, A. S., & Stephens, G. J. (2021). Markerless tracking of an entire honey bee colony. Nature Communications, 12(1), 1733. https://doi.org/10.1038/s41467-021-21769-1
Ebbesen, C. L., & Froemke, R. C. (2021). Body language signals for rodent social communication. Current Opinion in Neurobiology, 68, 91–106. https://doi.org/10.1016/j.conb.2021.01.008
Mayo, C. A., & Marx, M. K. (1990). Surface foraging behaviour of the North Atlantic right whale, Eubalaena glacialis , and associated zooplankton characteristics. Canadian Journal of Zoology, 68(10), 2214–2220. https://doi.org/10.1139/z90-308
Mearns, D. S., Donovan, J. C., Fernandes, A. M., Semmelhack, J. L., & Baier, H. (2020). Deconstructing Hunting Behavior Reveals a Tightly Coupled Stimulus-Response Loop. Current Biology, 30(1), 54-69.e9. https://doi.org/10.1016/j.cub.2019.11.022
Nielsen, M., Sprogis, K., Bejder, L., Madsen, P., & Christiansen, F. (2019). Behavioural development in southern right whale calves. Marine Ecology Progress Series, 629, 219–234. https://doi.org/10.3354/meps13125
Pereira, T. D., Shaevitz, J. W., & Murthy, M. (2020). Quantifying behavior to understand the brain. Nature Neuroscience, 23(12), 1537–1549. https://doi.org/10.1038/s41593-020-00734-z
Over six field seasons the GEMM lab team has conducted nearly 500 drone flights over gray whales, equaling over 100 hours of footage. These hours of footage are the central dataset for my PhD dissertation, so it’s up to me to process them all. This process can be challenging, tedious, and daunting, but it is also quite fun and a privilege to be the one person who gets to watch all the footage. It’s fascinating to get to know the whales and their behaviors and pick up on patterns. It motivates me to get through this video processing step and start doing the data analysis. Recently, it’s been especially fun to notice patterns that I’ve seen mentioned in the literature. One example is adult social behavior.
There are two categories of social behavior that I’m interested in studying: maternal behavior, defined as interactions between a mom and its calf, and general social behaviors, defined as social interactions between non-mom/calf pairs. In this blog I’ll focus on general social behaviors, but if you’re interested in maternal behavior check out this blog. General social behavior, which I’ll refer to as social behavior moving forward, includes tactile interactions and promiscuous behaviors (Torres et al. 2018; Clip 1). While gray whales in the PCFG range are primarily foraging, researchers have observed increases in social behavior towards the end of the foraging season (Stelle et al., 2008; Torres et al., 2018). We think that this indicates that the whales are starting to focus less on feeding and more on breeding. This tradeoff of foraging vs. socializing time is interesting because it comes at an energetic cost.
Clip 1. Example of social interaction between a male and female gray whale off the coast of Oregon, USA. Collected under NOAA/NMFS permit #21678
Broadly, animals need to balance the energetic demands of survival with those of reproduction. They need to reproduce to pass on their genes, but reproduction is energetically demanding, and animals also need to survive and grow to be able to reproduce. The decision to reproduce is costly because reproduction requires energetic investment and time investment since animals do not forage (gaining energy) when they are socializing. Consequently, only animals with sufficient energy reserves (i.e., body condition) to invest in reproduction actually engage in reproduction. Given these costs associated with reproduction, we expect to see a relationship between social behavior and body condition (Green, 2001) with mainly animals in good body condition engaging in social behavior because these animals have sufficient reserves to sustain the cost. Furthermore, since body condition is an indicator of foraging success and prey availability, environmental conditions can also affect social behavior and reproduction through this pathway.
Rahman et al. (2014) used a lab experiment to study the relationship between nutritional stress and male guppy courtship behavior (Figure 1). In their experiment they tested for the effects of both decreased diet quantity and quality on the frequency of male courtship behaviors. Rahman et al (2014) found that individuals in the low-quantity group were significantly smaller than those in the high-quality group and that diet quantity had a significant effect on the frequency of courtship behaviors. Males fed a low-quantity diet performed fewer courtship behaviors. Interestingly, there was no significant effect of diet quality on courtships behavior, although there was some evidence of an interaction effect, which suggests that within the low-quantity group, males fed with high-quality food performed more courtship behaviors that those fed with low-quality food. This study is interesting because it shows how foraging success (diet quantity and quality) can affect courting behavior.
Figure 1. A guppy (Rahman et al., 2013)
However, guppies are not the ideal species for comparison to gray whales because gray whales and guppies have quite different life history traits. A more fitting comparison would be with an example species with more in common with gray whales, such as viviparous capital breeders. Viviparous animals develop the embryo inside the body and give live birth. Capital breeders forage to build energy reserves and then rely on those energy reserves during reproduction. Surprisingly, I found asp vipers to be a good example species for comparison to gray whales.
Asp vipers (Figure 2) are viviparous snakes who are considered capital breeders because they forage prior to hibernation, and then begin reproduction immediately following hibernation without additional foraging. Naulleau & Bonnet (1996) conducted a field study on female asp vipers to determine if there was a difference in body condition at the start of the breeding season between females who reproduced or not during that season. To do this they marked individuals and measured their body condition at the start of the breeding season and then recaptured those individuals at the end of the breeding season and recorded whether the individual had reproduced. Interestingly, they found that there was a strongly significant difference in body condition between females that did and did not reproduce. In fact, they discovered that no female below a certain body condition value reproduced, meaning that they found a body condition threshold for reproduction.
Figure 2. An asp viper
Additionally, a study on water pythons found that their body condition threshold for reproduction shifted over time in response to prey availability (Madsen & Shine, 1999). These authors found that females lowered their threshold after several consecutive years of poor prey availability. These studies are really exciting to me because they address questions that the GRANITE project team is interested in tackling.
Understanding the relationship between body condition and reproduction in gray whales is an important puzzle piece for our work. The aim of the GRANITE project is to understand how the effects of stressors on individual whales scales up to population level impacts (read Lisa’s blog to learn more). Reproduction rates play a big role in population dynamics, so it is important to understand what factors affect reproduction. Since we’re studying these whales on their foraging grounds, assessing body condition provides an important link between foraging behavior and reproduction.
For example, if an individual’s response to a stressor is to forage less, that may lead to poorer body condition, meaning that they may be less likely to reproduce. While reduced reproduction in one individual may not have a big effect on the population, the same response from multiple individuals could impact the population’s dynamics (i.e., increasing or decreasing abundance). Understanding these different relationships between behavior, body condition, and reproduction rates is a big undertaking, but it’s exciting to be a member of the GRANITE team as this strong group of scientists works to bring together different data streams to work on this big picture question. We’re all deep into data processing right now so stay tuned over the next few years to learn more about gray whale social behavior and to find out if fat whales are more social than skinny whales.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get weekly updates and more! Just add your name into the subscribe box on the left panel.
References
Green, A. J. (2001). Mass/Length Residuals: Measures of Body Condition or Generators of Spurious Results? Ecology, 82(5), 1473–1483. https://doi.org/10.1890/0012-9658(2001)082[1473:MLRMOB]2.0.CO;2
Madsen, T., & Shine, R. (1999). The adjustment of reproductive threshold to prey abundance in a capital breeder. Journal of Animal Ecology, 68(3), 571–580. https://doi.org/10.1046/j.1365-2656.1999.00306.x
Naulleau, G., & Bonnet, X. (1996). Body Condition Threshold for Breeding in a Viviparous Snake. Oecologia, 107(3), 301–306.
Rahman, M. M., Kelley, J. L., & Evans, J. P. (2013). Condition-dependent expression of pre- and postcopulatory sexual traits in guppies. Ecology and Evolution, 3(7), 2197–2213. https://doi.org/10.1002/ece3.632
Rahman, M. M., Turchini, G. M., Gasparini, C., Norambuena, F., & Evans, J. P. (2014). The Expression of Pre- and Postcopulatory Sexually Selected Traits Reflects Levels of Dietary Stress in Guppies. PLOS ONE, 9(8), e105856. https://doi.org/10.1371/journal.pone.0105856
Stelle, L. L., Megill, W. M., & Kinzel, M. R. (2008). Activity budget and diving behavior of gray whales (Eschrichtius robustus) in feeding grounds off coastal British Columbia. Marine Mammal Science, 24(3), 462–478. https://doi.org/10.1111/j.1748-7692.2008.00205.x
Torres, L. G., Nieukirk, S. L., Lemos, L., & Chandler, T. E. (2018). Drone up! Quantifying whale behavior from a new perspective improves observational capacity. Frontiers in Marine Science, 5(SEP). https://doi.org/10.3389/fmars.2018.00319
Dr. Alejandro Fernández Ajó, OSU Department of Fisheries, Wildlife, & Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
Rises in ocean temperatures can lead to multiple alterations in marine ecosystems, including the increase and the frequency of Harmful Algal Blooms (HABs). HABs are characterized by the rapid growth of toxin-producing species of algae that can be harmful to people, animals, and the local ecology, even causing death in severe cases. Species of marine diatom within the genus Pseudo-nitzschia and Nitzschia can form HABs when they produce domoic acid (DA), a potent neurotoxin responsible for amnesic shellfish poisoning (D’Agostino et al., 2018, 2017).
Figure 1. Southern right whale (E. australis) mother and calf swimming at the gulfs of Peninsula Valdes, Argentina, during a phytoplankton bloom. Photo: Mariano Sironi / Instituto de Conservacion de Ballenas de Argentina.
During HABs, DA is transferred to higher organisms through the pelagic food web and is accumulated by intermediate vectors, such as copepods, euphausiids (i.e., krill), shellfish, and fish. As this neurotoxin affects top predators, DA poisoning poses a risk to the safety and health of humans and wildlife. This neurotoxin has caused mortality in many marine mammal species, including both pinnipeds and cetaceans (Gulland 1999; Lefebvre et al. 1999; Fire et al. 2010, 2021; Broadwater et al. 2018). In addition, the exposure to DA constitutes a stressor that may affect glucocorticoids (hormones involved in the stress response) concentrations.
The glucocorticoids (GCs; cortisol and corticosterone) are adrenal steroid hormones that maintain the essential functions of metabolism and energy balance in mammals. GCs can increase sharply in response to environmental stressors to elicit physiological and behavioral adaptations by individuals to support survival (Sapolsky et al. 2000; Bornier et al. 2009). However, with the chronic exposure to a stressor, this relationship can reverse, with GCs sometimes declining below its baseline levels (Dickens and Romero, 2013; Fernández Ajó et al., 2018). Moreover, DA can interfere with the stress response in mammals, and cause alterations in their physiological response. DA is an excitatory amino acid analog of glutamate (Pulido 2008), a well-known brain neurotransmitter that play an important role in the activation of the adrenal axis (which in turn regulate the production and secretion of the GCs) and regulate many of the pituitary hormones involved in the stress response (Brann and Mahesh 1994; Johnson et al. 2001). Hence, monitoring GC levels in marine mammals can be a potential useful metric for assessing the physiological impacts of exposure to DA.
Glucocorticoids are traditionally measured in plasma, but given that plasma sampling from free-ranging large whales is currently impossible, alternative sample types such as fecal samples, among others, can be utilized to quantify GCs in large whales (Ajó et al., 2021; Burgess et al., 2018, 2016; Fernández Ajó et al., 2020, 2018; Hunt et al., 2019, 2014, 2006; Rolland et al., 2017, 2005)(Figure 2). The analyses of fecal glucocorticoid metabolites (fGCm) is particularly useful for endocrine assessments of free-swimming whales, with several studies showing that fGCm correlate in meaningful ways with presumed stressors. For example, high levels of fGCm in North Atlantic right whales (NARW, Eubalaena glacialis) and in gray whales (Eschrichtius robustus) correlate with poor body condition (Hunt et al., 2006; Lemos et al., 2021), and fGCm increases were associated with whale entanglements and ship strikes (i.e., Lemos et al., 2020; Rolland et al., 2017).
Figure 2. Alternative samples types can be used to study hormones in large whales. 1-2-3 are sample types that can be obtained from free-living whales and provide a more instantaneous and acute measurement of the whales´ physiology. 4-5 can be obtained at necropsy when the whale is found dead at the beach and provide an integrated measure of the whale physiology that can expand through years or even the lifespan of an individual.
In Península Valdés, Argentina, southern right whales (SRW, E. australis) gather in large numbers to mate and nurse their calves during the austral winter months (Bastida and Rodríguez, 2009). SRWs are capital breeders, largely fasting during the breeding season and instead relying on stored blubber fuel reserves. However, they can occasionally feed on calanoid copepods (D’Agostino et al., 2018, 2016), particularly during the phytoplankton blooms that are dominated by diatoms of the genus Pseudo-nitzschia (Sastre et al. 2007; D’Agostino et al. 2015, 2018). Therefore, feeding SRWs in Península Valdés temporally overlap with these Pseudo-nitzschia blooms (D’Agostino et al. 2018, 2015) and represents a test case for assessing the relationship of DA exposure with GC levels (Figure 3).
Figure 3. Southern right whale (E. australis) skim feeding at the Peninsula Valdes breeding ground. Photo: Lucas Beltranino.
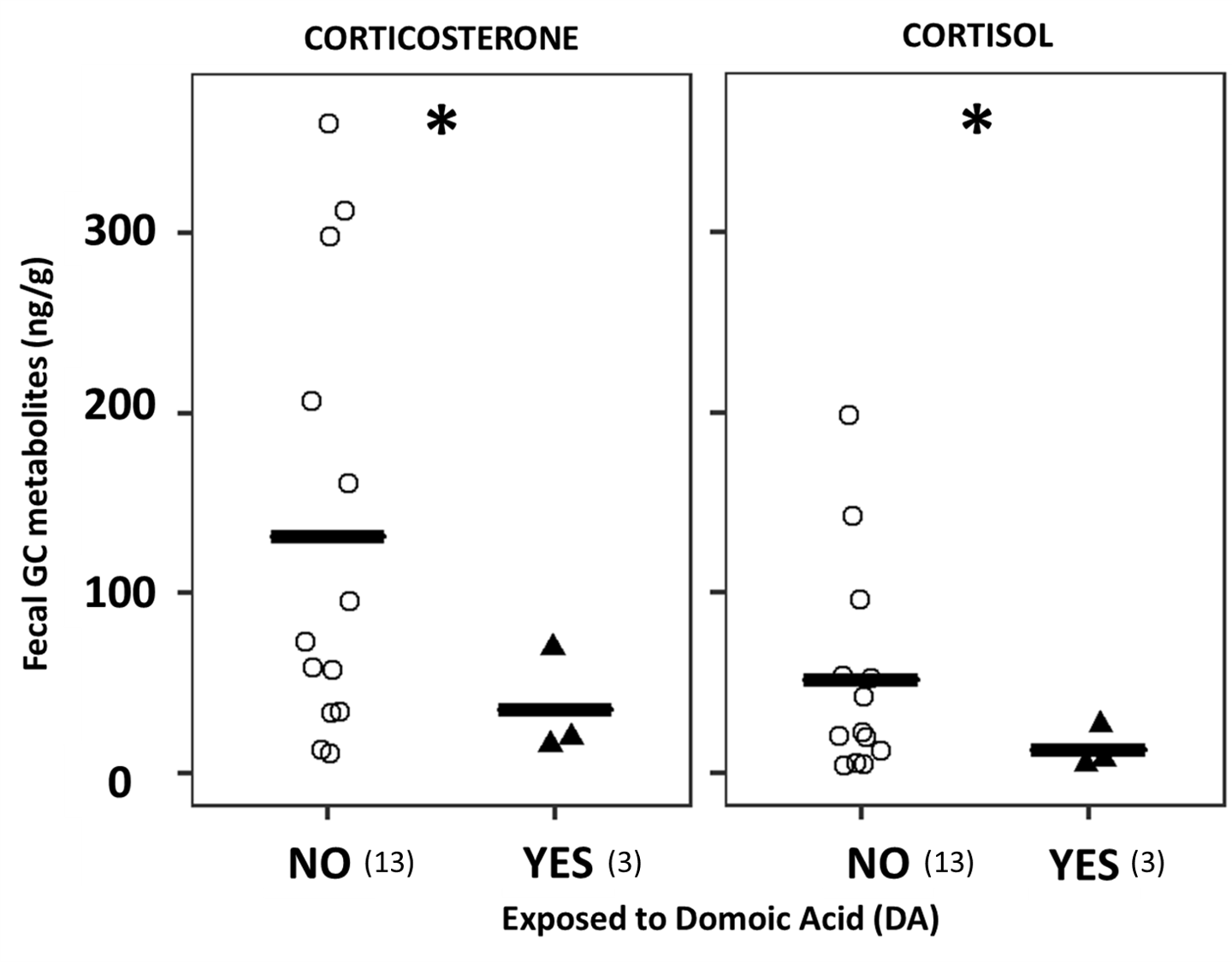
In our recent scientific publication (D’Agostino et al. 2021), we investigate SRW exposure to DA at their breeding ground in Peninsula Valdes and assessed its effects on fecal glucocorticoid concentrations. Although the sample size of this study is unavoidably small due to the difficulties of obtaining fecal samples from whales at their calving grounds where defecation is infrequent, we observed significantly lower fGCm in samples from whales exposed to DA (Figure 4). Our results agree with findings from a previous study in California sea lions (Zalophus californianus) exposed to DA, where these authors found a significant association of DA exposure with reduced serum cortisol (Gulland et al., 2009), which can be tentatively attributed to abnormal function of the adrenal axis due to the exposure.
Figure 4. Fecal glucocorticoid metabolite levels in southern right whales exposed (YES, solid triangles) and not-exposed (NO, open circles) to DA. Left panel: immunoreactive fecal corticosterone metabolites. Right panel: immunoreactive fecal cortisol metabolites. Hormone concentrations are expressed in ng of immunoreactive hormone per gram of dry fecal sample. Significant differences between groups are denoted with an asterisk (P<0.05). The black solid line indicates the mean for each group, and in parenthesis is the sample size for each group. Adapted from D’Agostino et al. 2021.
If ingestion of toxins produced by phytoplankton can result in long-term suppression of baseline GCs, whales and marine mammals in general, could suffer reduced ability to cope with additional stressors. The adrenal function is essential to maintain circulating blood glucose and other aspects of metabolism within normal bounds. Additionally, the ability to elevate GCs facilitates energy mobilization to physiologically cope with a stressful event and to initiate appropriate behavioral responses (i.e., flee from predators, heal wounds). Various toxicants have been shown to reduce adrenal function across taxa (Romero and Wingfield, 2016) and could have negative consequences on the ability of cetaceans to respond and adapt to ongoing environmental and anthropogenic changes. Compounding this problem, whales are exposed to an increasing number of stressors from multiple sources and with cumulative effects and they need to be able to physiologically respond to continue to reproduce and survive.
To our knowledge, this study provides the first quantification of fGCm levels in whales exposed to DA; and we hope this effort starts a growing dataset to which other researchers can add. Sampling and analysis of non-traditional matrices, such as feces, blubber, baleen and others, would likely increase sample sizes and thus our understanding of the interrelationships among DA exposure and age, sex, and reproductive status of cetaceans. Given that chronic exposure to DA could alter the capacity of animals to respond to stress, and indications that HABs are becoming more frequent and intense world-wide (Van Dolah 2000; Masó et al. 2006; Erdner et al. 2008), we believe that research evaluating the health status of marine mammal populations should include the assessment of stress physiology relative to natural and anthropogenic stressors including exposure to toxicants.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get weekly updates and more! Just add your name into the subscribe box on the left panel.
References
Bastida R, Rodríguez D (2009) Ballena franca austral [Southern right whale]. In: Mazzini V (ed) Mamíferos marinos de Patagonia y Antártida [Marine mammals of Patagonia and Antarctica]. Zagier & Urruty Publications, Buenos Aires, pp 72–84.
Bonier F, Moore IT, Martin PR, Robertson RJ (2009) The relationship between fitness and baseline glucocorticoids in a passerine bird. Gen Comp Endocrinol 163:208–213. https:// doi. org/ 10. 1016/j.ygcen. 2008. 12. 013.
Brann DW, Mahesh VB (1994) Excitatory amino acids: function and significance in reproduction and neuroendocrine regulation. Front Neuroendocrinol 15:3–49. https:// doi. org/ 10. 1006/ frne. 1994. 1002.
Broadwater MH, Van Dolah FM, Fire SE (2018) Vulnerabilities of marine mammals to harmful algal blooms. Harmful Algal Blooms 2:191–222.
Burgess, E.A., Hunt, K.E., Kraus, S.D., Rolland, R.M., 2016. Get the most out of blow hormones: Validation of sampling materials, field storage and extraction techniques for whale respiratory vapour samples. Conserv. Physiol. 4, cow024. https://doi.org/10.1093/conphys/cow024
Burgess, E.A., Hunt, K.E., Kraus, S.D., Rolland, R.M., 2018. Quantifying hormones in exhaled breath for physiological assessment of large whales at sea. Sci. Rep. 8, 10031. https://doi.org/10.1038/s41598-018-28200-8
D’Agostino VC, Hoffmeyer MS, Almandoz GO, Sastre V, Degrati M., 2015. Potentially toxic Pseudo-nitzschia species in plankton and fecal samples of Eubalaena australis from Península Valdés calv-ing ground, Argentina. J Sea Res 106:39–43. https:// doi. org/ 10.1016/j. seares.
D’Agostino, V.C., Degrati, M., Santinelli, N., Sastre, V., Dans, S.L., Hoffmeyer, M.S., 2018. The seasonal dynamics of plankton communities relative to the foraging of the southern right whale (Eubalaena australis) in northern Patagonian gulfs, Península Valdés, Argentina. Cont. Shelf Res. 164, 45–57. https://doi.org/10.1016/j.csr.2018.06.003
D’Agostino, V.C., Degrati, M., Sastre, V., Santinelli, N., Krock, B., Krohn, T., Dans, S.L., Hoffmeyer, M.S., 2017. Domoic acid in a marine pelagic food web: Exposure of southern right whales Eubalaena australis to domoic acid on the Península Valdés calving ground, Argentina. Harmful Algae 68, 248–257. https://doi.org/10.1016/j.hal.2017.09.001
D’Agostino, V.C., Hoffmeyer, M.S., Degrati, M., 2016. Faecal analysis of southern right whales (Eubalaena australis) in Península Valdés calving ground, Argentina: Calanus australis, a key prey species. J. Mar. Biol. Assoc. United Kingdom 96, 859–868. https://doi.org/10.1017/S0025315415001897
Dickens, M.J., Romero, L.M., 2013. A consensus endocrine profile for chronically stressed wild animals does not exist. Gen. Comp. Endocrinol. 191, 177–189. https://doi.org/10.1016/j.ygcen.2013.06.014
Erdner DL, Dyble J, Parsons ML, Stevens RC, Hubbard KA, Wrabel ML, Moore SK, Lefebvre KA, Anderson DM, Bienfang P, Bidi-gare RR, Parker MS, Moeller P, Brand LE, Trainer VL (2008) Centers for Oceans and Human Health: a unified approach to the challenge of harmful algal blooms. Environ Health 7:S2. https://doi. org/ 10. 1186/ 1476- 069X-7- S2- S2.
Fernández Ajó, A., Hunt, K.E., Dillon, D., Uhart, M., Sironi, M., Rowntree, V., Buck, C.L., 2021. Optimizing hormone extraction protocols for whale baleen: tackling questions of solvent:sample ratio and variation. Gen. Comp. Endocrinol. 113828. https://doi.org/10.1016/j.ygcen.2021.113828
Fernández Ajó, A.A., Hunt, K.E., Giese, A.C., Sironi, M., Uhart, M., Rowntree, V.J., Marón, C.F., Dillon, D., DiMartino, M., Buck, C.L., 2020. Retrospective analysis of the lifetime endocrine response of southern right whale calves to gull wounding and harassment: A baleen hormone approach. Gen. Comp. Endocrinol. 296, 113536. https://doi.org/10.1016/j.ygcen.2020.113536
Fernández Ajó, A.A., Hunt, K.E., Uhart, M., Rowntree, V., Sironi, M., Marón, C.F., Di Martino, M., Buck, C.L., 2018. Lifetime glucocorticoid profiles in baleen of right whale calves: potential relationships to chronic stress of repeated wounding by Kelp Gulls. Conserv. Physiol. 6, 1–12. https://doi.org/10.1093/conphys/coy045
Fire SE, Bogomolni A, DiGiovanni RA Jr, Early G, Leighfield TA, Matassa K, Miller GA, Moore KM, Moore M, Niemeyer M, Pugliares K (2021) An assessment of temporal, spatial and taxonomic trends in harmful algal toxin exposure in stranded marine mammals from the US New England coast. PLoS ONE 16(1):e0243570. https:// doi. org/ 10. 1371/ journ al. pone. 02435 70
Fire SE, Wang Z, Berman M, Langlois GW, Morton SL, Sekula-Wood E, Benitez-Nelson CR (2010) Trophic transfer of the harmful algal toxin domoic acid as a cause of death in a minke whale (Balaenoptera acutorostrata) stranding in southern California. Aquat Mamm 36(4):342–350. https:// doi. org/ 10. 1578/ AM. 36.4.2010. 342.
Gulland F.M., 1999. Domoic acid toxicity in California sea lions stranded along the central California Coast, May-October 1998. NOAA Tech. Memo. NMFS-OPR-8. USA National Marine Fisheries Service, US Department of Commerce.
Gulland FMD, Hall AJ, Greig DJ, Fram ER, Colegrove KM, Booth RKN, Wasser SK, S.-M.C., 2009. Gulland, Hall – 2012 – Evaluation of circulating eosinophil count and adrenal gland function in California sea lions naturally exposed t. J. Am. Vet. Med. Assoc. 241, 943–949.
Gulland, F., Pérez-Cortés, H., Urbán, J.R., Rojas-Bracho, L., Ylitalo, G., Weir, J., Norman, S., Muto, M., Rugh, D., Kreuder, C., Rowles, T., 2005. Eastern North Pacific gray whale (Eschrichtius robustus) unusual mortality event, 1999-2000. U.S. Dep. Commer. NOAA Tech. Memo. NMFS-AFSC-150. 33 pp.
Hunt, K.E., Robbins, J., Buck, C.L., Bérubé, M., Rolland, R.M., 2019. Evaluation of fecal hormones for noninvasive research on reproduction and stress in humpback whales (Megaptera novaeangliae). Gen. Comp. Endocrinol. 280, 24–34. https://doi.org/10.1016/j.ygcen.2019.04.004
Hunt, K.E., Rolland, R.M., Kraus, S.D., Wasser, S.K., 2006. Analysis of fecal glucocorticoids in the North Atlantic right whale (Eubalaena glacialis). Gen. Comp. Endocrinol. 148, 260–272. https://doi.org/10.1016/j.ygcen.2006.03.012
Hunt, K.E., Stimmelmayr, R., George, C., Hanns, C., Suydam, R., Brower, H., Rolland, R.M., 2014. Baleen hormones: a novel tool for retrospective assessment of stress and reproduction in bowhead whales (Balaena mysticetus). Conserv. Physiol. 2, cou030–cou030. https://doi.org/10.1093/conphys/cou030
Johnson MP, Kelly G, Chamberlain M (2001) Changes in rat serum corticosterone after treatment with metabotropic glutamate receptor agonists or antagonists. J Neuroendocrinol 13:670–677. https:// doi. org/ 10. 1046/j. 1365- 2826. 2001. 00678.x.
Lefebvre KA, Powell CL, Busman M, Doucette GJ, Moeller PDR, Sliver JB, Miller PE, Hughes MP, Singaram S, Silver MW, Tjeer-dema RS (1999) Detection of domoic acid in northern anchovies and California sea lions associated with an unusual mortality event. Nat Toxins 7(3):85–92. https:// doi. org/ 10. 1002/ (SICI) 1522-7189(199905/ 06)7: 3% 3C85:: AID- NT39% 3E3.0. CO;2-Q.
Lefebvre, K.A., Kendrick, P.S., Ladiges, W., Hiolski, E.M., Ferriss, B.E., Smith, D.R., Marcinek, D.J., 2017. Chronic low-level exposure to the common seafood toxin domoic acid causes cognitive deficits in mice. Harmful Algae 64, 20–29. https://doi.org/10.1016/j.hal.2017.03.003
Lemos, L.S., Olsen, A., Smith, A., Burnett, J.D., Chandler, T.E., Larson, S., Hunt, K.E., Torres, L.G., 2021. Stressed and slim or relaxed and chubby? A simultaneous assessment of gray whale body condition and hormone variability. Mar. Mammal Sci. 1–11. https://doi.org/10.1111/mms.12877
Lemos, L.S., Olsen, A., Smith, A., Chandler, T.E., Larson, S., Hunt, K., Torres, L.G., 2020. Assessment of fecal steroid and thyroid hormone metabolites in eastern North Pacific gray whales. Conserv. Physiol. 8. https://doi.org/10.1093/conphys/coaa110
Masó M, Garcés E., 2006. Harmful microalgae blooms (HAB); prob-lematic and conditions that induce them. Mar Pollut Bull 53:620–630. https:// doi. org/ 10. 1016/j. marpo lbul. 2006. 08. 006.
Pulido O.M., 2008. Domoic acid toxicologic pathology: a review. Mar Drugs 6(2):180–219. https:// doi. org/ 10. 3390/ md602 0180.
Rolland, R., McLellan, W., Moore, M., Harms, C., Burgess, E., Hunt, K., 2017. Fecal glucocorticoids and anthropogenic injury and mortality in North Atlantic right whales Eubalaena glacialis. Endanger. Species Res. 34, 417–429. https://doi.org/10.3354/esr00866
Rolland, R.M., Hunt, K.E., Kraus, S.D., Wasser, S.K., 2005. Assessing reproductive status of right whales (Eubalaena glacialis) using fecal hormone metabolites. Gen. Comp. Endocrinol. 142, 308–317. https://doi.org/10.1016/j.ygcen.2005.02.002
Romero, M.L., Wingfield, J.C., 2016. Oxford series in behavioral neuroendocrinology. Tempests, poxes, predators, and people: stress in wild animals and how they cope 1–2.
Sastre V, Santinelli N, Marino G, Solís M, Pujato L, Ferrario M., 2007. First detection of domoic acid produced by Pseudo-nitzschia spe-cies, Chubut coastal waters, Patagonia, Argentina. Harmful Algae News 34:12–14.
Van Dolah FM., 2000. Marine algal toxins: origins, health effects, and their increased occurrence. Environ Health Perspect 108:133–141. https:// doi. org/ 10. 1289/ ehp. 00108 s1133
By: Alexa Kownacki, Ph.D. Student, OSU Department of Fisheries and Wildlife, Geospatial Ecology of Marine Megafauna Lab
Humans are fascinated by food. We want to know its source, its nutrient content, when it was harvested and by whom, and so much more. Since childhood, I was the nagging child who interrogated wait staff about the seafood menu because I cared about the sustainability aspect as well as consuming ethically-sourced seafood. Decades later I still do the same: ask a myriad of questions from restaurants and stores in order to eat as sustainably as possible. But in addition to asking these questions about my food, I also question what my study species eats and why. My study populations, common bottlenose dolphins, are described as top opportunistic predators (Norris and Prescott 1961, Shane et al. 1986, Barros and Odell 1990). In my study area off of California, this species exists in two ecotypes. The coastal ecotype off of California, USA are generalist predators, feeding on many different species of fish using different foraging techniques (Ballance 1992, Shane 1990). The offshore ecotype, on the other hand, is less well-studied, but is frequently observed in association with sperm whales, although the reason is still unknown (Díaz-Gamboa et al. 2018). Stable isotope analysis from skin samples from the two ecotypes indicates that the ecotypes exhibit different foraging strategies based on different isotopic carbon and nitrogen levels (Díaz-Gamboa et al. 2018).
Growing up, I kept the Monterey Bay Aquarium’s Seafood Watch Guide with me to choose the most sustainably-sourced seafood at restaurants. Today there is an easy-to-use application for mobile phones that replaced the paper guide. (Image Source: https://www.seafoodwatch.org/)
Preliminary and historical data on common bottlenose dolphins (Tursiops truncatus) suggest that the coastal ecotype spend more time near estuary mouths than offshore dolphins (Ballance 1992, Kownacki et al. unpublished data). Estuaries contain large concentrations of nutrients from runoff, which support zooplankton and fishes. It is for this reason that these estuaries are thought to be hotspots for bottlenose dolphin foraging. Some scientists hypothesize that these dolphins are estuarine-based prey specialists (Barros and Odell 1990), or that the dolphins simply aggregate in estuaries due to higher prey abundance (Ballance 1992).
Coastal bottlenose dolphins traveling near an estuary mouth in San Diego, CA. (Photographed under NOAA NMFS Permit # 19091).
In an effort to understand diet compositions of bottlenose dolphins, during coastal surveys seabirds were recorded in association with feeding groups of dolphins. Therefore, it is reasonable to believe that dolphins were feeding on the same fishes as Brown pelicans, blue-footed and brown boobies, double-crested cormorants, and magnificent frigatebirds, seeing as they were the most common species associated with bottlenose dolphin feeding groups (Ballance 1992). A shore-based study by Hanson and Defran (1993) found that coastal dolphins fed more often in the early morning and late afternoon, as well as during periods of high tide current. These patterns may have to do with the temporal and spatial distribution of prey fish species. From the few diet studies conducted on these bottlenose dolphins in this area, 75% of the prey were species from the families Ebiotocidae (surf perches) and Sciaendae (croakers) (Norris and Prescott 1961, Walker 1981). These studies, in addition to optimal foraging models, suggest this coastal ecotype may not be as much of a generalist as originally suggested (Defran et al. 1999).
A redtail surfperch caught by a fisherman from a beach in San Diego, CA. These fish are thought to be common prey of coastal bottlenose dolphins. (Image Source: FishwithJD)
Diet studies on the offshore ecotype of bottlenose dolphins worldwide show a preference for cephalopods, similar to other toothed cetaceans who occupy similar regions, such as Risso’s dolphin, sperm whales, and pilot whales (Clarke 1986, Cockcroft and Ross 1990, Gonzalez et al. 1994, Barros et al. 2000, Walker et al. 1999). Because these animals seldom strand on accessible beaches, stomach contents analyses are limited to few studies and isotope analysis is more widely available from biopsies. We know these dolphins are sighted in deeper waters than the habitat of coastal dolphins where there are fewer nutrient plumes, so it is reasonable to hypothesize that the offshore ecotype consumes different species and may be more specialized than the coastal ecotype.
An bottlenose dolphin forages on an octopus. (Image source: Mandurah Cruises)
For a species that is so often observed from shore and boats, and is known for its charisma, it may be surprising that the diets of both the coastal and offshore bottlenose dolphins are still largely unknown. Such is the challenge of studying animals that live and feed underwater. I wish I could simply ask a dolphin, much like I would ask staff at restaurants: what is on the menu today? But, unfortunately, that is not possible. Instead, we must make educated hypotheses about the diets of both ecotypes based on necropsies and stable isotope studies, and behavioral and spatial surveys. And, I will continue to look to new technologies and creative thinking to provide the answers we are seeking.
Literature cited:
Ballance, L. T. (1992). Habitat use patterns and ranges of the bottlenose dolphin in the Gulf of California, Mexico. Marine Mammal Science, 8(3), 262-274.
Barros, N.B., and D. K. Odell. (1990). Food habits of bottlenose dolphins in the southeastern United States. Pages 309-328 in S. Leatherwood and R. R. Reeves, eds. The bottlenose dolphin. Academic Press, San Diego, CA.
Barros, N., E. Parsons and T. Jefferson. (2000). Prey of bottlenose dolphins from the South China Sea. Aquatic Mammals 26:2–6.
Clarke, M. 1986. Cephalopods in the diet of odontocetes. Pages 281–321 in M. Bryden and R. Harrison, eds. Research on dolphins. Clarendon Press, Oxford, NY.
Cockcroft, V., and G. Ross. (1990). Food and feeding of the Indian Ocean bottlenose dolphin off southern Natal, South Africa. Pages 295–308 in S. Leatherwood and R. R. Reeves, eds. The bottlenose dolphin. Academic Press, San Diego, CA.
Defran, R. H., Weller, D. W., Kelly, D. L., & Espinosa, M. A. (1999). Range characteristics of Pacific coast bottlenose dolphins (Tursiops truncatus) in the Southern California Bight. Marine Mammal Science, 15(2), 381-393.
Díaz‐Gamboa, R. E., Gendron, D., & Busquets‐Vass, G. (2018). Isotopic niche width differentiation between common bottlenose dolphin ecotypes and sperm whales in the Gulf of California. Marine Mammal Science, 34(2), 440-457.
Gonzalez, A., A. Lopez, A. Guerra and A. Barreiro. (1994). Diets of marine mammals stranded on the northwestern Spanish Atlantic coast with special reference to Cephalopoda. Fisheries Research 21:179–191.
Hanson, M. T., and Defran, R. H. (1993). The behavior and feeding ecology of the Pacific coast bottlenose dolphin, Tursiops truncatus. Aquatic Mammals, 19, 127-127.
Norris, K. S., and J. H. Prescott. (1961). Observations on Pacific cetaceans of Californian and Mexican waters. University of California Publications of Zoology 63:29, 1-402.
Shane, S. H. (1990). Comparison of bottlenose dolphin behavior in Texas and Florida, with a critique of methods for studying dolphin behavior. Pages 541-558 in S. Leatherwood and R. R. Reeves, eds. The bottlenose dolphin. Academic Press, San Diego, CA.
Shane, S., R. Wells and B. Wursig. (1986). Ecology, behavior and social organization of bottlenose dolphin: A review. Marine Mammal Science 2:34–63.
Walker, W.A. (1981). Geographical variation in morphology and biology of the bottlenose dolphins (Tursiops) in the eastern North Pacific. NMFS/SWFC Administrative Report. No, LJ-91-03C.
Walker, J., C. Potter and S. Macko. (1999). The diets of modern and historic bottlenose dolphin populations reflected through stable isotopes. Marine Mammal Science 15:335–350.
Another year has come and gone, and with the final days of 2019 upon us, it is fulfilling to look back and summarize all of the achievements in the GEMM Lab this year. So, snuggle up with your favorite holiday drink and enjoy our recap of 2019!
We wrapped up two intense but rewarding gray whale field seasons this summer. Our project investigating the health of Pacific Coast Feeding Group (PCFG) gray whales through fecal hormone and body condition sampling in the context of ocean noise went into its fourth year, while the Port Orford project where we track whales and prey at a very fine-scale celebrated its wood anniversary (five years!). The dedication and hard work of lots of people to help us collect our data meant that we were able to add a considerable amount of samples to our growing gray whale datasets. Our trusty red RHIB Ruby zipped around the Pacific and enabled us to collect 58 fecal samples, fly the drone 102 times, undertake 105 GoPro drops and record 141 gray whale sightings. Our Newport crew was a mix of full-time GEMMers (Leigh, Todd, Dawn, Leila, Clara, and myself) as well as part-time summer GEMMers (Ale, Sharon, and Cassy). Further south in Port Orford, my team of undergraduate and high school students and I had an interesting field season. We only encountered four different individuals (Buttons, Glacier, Smudge, and Primavera), however saw them repeatedly throughout the month of August, resulting in as many as 15 tracklines for one individual. Furthermore, we collected 249 GoPro drops and 248 zooplankton net samples.
Leila taking photos of gray whales from Ruby’s bow pulpit. Photo: Leigh Torres
2019 Port Orford team members Anthony & Lisa collecting prey samples from research kayak ‘Robustus’.
Gray whale fluke. Photo: Lisa Hildebrand.
The GEMM Lab’s fieldwork was not just restricted to gray whales. After last year’s successes aboard the NOAA ship Bell M. Shimada, Alexa and Dawn both boarded the ship again this year as marine mammal observers for the May and September cruises, respectively. They spied humpback, blue, sperm, and fin whales, as well as many dolphins and seabirds, adding to the GEMM Lab’s growing database of megafauna distribution off the Oregon coast.
Alexa observing on the R/V Shimada in May 2019, all bundled up. Image Photo: Alexa Kownacki
Dawn Barlow on the flying bridge of NOAA Ship Bell M. Shimada, heading out to sea with the Newport bridge in the background. Photo: Anna Bolm.
After winning the prestigious L’Oréal-UNESCO For Women in Science fellowship and the inaugural Louis Herman Scholarship, GEMM Lab grad Solène Derville lead her first research cruise aboard the French R/V Alis. She and her team conducted line transect surveys and micronekton/oceanographic sampling over several seamounts to try to solve the mystery of why humpbacks hang out there. We are also very excited to announce that Solène will be returning to the GEMM Lab as a post-doc in 2020! She will be creating distribution models of whales off the coast of Oregon with the data collected by Leigh during helicopter flights with the US Coast Guard. The primary aim of this work is to identify potential whale hotspots in an effort to avoid spatial overlap with fisheries gear and reduce entanglement risk.
Solène soaking wet after spending several hours observing cetaceans and seabirds on R/V Alis. Photo: Jérôme Jambou
A group of bottlenose dolphins observed over one of the seamounts. Photo: Elodie Vourey
Solène at the L’Oréal ceremony in the French National Museum of Natural History in Paris. Photo: Jean-Charles Caslot
Switching the focus from marine mammals to seabirds, Rachael has had an extremely busy year of field work all across the globe. She island-hopped from Midway (Hawaiian Northwest island) to the Falkland Islands in the first half of the year, and is currently overwintering on South Georgia, where she will be until end of February. Rachael is tracking albatross at all three locations by tagging individual birds to understand movements relative to fishing vessels and flight energetics.
Albatross chick. Photo: Rachael Orben
Recording data. Photo: V. Ternisien
Albatross chick and mother. Photo: Rachael Orben.
Besides several field efforts, the GEMM Lab was also busy disseminating our research and findings to various audiences. Our conferences kicked off in late February when Leigh and Rachael both flew to Kauai to present at the Pacific Seabird Group’s 46th Annual Meeting. In the spring, Leila, Dawn, Alexa, Dom, and myself drove to Seattle where the University of Washington hosted the Northwest Student Society of Marine Mammalogy chapter meeting and we all gave talks. Additionally, the Fisheries & Wildlife grad students in the lab also presented at the department’s annual Research Advances in Fisheries, Wildlife, and Ecology conference. Later in the year, Dom and I attended the State of the Coast conference where Dom was invited to participate in a panel about the holistic approaches to management in the nearshore while I presented a poster on preliminary findings of my Master’s thesis. Most recently, the entire GEMM Lab (bar Rachael) flew to Barcelona to present at the World Marine Mammal Conference (WMMC).
GEMM Lab at the WMMC. Photo: Karen Lohman
Our science communication and outreach efforts were not just restricted to conferences though. Over the course of this year, the GEMM Lab supervised a total of 10 undergraduate and high school interns that assisted in a variety of ways (field and/or lab work, data analyses, independent projects) on a number of projects going on in the lab. Leigh and Dawn boarded the R/V Oceanus in the fall to co-lead a STEM research cruise aimed at providing high school students and teachers hands-on marine research. Dawn and I were guests on Inspiration Dissemination, a live radio show run by graduate students about graduate research going on at OSU. Our weekly blog, now in its fifth year, reached its highest viewership with a total of 14,814 views this year!
The GEMMers were once again prolific writers too! The 13 new publications in 10 scientific journals include contributions from Leigh (7), Rachael (6), Solène (2), Dawn (2), and Leila (1). Scroll down to the end of the post to see the list.
Academic milestones were also reached by several of us. Most notably and recently, Dom successfully defended his Master’s thesis this past week – congratulations Dom!! Unsurprisingly, he already has a job lined up starting in January as a Science Officer with the California Ocean Science Trust. Dom is the 6th GEMM Lab graduate, which after just five years of the GEMM Lab existing is a huge testament to Leigh as an advisor. Leila, who is in the 4th year of her PhD, anticipates finishing this coming March. We also had three successful research reviews – I met with my committee in late March to discuss my Master’s proposal, while Alexa and Dawn met with their committees in the summer to review their PhD proposals. All three reviews were fruitful and successful. And we want to highlight the success of a GEMM Lab grad, Florence Sullivan, who started a job in Maui with the Pacific Whale Foundation in September as a Research Analyst.
Dom during his MS seminar. Photo: Leila Lemos
Post-defense happiness. Photo: Karen Lohman
Leigh was recognized for her expertise in gray whale ecology and was appointed to the IUCN Western Gray Whale Advisory Panel (WGWAP). The western gray whales are a critically endangered population. At one point in the 1960s, the population was so scarce that they were believed to have been extinct. While this concern did not prove to be the case, the population still is not doing well, which is why the IUCN formed WGWAP to provide advice on the conservation of the western gray whales. Leigh was appointed to the panel this year and traveled to Switzerland and Russia for meetings.
Clara aboard Ruby on her first day of gray whale field work in Oregon. Photo: Leigh Torres
We are excited about a new addition to the lab. Clara Bird started her MS in Wildlife Science in the Department of Fisheries & Wildlife this fall. She jumped straight into field work when she came in early September and got a taste of the Pacific. Clara joins us from the Duke University where she did her undergraduate degree and worked for the past year in their Marine Robotics and Remote Sensing Lab. Clara is digging into the gray whale drone footage collected over the last four field seasons and scrutinize them from a behavioral point of view.
If you are reading this post, we would like to say that we really appreciate your support and interest in our work! We hope you will continue to join us on our journeys in 2020. Until then, happy holidays from the GEMM Lab!
GEMM Lab at the beginning of June with some permanents GEMMs and some temporary summer GEMM helpers.
Barlow, D. R., M. Fournet, and F. Sharpe. 2019. Incorporating tides into the acoustic ecology of humpback whales. Marine Mammal Science 35:234-251.
Barlow, D. R., A. L. Pepper, and L. G. Torres. 2019. Skin deep: an assessment of New Zealand blue whale skin condition. Frontiers in Marine Science doi.org/10.3389/fmars.2019.00757.
Baylis, A. M. M., R. A. Orben, A. A. Arkhipkin, J. Barton, R. L. Brownell Jr., I. J. Staniland, and P. Brickle. 2019. Re-evaluating the population size of South American fur seals and conservation implications. Aquatic Conservation: Marine and Freshwater Ecosystems 29(11):1988-1995.
Baylis, A. M. M., M. Tierney, R. A. Orben, et al. 2019. Important at-sea areas of colonial breeding marine predators on the southern Patagonian Shelf. Scientific Reports 9:8517.
Cockerham, S., B. Lee, R. A. Orben, R. M. Suryan, L. G. Torres, P. Warzybok, R. Bradley, J. Jahncke, H. S. Young, C. Ouverney, and S. A. Shaffer. 2019. Microbial biology of the western gull (Larus occidentalis). Microbial Ecology 78:665-676.
Derville, S., L. G. Torres, R. Albertson, O. Andrews, C. S. Baker, P. Carzon, R. Constantine, M. Donoghue, C. Dutheil, A. Gannier, M. Oremus, M. M. Poole, J. Robbins, and C. Garrigue. 2019. Whales in warming water: assessing breeding habitat diversity and adaptability in Oceania’s changing climate. Global Change Biology 25(4):1466-1481.
Derville, S., L. G. Torres, R. Dodémont, V. Perard, and C. Garrigue. 2019. From land and sea, long-term data reveal persistent humpback whale (Megaptera novaeangliae) breeding habitat in New Caledonia. Aquatic Conservation: Marine and Freshwater Ecosystems 29(10):1697-1711.
Fleischman, A. B., R. A. Orben, N. Kokubun, A. Will, R. Paredes, J. T. Ackerman, A. Takahashi, A. S. Kitaysky, and S. A. Shaffer. 2019. Wintering in the western Subantarctic Pacific increases mercury contamination of red-legged kittiwakes. Environmental Science & Technology 53(22):13398-13407.
Holdman, A. K., J. H. Haxel, H. Klinck, and L. G. Torres. 2019. Acoustic monitoring reveals the times and tides of harbor porpoise (Phocoena phocoena) distribution off central Oregon, U.S.A. Marine Mammal Science 35:164-186.
Kroeger, C., D. E. Crocker, D. R. Thompson, L. G. Torres, P. Sagar, and S. A. Shaffer. 2019. Variation in corticosterone levels in two species of breeding albatrosses with divergent life histories: responses to body condition and drivers of foraging behavior. Physiological and Biochemical Zoology 92(2):223:238.
Loredo, S. A., R. A. Orben, R. M. Suryan, D. E. Lyons, J. Adams, and S. W. Stephensen. 2019. Spatial and temporal diving behavior of non-breeding common murres during two summers of contrasting ocean conditions. Journal of Experimental Biology and Ecology 517:13-24.
Monteiro, F., L. S. Lemos, J. Fulgêncio de Moura, R. C. C. Rocha, I. Moreira, A. P. Di Beneditto, H. A. Kehrig, I. C. A. C. Bordon, S. Siciliano, T. D. Saint’Pierre, and R. A. Hauser-Davis. 2019. Subcellular metal distributions and metallothionein associations in rough-toothed dolphins (Steno bredanensis) from southeastern Brazil. Marine Pollution Bulletin 146:263-273.
Orben, R. A., A. B. Fleischman, A. L. Borker, W. Bridgeland, A. J. Gladics, J. Porquez, P. Sanzenbacher, S. W. Stephensen, R. Swift, M. W. McKown, and R. M. Suryan. 2019. Comparing imaging, acoustics, and radar to monitor Leach’s storm-petrel colonies. PeerJ 7:e6721.
Yates, K. L., …, L. G. Torres, et al. 2019. Outstanding challenges in the transferability of ecological models. Trends in Ecology & Evolution 33(10):790-802.
By Clara Bird, Masters Student, OSU Department of Fisheries and Wildlife, Geospatial Ecology of Marine Megafauna Lab
Hi everyone! My name is Clara Bird and I am
the newest graduate student in the GEMM lab. For my master’s thesis I will be
using drone footage of gray whales to study their foraging ecology. I promise
to talk about how cool gray whales in a following blog post, but for my first effort
I am choosing to write about something that I have wanted to explain for a
while: algorithms. As part of previous research projects, I developed a few semi-automated
image analysis algorithms and I have always struggled with that jargon-filled
phrase. I remember being intimidated by the term algorithm and thinking that I
would never be able to develop one. So, for my first blog I thought that I
would break down what goes into image analysis algorithms and demystify a term
that is often thrown around but not well explained.
What is an algorithm?
The dictionary broadly defines an
algorithm as “a step-by-step procedure for solving a problem or accomplishing
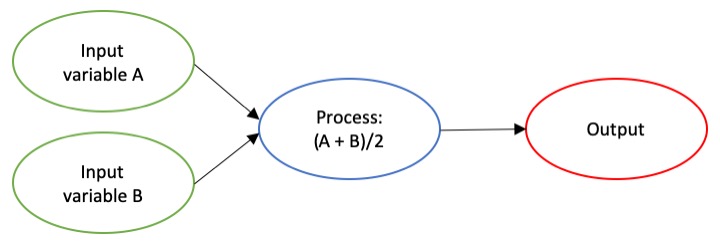
some end” (Merriam-Webster). Imagine an algorithm as a flow chart (Fig. 1), where
each step is some process that is applied to the input(s) to get the desired
output. In image analysis the output is usually isolated sections of the image
that represent a specific feature; for example, isolating and counting the
number of penguins in an image. Algorithm development involves figuring out
which processes to use in order to consistently get desired results. I have
conducted image analysis previously and these processes typically involve figuring
out how to find a certain cutoff value. But, before I go too far down that
road, let’s break down an image and the characteristics that are important for
image analysis.
Figure 1. An example of a basic algorithm flow chart. There are two inputs: variables A and B. The process is the calculation of the mean of the two variables.
What is an image?
Think of an image as a spread sheet,
where each cell is a pixel and each pixel is assigned a value (Fig. 2). Each
value is associated with a color and when the sheet is zoomed out and viewed as
a whole, the image comes together. In
color imagery, which is also referred to as RGB, each pixel is associated with
the values of the three color bands (red, green, and blue) that make up that
color. In a thermal image, each pixel’s value is a temperature value. Thinking
about an image as a grid of values is helpful to understand the challenge of
translating the larger patterns we see into something the computer can interpret.
In image analysis this process can involve using the values of the pixels
themselves or the relationships between the values of neighboring pixels.
Our brains take in the whole
picture at once and we are good at identifying the objects and patterns in an
image. Take Figure 3 for example: an astute human eye and brain can isolate and
identify all the different markings and scars on the fluke. Yet, this process
would be very time consuming. The trick to building an algorithm to conduct
this work is figuring out what processes or tools are needed to get a computer
to recognize what is marking and what is not. This iterative process is the algorithm
development.
Figure 3. Photo ID image of a gray whale fluke.
Development
An image analysis algorithm will
typically involve some sort of thresholding. Thresholds are used to classify an
image into groups of pixels that represent different characteristics. A
threshold could be applied to the image in Figure 3 to separate the white color
of the markings on the fluke from the darker colors in the rest of the image.
However, this is an oversimplification, because while it would be pretty simple
to examine the pixel values of this image and pick a threshold by hand, this threshold
would not be applicable to other images. If a whale in another image is a
lighter color or the image is brighter, the pixel values would be different
enough from those in the previous image for the threshold to inaccurately
classify the image. This problem is why a lot of image analysis algorithm
development involves creating parameterized processes that can calculate the
appropriate threshold for each image.
One successful method used to
determine thresholds in images is to first calculate the frequency of color in
each image, and then apply the appropriate threshold. Fletcher et al. (2009)
developed a semiautomated algorithm to detect scars in seagrass beds from
aerial imagery by applying an equation to a histogram of the values in each
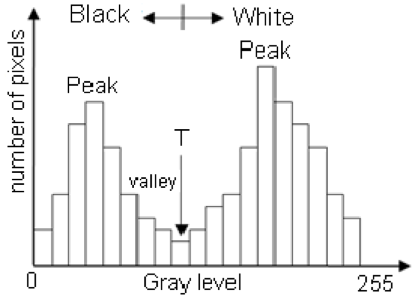
image to calculate the threshold. A histogram is a plot of the frequency of
values binned into groups (Fig. 4). Essentially, it shows how many times each value
appears in an image. This information can be used to define breaks between
groups of values. If the image of the fluke were transformed to a gray scale, then
the values of the marking pixels would be grouped around the value for white
and the other pixels would group closer to black, similar to what is shown in
Figure 4. An equation can be written that takes this frequency information and
calculates where the break is between the groups. Since this method calculates
an individualized threshold for each image, it’s a more reliable method for
image analysis. Other characteristics could also be used to further filter the
image, such as shape or area.
However, that approach is not the
only way to make an algorithm applicable to different images; semi-automation
can also be helpful. Semi-automation involves some kind of user input. After
uploading the image for analysis, the user could also provide the threshold, or
the user could crop the image so that only the important components were maintained.
Keeping with the fluke example, the user could crop the image so that it was
only of the fluke. This would help reduce the variety of colors in the image
and make it easier to distinguish between dark whale and light marking.
Figure 4. Example histogram of pixel values. Source: Moallem et al. 2012
Why algorithms are important
Algorithms are helpful because they
make our lives easier. While it would be possible for an analyst to identify
and digitize each individual marking from a picture of a gray whale, it would
be extremely time consuming and tedious. Image analysis algorithms significantly
reduce the time it takes to process imagery. A semi-automated algorithm that I
developed to count penguins from still drone imagery can count all the penguins
on a one km2 island in about 30 minutes, while it took me 24 long hours
to count them by hand (Bird et al. in prep). Furthermore, the process
can be repeated with different imagery and analysts as part of a time series
without bias because the algorithm eliminates human error introduced by
different analysts.
Whether it’s a simple combination
of a few processes or a complex series of equations, creating an algorithm requires
breaking down a task to its most basic components. Development involves
translating those components step by step into an automated process, which after
many trials and errors, achieves the desired result. My first algorithm project
took two years of revising, improving, and countless trials and errors. So, whether creating an algorithm or working
to understand one, don’t let the jargon nor the endless trials and errors stop
you. Like most things in life, the key is to have patience and take it one step
at a time.
References
Bird, C. N., Johnston, D.W., Dale, J. (in prep).
Automated counting of Adelie penguins (Pygoscelis adeliae) on Avian and
Torgersen Island off the Western Antarctic Peninsula using Thermal and
Multispectral Imagery. Manuscript in preparation
Fletcher, R. S., Pulich, W. ‡, & Hardegree, B. (2009). A Semiautomated Approach for Monitoring Landscape Changes in Texas Seagrass Beds from Aerial Photography. https://doi.org/10.2112/07-0882.1
Moallem, Payman & Razmjooy, Navid. (2012). Optimal Threshold Computing in Automatic Image Thresholding using Adaptive Particle Swarm Optimization. Journal of Applied Research and Technology. 703.
By: Alexa Kownacki, Ph.D. Student, OSU Department of Fisheries and Wildlife, Geospatial Ecology of Marine Megafauna Lab
Marine mammals are challenging to study for many reasons, and
specifically because they inhabit the areas of the Earth that are uninhabited
by people: the oceans. Monitoring marine mammal populations to gather baselines
on their health condition and reproductive status is not as simple as trap and
release, which is a method often conducted for terrestrial animals. Marine
mammals are constantly moving in vast areas below the surface. Moreover,
cetaceans, which do not spend time on land, are arguably the most challenging
to sample.
One component of my project, based in California, USA, is a health assessment analyzing hormones of the bottlenose dolphins that frequent both the coastal and the offshore waters. Therefore, I am all too familiar with the hurdles of collecting health data from living marine mammals, especially cetaceans. However, the past few decades have seen major advancements in technology both in the laboratory and with equipment, including one tool that continues to be critical in understanding cetacean health: blubber biopsies.
Biopsy dart hitting a bottlenose dolphin below the dorsal fin. Image Source: NMFS
Blubber biopsies are typically obtained via low-powered crossbow with a bumper affixed to the arrow to de-power it once it hits the skin. The arrow tip has a small, pronged metal attachment to collect an eraser-tipped size amount of tissue with surface blubber and skin. I compare this to a skin punch biopsies in humans; it’s small, minimally-invasive, and requires no follow-up care. With a small team of scientists, we use small, rigid-inflatable vessels to survey the known locations of where the bottlenose dolphins tend to gather. Then, we assess the conditions of the seas and of the animals, first making sure we are collecting from animals without potentially lowered immune systems (no large, visible wounds) or calves (less than one years old). Once we have photographed the individual’s dorsal fin to identify the individual, one person assembles the biopsy dart and crossbow apparatus following sterile procedures when attaching the biopsy tips to avoid infection. Another person prepares to photograph the animal to match the biopsy information to the individual dolphin. One scientist aims the crossbow for the body of the dolphin, directly below the dorsal fin, while the another photographs the biopsy dart hitting the animal and watches where it bounces off. Then, the boat maneuvers to the floating biopsy dart to recover the dart and the sample. Finally, the tip with blubber and skin tissue is collected, again using sterile procedures, and the sample is archived for further processing. A similar process, using an air gun instead of a crossbow can be viewed below:
GEMM Lab members using an air gun loaded with a biopsy dart to procure marine mammal blubber from a blue whale in New Zealand. Video Source: GEMM Laboratory.
Part of the biopsy process is holding ourselves to the highest standards in our minimally-invasive technique, which requires constant practice, even on land.
Alexa practicing proper crossbow technique on land under supervision. Image Source: Alexa Kownacki
Blubber is the lipid-rich, vascularized tissue under the
epidermis that is used in thermoregulation and fat storage for marine mammals. Blubber
is an ideal matrix for storing lipophilic (fat-loving) steroid hormones because
of its high fat content. Steroid hormones, such as cortisol, progesterone, and
testosterone, are naturally circulating in the blood stream and are released in
high concentrations during specific events. Unlike blood, blubber is less
dynamic and therefore tells a much longer history of the animal’s nutritional
state, environmental exposure, stress level, and life history status. Blubber
is the cribs-notes version of a marine mammal’s biography over its previous few
months of life. Blood, on the other hand, is the news story from the last 24
hours. Both matrices serve a specific purpose in telling the story, but blubber
is much more feasible to obtain from a cetacean and provides a longer time
frame in terms of information on the past.
A simplified depiction of marine mammal blubber starting from the top (most exterior surface) being the skin surface down to the muscle (most interior). Image Source: schoolnet.org.za
I use blubber biopsies for assessing cortisol, testosterone,
and progesterone in the bottlenose dolphins. Cortisol is a glucocorticoid that
is frequently associated with stress, including in humans. Marine mammals
utilize the same hypothalamic-pituitary-adrenal (HPA) axis that is responsible
for the fight-or-flight response, as well as other metabolic regulations.
During prolonged stressful events, cortisol levels will remain elevated, which
has long-term repercussions for an animal’s health, such as lowered immune
systems and decreased ability to respond to predators. Testosterone and
progesterone are sex hormones, which can be used to indicate sex of the
individual and determine reproductive status. This reproductive information
allows us to assess the population’s composition and structure of males and
females, as well as potential growth or decline in population (West et al.
2014).
Alexa using a crossbow from a small boat off of San Diego, CA. Image Source: Alexa Kownacki
The coastal and offshore bottlenose dolphin ecotypes of interest in my research occupy different locations and are therefore exposed to different health threats. This is a primary reason for conducting health assessments, specifically analyzing blubber hormone levels. The offshore ecotype is found many kilometers offshore and is most often encountered around the southern Channel Islands. In contrast, the coastal ecotype is found within 2 kilometers of shore (Lowther-Thieleking et al. 2015) where they are subjected to more human exposure, both directly and indirectly, because of their close proximity to the mainland of the United States. Coastal dolphins have a higher likelihood of fishery-related mortality, the negative effects of urbanization including coastal runoff and habitat degradation, and recreational activities (Hwang et al. 2014). The blubber hormone data from my project will inform which demographics are most at-risk. From this information, I can provide data supporting why specific resources should be allocated differently and therefore help vulnerable populations. Further proving that the small amount of tissue from a blubber biopsy can help secure a better future for population by adjusting and informing conservation strategies.
Literature Cited:
Hwang, Alice, Richard H Defran, Maddalena Bearzi, Daniela. Maldini, Charles A Saylan, Aime ́e R Lang, Kimberly J Dudzik, Oscar R Guzo n-Zatarain, Dennis L Kelly, and David W Weller. 2014. “Coastal Range and Movements of Common Bottlenose Dolphins (Tursiops Truncatus) off California and Baja California, Mexico.” Bulletin of the Southern California Academy of Sciences 113 (1): 1–13. https://doi.org/10.3390/toxins6010211.
Lowther-Thieleking, Janet L.,
Frederick I. Archer, Aimee R. Lang, and David W. Weller. 2015. “Genetic
Differentiation among Coastal and Offshore Common Bottlenose Dolphins, Tursiops
Truncatus, in the Eastern North Pacific Ocean.” Marine Mammal Science 31
(1): 1–20. https://doi.org/10.1111/mms.12135.
West, Kristi L., Jan Ramer, Janine L. Brown, Jay Sweeney, Erin M. Hanahoe, Tom Reidarson, Jeffry Proudfoot, and Don R. Bergfelt. 2014. “Thyroid Hormone Concentrations in Relation to Age, Sex, Pregnancy, and Perinatal Loss in Bottlenose Dolphins (Tursiops Truncatus).” General and Comparative Endocrinology 197: 73–81. https://doi.org/10.1016/j.ygcen.2013.11.021.
I graduated in March 2017 from the GEMM lab at Oregon State, with a Master’s of Science in Wildlife Management. Graduate school was finally over! No more constant coffee refills, popcorn dinners and overnight library stays; I had submitted my final thesis and I was done! Graduate school was no walk in the park for me, and finishing a master’s or a doctorate degree for anyone is no easy feat! It takes years of hard work, commitment, long hours, and a dedication to learning. I remember feeling both excited and a bit disoriented to be done with this phase of much stress and growth. After submitting my thesis, I took a much-needed month off to unknot the muscles in my back and get myself reacquainted with sunlight. The breath of fresh air was exactly what I needed to recover, but it took no time at all for a new type of challenge to emerge: the arduous task of finding a job.
I did what most job seekers do, I sat behind my computer
applying for opportunities, hit as many roles as I could, and hoped for the
best. Days turned into weeks and weeks turned into months. I was getting
desperate, I resorted to applying for a whole spectrum of roles – consulting,
project management, administration, youth team leader – hoping that something
would land. Soon enough, almost 3 months had passed and I was still in the same
spot as before. I was ready to throw in the towel.
In theory, landing a job after graduation sounds like it should be technically easy because more education should mean you are more qualified for the job, but anyone who has been out of grad school for more than an hour can tell you that landing a job after graduate school can be a long and frustrating process. I did not enter this field and its job prospects blindly – that is, I had a working idea of what type of research career I wanted when I completed my education and how much education I would need to get there. I was aware that navigating the job market in a competitive field could be tricky and time-consuming, especially as a green-job seeker. I knew it would be an added difficulty to land a position near the ocean but also close enough to family (I’m from the Midwest). Or at least, I thought I knew how hard it would be to secure a job. The process turned out to be much harder. Mental preparation alone was not enough and months and months of rejection and feeling stuck within the hamster wheel of the job search cycle was becoming my normal.
So, when I was stuck in the depths of a seemingly fruitless
job search, and trying as hard as I possibly could, it was hard for me to do
anything but roll my eyes, sigh, and give up. But I had to find a way to work
through an apparently endless string of rejection by figuring out some way to
accept, address and navigate my emotions. I needed to take charge of my own
personal development. I started reflecting on what areas of my work on my
master’s thesis that I found most difficult and wanted to improve, and would be
an important component of the job I
wanted. Identifying my own “knowledge gaps” led me to seek out courses,
workshops, job-shadowing and online courses that could fill those holes.
The first thing at the top of my list was to be more
efficient at coding.
Every job description that made me excited to apply had some description of a
coding program: R, Python, MATLAB. I was
lucky enough to attend courses and workshops during my time at the GEMM lab
that provided me much of the code I would need to create my habitat models with
minimal tweaking. On top of that I was surrounded by supervisors and a lab full
of coding geniuses that had an almost, if not completely, open door policy.
When I was stuck and a deadline was quickly approaching, it was great to have
an army of people to help me get through my obstacles. However, I knew if I
wanted to be successful, I needed to become like them: experts and not a
beginner. I purchased a subscription to DataCamp, and started
searching out courses that could help keep my skills fresh and learn new
things. I was over the moon to discover the course “Where are the Fishes?”.
It checked all my boxes: geospatial analysis, R, marine related, acoustics….
perfect. Within this course, there were plenty of DataCamp prerequisites, like
working with data in the tidyverse and working with dates and times in R, so I
had plenty to keep me busy.
I also started looking for in-person, hands-on courses I could enroll in. Since the majority of my marine experience took place on the west coast but I was searching for jobs on the east coast, I enrolled in the Marine Mammal and Sea Turtle Observer Certification Course for the US Atlantic and Gulf of Mexico Oceans in order to learn a little more about identifying species I did not commonly see in nearshore, northern Pacific waters. In this course, I learned about regulations surrounding protected species monitoring, proper camera settings for photographing marine life, and gained the certification needed to work as an observer during seismic surveys for Bureau of Ocean Energy Management (BOEM) and Bureau of Safety and Environmental Enforcement (BSEE) in coordination with the National Marine Fisheries Service. Most of these topics were familiar to me, other than identifying new species, but it was nice to have the refresher and the renewed certification. Heads up this course is coming to Newport in October and I highly recommend it! During this observer course in Charleston, I was able to network with others in the field taking the course, the Charleston aquarium, and the South Carolina DNR. By introducing myself and providing a little bit of my background, I was invited by the South Carolina DNR to watch a satellite tag and release of a sea turtle that the aquarium had been rehabilitating. From the sea turtle release I learned of the International Sea Turtle Symposium that would take place in February in Myrtle Beach, North Carolina and was invited to attend and network by one of the conference chairs, which lead me to my current position. See below…
Releasing of a satellite-tagged Kemp’s Ridley Sea Turtle off Foley Beach, South Carolina
Downtown Charleston
I tried everything I could to keep myself attached to the field. I attended the Biannual Marine Mammal Conference, enrolled in a bioacoustics short course, watched webinars every Friday, read recent journal articles, looked for voluntary work. I even dropped in on offices like NOAA or Universities of towns I was driving through or visiting to see what they were researching, and if they were looking for researchers. Continuous learning and developing took a lot of time, money, and energy but being conscientious about my personal development kept me motivated and engaged. Graduate school prepared me for all of this. My GEMM lab experience taught me to be open to learning, to be flexible and adaptable, to accept, overcome and learn from failures and find solutions. In fact, graduate school provided me a variety of skills that have been transferable to almost everything I have done since graduation.
Manatees spotted while working as a Protected Species Observer in the Gulf, 2018
In December of 2017, I began volunteering at the University of Alabama, Birmingham, under the supervision of Dr. Thane Wibbels, and I began to use those skills I learned from graduate school more than ever. Flash forward and I am now part of a team, called the Kemp’s Ridley Working Group, which is made up of researchers from state, federal and international agencies working together on conservation strategies and programs for Kemp’s Ridley Sea Turtles. Specifically, we are hoping to identify the cues Kemp’s Ridley sea turtles are using to control arribadas (synchronized, large-scale nesting behaviors) in Rancho Nuevo, Mexico. We have a long-term dataset on the number of nests and weather conditions during arribadas from 2007 to 2019 collected using a variety of methods that we are trying to standardize and analyze. Historically, the number of nests has been counted by hand, but over the last few years Dr. Wibbels and his lab have worked to create a protocol for using drones to track the number of sea turtle nests, which has been highly successful. In 2018, the drone recorded the largest sea turtle arribada in 30 years, which consisted of about 4,000 Kemp’s Ridley sea turtle nests within 900 meters of beach.
June 2018 Kemp’s Ridley Sea Turtle Arribada, Rancho Nuevo, Mexico
It’s ironic how incredibly similar my current project is to my
master’s thesis I am gathering environmental data from weather stations and
remote sensing to analyze tides, currents, wind speed, wind direction, water
temperature, air temperature, salinity, etc. in relation to these large
arribadas. I am arguably much faster at this process than I was before due to
my GEMM lab experience. I am quickly
able to recognize when something isn’t right, and am able to debug where I went
wrong. I feel comfortable contributing new ideas and approaches of how to
standardize data from old and new technology, how close to fly drones to the
animals to capture the data we need without animal disturbance, and at what
scales to look for temporal and spatial patterns within our data. The GEMM lab
allowed me to gain knowledge through my own work and by association of my lab
mates projects, trials and tribulations that have directly transferred into
what I am doing now. I am still grant-writing, presenting, collaborating,
managing time, and mentoring – all of which I learned in graduate school. I am also
still coding, and I have joined a local coding group in Birmingham, Bham Quants, and have been asked to give a
series of lectures called “Introduction to R”. The GEMM lab and my own
drawn-out job-hunting process allowed me to end up in the position that I am in
today, and the struggles and cycle of no’s I heard along the way led me to these
opportunities that I am so grateful that I took.
Building on the foundation of my GEMM lab experience, adding my personal development and a couple of years of post-graduate work experience, I no longer feel disoriented. I feel like I have an identity and I know how I want to market myself in the future. I have always considered myself a spatial ecologist, as this is the GEMM labs specializes in, but now I know I’m more of a generalist in terms of species, methods, models and analysis and I want to continue learning and growing in this field to become a jack-of-all-trades. I’ve always had a love for the marine environment, but I also know I have the skills and confidence to transition into terrestrial if I need to. I have fallen in love with geospatial ecology and it isn’t a field that would have even been on my radar, if I had not met Leigh almost 5 years ago *gasp*. Working and studying in the GEMM lab opened up doors for me that I will appreciate for the rest of my life. My advice for anyone studying and working in this field is to stay alert with your eye always on the next step, poised for the next opportunity, whatever it is: to present a paper, attend a conference, meet a scholar in your field, forge a connection, gain a professional skill. There are tons of opportunities (and jobs) that are never posted online, which you will only find out about if you talk to people in your personal network or start knocking on doors. You never know where these doors might lead.
By Dominique Kone, Masters Student in Marine Resource Management
By now, I’m sure you’re aware of recent interests to reintroduce sea otters to Oregon. To inform this effort, my research focuses on predicting suitable sea otter habitat and investigating the potential ecological effects if sea otters are reintroduced in the future. This information will help managers gain a better understanding of the potential for sea otters to reestablish in Oregon, as well as how Oregon’s ecosystems may change via top-down processes. These analyses will address some sources of uncertainties of this effort, but there are still many more questions researchers could address to further guide this process. Here, I note some lingering questions I’ve come across in the course of conducting my research. This is not a complete list of all questions that could or should be investigated, but they represent some of the most interesting questions I have and others have in Oregon.
Credit: Todd Mcleish
The questions, and our associated knowledge on each of these topics:
Is there enough available prey to support a robust sea otter population in Oregon?
Sea otters require approximately 30% of their own body weight in food every day (Costa 1978, Reidman & Estes 1990). With a large appetite, they not only need to spend most of their time foraging, but require a steady supply of prey to survive. For predators, we assume the presence of suitable habitat is a reliable proxy for prey availability (Redfern et al. 2006). Whereby, quality habitat should supply enough prey to sustain predators at higher trophic levels.
In making these habitat predictions for sea otters, we must also recognize the potential limitations of this “habitat equals prey” paradigm, in that there may be parcels of habitat where prey is unavailable or inaccessible. In Oregon, there could be unknown processes unique to our nearshore ecosystems that would support less prey for sea otters. This possibility highlights the importance of not only understanding how much suitable habitat is available for foraging sea otters, but also how much prey is available in these habitats to sustain a viable otter population in the future. Supplementing these habitat predictions with fishery-independent prey surveys is one way to address this question.
Credit: Suzi Eszterhas via Smithsonian Magazine
How will Oregon’s oceanographic seasonality alter or impact habitat suitability?
Sea otters along the California coast exist in an environment with persistent Giant kelp beds, moderate to low wave intensity, and year-round upwelling regimes. These environmental variables and habitat factors create productive ecosystems that provide quality sea otter habitat and a steady supply of prey; thus, supporting high densities of sea otters. This environment contrasts with the Oregon coast, which is characterized by seasonal changes in bull kelp and wave intensity. Summer months have dense kelp beds, calm surf, and strong upwellings. While winter months have little to no kelp, weak upwellings, and intense wave climates. These seasonal variations raise the question as to how these temporal fluctuations in available habitat could impact the number of sea otters able to survive in Oregon.
In Washington – an environment like Oregon – sea otters exhibit seasonal distribution patterns in response to intensifying wave climates. During calm summer months, sea otters primarily forage along the outer coast, but move into more protected areas, such as the Strait of Juan de Fuca, during winter months (Laidre et al. 2009). If sea otters were reintroduced to Oregon, we may very well observe similar seasonal movement patterns (e.g. dispersal into estuaries), but the degree to which this seasonal redistribution and reduction in foraging habitat could impact sea otter reestablishment and recovery is currently unknown.
Credit: Oregon Coast Aquarium
In the event of a reintroduction, do northern or southern sea otters have a greater capacity to adapt to Oregon environments?
In the early 1970’s, Oregon’s first sea otter translocation effort failed (Jameson et al. 1982). Since then, hypotheses on the potential ecological differences between northern and southern sea otters have been proposed as potential factors of the failed effort, potentially due to different abilities to exploit specific prey species. Studies have demonstrated that northern and southern sea otters have slight morphological differences – northern otters having larger skulls and teeth than southern otters (Wilson et al. 1991). This finding has created the hypothesis that the northern otter’s larger skull and teeth allow it to consume prey with denser exoskeletons, and thereby can exploit a greater diversity of prey species. However, there appears to be a lack of evidence to suggest larger skulls and teeth translate to greater bite force. Based on morphology alone, either sub-species could be just as successful in exploiting different prey species.
A different direction to address questions around adaptability is to look at similarities in habitat and oceanographic characteristics. Sea otters exist along a gradient of habitat types (e.g. kelp forests, estuaries, soft-sediment environments) and oceanographic conditions (e.g. warm-temperature to cooler sub-Arctic waters) (Laidre et al. 2009, Lafferty et al. 2014). Yet, we currently don’t know how well or quickly otters can adapt when they expand into new habitats that differ from ones they are familiar with. Sea otters must be efficient foragers and need to acquire skills that allow them to effectively hunt specific prey species (Estes et al. 2003). Hypothetically, if we take sea otters from rocky environments where they’ve developed foraging skills to hunt sea urchins and abalones, and place them in a soft-sediment environment, how quickly would they develop new foraging skills to exploit soft-sediment prey species? Would they adapt quickly enough to meet their daily prey requirements?
Credit: Eric Risberg/Associated Press via The Columbian
In Oregon, specifically, how might climate change impact sea otters, and how might sea otters mediate climate impacts?
Climate change has been shown to directly impact many species via changes in temperature (Chen et al. 2011). Some species have specific thermal tolerances, in which they can only survive within a specified temperature range (i.e. maximum and minimum). Once the temperature moves out of that range, the species can either move with those shifting water masses, behaviorally adapt or perish (Sunday et al. 2012). It’s unclear if and how changing temperatures will impact sea otters, directly. However, sea otters could still be indirectly affected via impacts to their prey. If prey species in sea otter habitat decline due to changing temperatures, this would reduce available food for otters. Ocean acidification (OA) is another climate-induced process that could indirectly impact sea otters. By creating chemical conditions that make it difficult for species to form shells, OA could decrease the availability of some prey species, as well (Gaylord et al. 2011).
Interestingly, these pathways between sea otters and climate change become more complex when we consider the potentially mediating effects from sea otters. Aquatic plants – such as kelp and seagrass – can reduce the impacts of climate change by absorbing and taking carbon out of the water column (Krause-Jensen & Duarte 2016). This carbon sequestration can then decrease acidic conditions from OA and mediate the negative impacts to shell-forming species. When sea otters catalyze a tropic cascade, in which herbivores are reduced and aquatic plants are restored, they could increase rates of carbon sequestration. While sea otters could be an effective tool against climate impacts, it’s not clear how this predator and catalyst will balance each other out. We first need to investigate the potential magnitude – both temporal and spatial – of these two processes to make any predictions about how sea otters and climate change might interact here in Oregon.
Credit: National Wildlife Federation
In Summary
There are several questions I’ve noted here that warrant further investigation and could be a focus for future research as this potential sea otter reintroduction effort progresses. These are by no means every question that should be addressed, but they do represent topics or themes I have come across several times in my own research or in conversations with other researchers and managers. I think it’s also important to recognize that these questions predominantly relate to the natural sciences and reflect my interest as an ecologist. The number of relevant questions that would inform this effort could grow infinitely large if we expand our disciplines to the social sciences, economics, genetics, so on and so forth. Lastly, these questions highlight the important point that there is still a lot we currently don’t know about (1) the ecology and natural behavior of sea otters, and (2) what a future with sea otters in Oregon might look like. As with any new idea, there will always be more questions than concrete answers, but we – here in the GEMM Lab – are working hard to address the most crucial ones first and provide reliable answers and information wherever we can.
References:
Chen, I., Hill, J. K., Ohlemuller, R., Roy, D. B., and C. D. Thomas. 2011. Rapid range shifts of species associated with high levels of climate warming. Science. 333: 1024-1026.
Costa, D. P. 1978. The ecological energetics, water, and electrolyte balance of the California sea otter (Enhydra lutris). Ph.D. dissertation, University of California, Santa Cruz.
Estes, J. A., Riedman, M. L., Staedler, M. M., Tinker, M. T., and B. E. Lyon. 2003. Individual variation in prey selection by sea otters: patterns, causes and implications. Journal of Animal Ecology. 72: 144-155.
Gaylord et al. 2011. Functional impacts of ocean acidification in an ecologically critical foundation species. Journal of Experimental Biology. 214: 2586-2594.
Jameson, R. J., Kenyon, K. W., Johnson, A. M., and H. M. Wight. 1982. History and status of translocated sea otter populations in North America. Wildlife Society Bulletin. 10(2): 100-107.
Krause-Jensen, D., and C. M. Duarte. 2016. Substantial role of macroalgae in marine carbon sequestration. Nature Geoscience. 9: 737-742.
Lafferty, K. D., and M. T. Tinker. 2014. Sea otters are recolonizing southern California in fits and starts. Ecosphere.5(5).
Laidre, K. L., Jameson, R. J., Gurarie, E., Jeffries, S. J., and H. Allen. 2009. Spatial habitat use patterns of sea otters in coastal Washington. Journal of Marine Mammalogy. 90(4): 906-917.
Redfern et al. 2006. Techniques for cetacean-habitat modeling. Marine Ecology Progress Series. 310: 271-295.
Reidman, M. L. and J. A. Estes. 1990. The sea otter (Enhydra lutris): behavior, ecology, and natural history. United States Department of the Interior, Fish and Wildlife Service, Biological Report. 90: 1-126.
Sunday, J. M., Bates, A. E., and N. K. Dulvy. 2012. Thermal tolerance and the global redistribution of animals. Nature: Climate Change. 2: 686-690.
Wilson, D. E., Bogan, M. A., Brownell, R. L., Burdin, A. M., and M. K. Maminov. 1991. Geographic variation in sea otters, Ehydra lutris. Journal of Mammalogy. 72(1): 22-36.
By Christina Garvey, University of Maryland, GEMM Lab REU Intern
It is July 8th and it is my 4th week here in Hatfield as an REU intern for Dr. Leigh Torres. My name is Christina Garvey and this summer I am studying the spatial ecology of blue whales in the South Taranaki Bight, New Zealand. Coming from the east coast, Oregon has given me an experience of a lifetime – the rugged shorelines continue to take my breath away and watching sea lions in Yaquina Bay never gets old. However, working on my first research project has by far been the greatest opportunity and I have learned so much in so little time. When Dr. Torres asked me to contribute to this blog I was unsure of how I would write about my work thus far but I am excited to have the opportunity to share the knowledge I have gained with whoever reads this blog post.
The research project that I will be conducting this summer will use remotely sensed environmental data (information collected from satellites) to predict blue whale distribution in the South Taranaki Bight (STB), New Zealand. Those that have read previous blogs about this research may remember that the STB study area is created by a large indentation or “bight” on the southern end of the Northern Island. Based on multiple lines of evidence, Dr. Leigh Torres hypothesized the presence of an unrecognized blue whale foraging ground in the STB (Torres 2013). Dr. Torres and her team have since proved that blue whales frequent this region year-round; however, the STB is also very industrial making this space-use overlap a conservation concern (Barlow et al. 2018). The increasing presence of marine industrial activity in the STB is expected to put more pressure on blue whales in this region, whom are already vulnerable from the effects of past commercial whaling (Barlow et al. 2018) If you want to read more about blue whales in the STB check out previous blog posts that talk all about it!
Figure 1. A blue whale surfaces in front of a floating production storage and offloading vessel servicing the oil rigs in the South Taranaki Bight. Photo by D. Barlow.
Figure 2. South Taranaki Bight, New Zealand, our study site outlined by the red box. Kahurangi Point (black star) is the site of wind-driven upwelling system.
The possibility of the STB as an important foraging ground for a resident population of blue whales poses management concerns as New Zealand will have to balance industrial growth with the protection and conservation of a critically endangered species. As a result of strong public support, there are political plans to implement a marine protected area (MPA) in the STB for the blue whales. The purpose of our research is to provide scientific knowledge and recommendations that will assist the New Zealand government in the creation of an effective MPA.
In order to create an MPA that would help conserve the blue whale population in the STB, we need to gather a deeper understanding of the relationship between blue whales and this marine environment. One way to gain knowledge of the oceanographic and ecological processes of the ocean is through remote sensing by satellites, which provides accessible and easy to use environmental data. In our study we propose remote sensing as a tool that can be used by managers for the design of MPAs (through spatial and temporal boundaries). Satellite imagery can provide information on sea surface temperature (SST), SST anomaly, as well as net primary productivity (NPP) – which are all measurements that can help describe oceanographic upwelling, a phenomena that is believed to be correlated to the presence of blue whales in the STB region.
Figure 3. The stars of the show: blue whales. A photograph captured from the small boat of one animal fluking up to dive down as another whale surfaces close by. (Photo credit: L. Torres)
Past studies in the STB showed evidence of a large upwelling event that occurs off the coast of Kahurangi Point (Fig. 2), on the northwest tip of the South Island (Shirtcliffe et al. 1990). In order to study the relationship of this upwelling to the distribution of blue whales, I plan to extract remotely sensed data (SST, SST anomaly, & NPP) off the coast of Kahurangi and compare it to data gathered from a centrally located site within the STB, which is close to oil rigs and so is of management interest. I will first study how decreases in sea surface temperature at the site of upwelling (Kahurangi) are related to changes in sea surface temperature at this central site in the STB, while accounting for any time differences between each occurrence. I expect that this relationship will be influenced by the wind patterns, and that there will be changes based on the season. I also predict that drops in temperature will be strongly related to increases in primary productivity, since upwelling brings nutrients important for photosynthesis up to the surface. These dips in SST are also expected to be correlated to blue whale occurrence within the bight, since blue whale prey (krill) eat the phytoplankton produced by the productivity.
Figure 4. A blue whale lunges on an aggregation of krill. UAS piloted by Todd Chandler.
To test the relationships I determine between remotely sensed data at different locations in the STB, I plan to use blue whale observations from marine mammal observers during a seismic survey conducted in 2013, as well as sightings recorded from the 2014, 2016, and 2017 field studies led by Dr. Leigh Torres. By studying the statistical relationships between all of these variables I hope to prove that remote sensing can be used as a tool to study and understand blue whale distribution.
I am very excited about this research, especially because the end goal of creating an MPA really gives me purpose. I feel very lucky to be part of a project that could make a positive impact on the world, if only in just a little corner of New Zealand. In the mean time I’ll be here in Hatfield doing the best I can to help make that happen.
References:
Barlow DR, Torres LG, Hodge KB, Steel D, Baker CS, Chandler TE, Bott N, Constantine R, Double MC, Gill P, Glasgow D, Hamner RM, Lilley C, Ogle M, Olson PA, Peters C, Stockin KA, Tessaglia-hymes CT, Klinck H (2018) Documentation of a New Zealand blue whale population based on multiple lines of evidence. Endanger Species Res 36:27–40.
Shirtcliffe TGL, Moore MI, Cole AG, Viner AB, Baldwin R, Chapman B (1990) Dynamics of the Cape Farewell upwelling plume, New Zealand. New Zeal J Mar Freshw Res 24:555–568.
Torres LG (2013) Evidence for an unrecognised blue whale foraging ground in New Zealand. New Zeal J Mar Freshw Res 47:235–248.