Dr. Alejandro A. Fernández Ajó, Postdoctoral Scholar, Marine Mammal Institute – OSU Department of Fisheries, Wildlife, & Conservation Sciences, Geospatial Ecology of Marine Megafauna (GEMM) Lab.
Whales are exposed to an increasing number of human-induced stressors—ranging from pollution and bycatch to the impacts of climate change on prey quality and distribution. Understanding how these factors affect whale health is critical for their conservation. The use of alternative approaches (i.e., alternative to blood samples) for gathering physiological information on large whales using a variety of non-lethal and non to minimally invasive sample matrices (i.e., blubber biopsies, blow, and fecal samples) provides a window into their endocrine state, allowing researchers to assess how these animals respond to both short-term and long-term stressors, and assess their reproductive and nutritional status. However, a lack of standardized protocols might hinder the comparability of results across studies, making it difficult to draw broad conclusions about the health and reproductive parameters of different whale populations.
Dr. Logan Pallin and I organized a lab exchange, funded by The Company of Biologists, to start a new collaboration aimed at bridging this gap by validating and standardizing methods for endocrine assessments in whale blubber. This is not just a technical exercise; it is a foundational step towards building equity and capacity in laboratories worldwide to conduct reliable and comparable endocrine assessments, enhancing the opportunities for multi-lab collaborations. Through this exchange, we aim to consolidate a standardized approach that will yield consistent results between laboratories, enabling better comparisons across different large whale populations. Hosted by the University of California Santa Cruz Biotelemetry and Behavioral Ecology Lab (UCSC-BTBEL Lab) under the mentorship of Dr. Logan Pallin, this experience is instrumental in advancing my research on large whale ecology and conservation.
Dr. Logan Pallin and Alejandro Fernandez Ajó conducting hormone extractions from gray whale blubber samples (left). Preparing a microtiter assay plate for hormone quantification in blubber (right).
During this exchange at the BBE Lab, I had the privilege of working closely with Dr. Logan Pallin, whose expertise in large whale endocrinology (particularly analyzing blubber biopsies) has been instrumental in shaping modern approaches to whale research. The lab’s cutting-edge equipment and Logan’s extensive experience with hormone extraction and quantification methods provided an ideal setting for refining our protocols. Our work focused on the extraction and quantification of progesterone from gray whale blubber samples provided by the Oregon State University Marine Mammal Stranding Network, part of MMI. These large blubber sections allow for repeated sub-sampling to ensure that the selected immunoassays reliably detect and measure the hormones of interest, while also assessing potential sources of variability when applying a standardized protocol. We initially focused our tests and validations on progesterone, as it is the precursor of all major steroid hormones and serves as an indicator of reproductive state in females.
A fieldwork day off Monterrey Bay, California with Dr. Logan Pallin, and PhD candidate Haley Robb. Blubber. Blubber biopsies can be obtained from free swimming whales with minimally invasive methods. From each sample we can derive multiple information about the reproductive status, genetics and overall health of the individuals.
The broader impact of our work The successful validation and standardization of these protocols represents a significant advancement in whale conservation physiology. Once these methods are established, we plan to acquire funds to apply them to a larger collection of blubber samples. We hope to expand our work to include other species and regions, building a broader network of researchers dedicated to studying large whales in a rapidly changing world, and to assess hormone profiles in relation to factors like reproductive success, body condition, and exposure to stressors such as vessel traffic and environmental changes.
During our fieldwork in Monterey Bay, we had fascinating encounters with Minke whales (Balaenoptera acutorostrata, top left), a large group of Risso’s dolphins (Grampus griseus, bottom left), playful Humpbacks (Megaptera novaeangliae, top right), and a Blue whale (Balaenoptera musculus, no photo).
As I conclude this lab exchange, I am filled with excitement for the future. The knowledge and skills gained during this experience will undoubtedly shape the next phase of my research, allowing me to contribute more effectively to the conservation of these incredible animals. I look forward to applying these standardized methods to ongoing and future projects, and to continuing this fruitful collaboration with the BBE Lab. This journey has reinforced the importance of collaboration, standardization, and innovation in the field of conservation physiology. By working together, we can better understand the complex lives of large whales and take meaningful steps towards their protection in an increasingly challenging environment.
Acknowledgments: This exchange was made possible by the support of The Company of Biologists Traveling Fellowship Grant. I would like to thank Dr. Ari Friedlaender (BBE Lab PI) for facilitating this exchange, and Dr. Leigh Torres (GEMM Lab PI) and Dr. Lisa Balance (MMI director) for their support in helping me expand my collaboration network and skillsets. Special thanks to PhD student Haley Robb for her assistance in the laboratory and fieldwork, and a heartfelt thank you to Dr. Logan Pallin for generously sharing his knowledge and time.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly message when we post a new blog. Just add your name and email into the subscribe box below.
Dr. Alejandro A. Fernández Ajó, Postdoctoral Scholar, Marine Mammal Institute – OSU Department of Fisheries, Wildlife, & Conservation Sciences, Geospatial Ecology of Marine Megafauna (GEMM) Lab.
The Eastern North Pacific (ENP) gray whale population has experienced at least two recorded Unusual Mortality Events (UMEs), from 1999–2000 and from 2019 to 2024, during which many gray whales stranded along the Pacific coast from northern Mexico to the Alaskan Arctic, USA (Martínez-Aguilar et al., 2019; Urbán, 2020). Several factors have been considered as possible causes for the high number of gray whale’s strandings, including variation in Arctic prey availability and the duration of their feeding season caused by the timing of sea ice formation and breakup (Stewart et al., 2023), starvation, anthropogenically derived toxicants, biotoxins, infectious diseases, parasites, fisheries interactions, and ship strikes (F. Gulland et al., 2005). In the most recent UME, many of the stranded whales showed signs of emaciation, indicating malnutrition as a causal factor of death (Christiansen et al., 2021; Torres et al., 2022). While the poor condition of many of the stranded whales supports the idea of starvation as a cause for these mortalities, the underlying causes of malnutrition are unknown, and it is also unclear whether the whales’ decline in body condition was rapid or gradual.
Large whales face a multitude of stressors in their environment, ranging from ocean noise to contaminants, climate change, and prey shifts. Understanding how individual whales respond to these disturbances is crucial for assessing potential impacts on the population as a whole. However, monitoring the health parameters and vital rates of whales presents significant challenges due to their large size, mobility, and the vast ranges of their marine habitat. Studying stranded whales can provide valuable insights into health risks, disease susceptibility, and the impacts of pollutants and other stressors on whale populations, thus informing conservation strategies (see post). Nonetheless, the quality of information obtained from necropsies heavily relies on the timeliness of stranding reports, as decomposition begins immediately after death, limiting detailed investigations into the cause of death. Therefore, establishing a robust network capable of promptly reporting and addressing stranding events is essential (Gulland & Stockin, 2020). An effective network involves having well-trained staff, proper infrastructure, sufficient funding, and the expertise and tools necessary to gather and analyze data and samples to infer their health and causes of mortality.
During my doctoral dissertation, I worked to develop and ground truth the endocrine analyses of whale baleen as a novel sample type that can be used for retrospective assessments of the whale’s physiology (see my previous post & post). Baleen, the filter-feeding apparatus of mysticetes whales (Figure 1), consists of long fringed plates of keratinized tissue that grow continuously and slowly downward from the whale’s upper jaw. These plates are routinely collected at necropsies; and unlike other tissue types, they are durable and have minimum storage requirements; they can be preserved dry at room temperature, allowing for the analysis of both historical and current whale populations. Moreover, while most sample types used for studying whale health and physiology provide a single time-point measure of current circulating hormone levels (e.g., skin or respiratory vapor) or hold integrated information from the previous few hours or days (e.g., urine and feces), baleen tissue provides a unique opportunity for retrospective and longitudinal analyses of multiple biological parameters of the individual during the time that the tissue was grown, i.e., months to years prior to death, helping to describe the whale’s physiology, migration patterns, and exposure to pollutants (see my previous post).
In our recent study, “A longitudinal study of endocrinology and foraging ecology of subadult gray whales prior to death based on baleen analysis”, published in the journal General and Comparative Endocrinology, we examine isotope and hormone levels in the baleen of five young gray whales stranded in central Oregon during the most recent UME. Our primary objectives were to retrospectively examine the hormone and isotopic profiles of the individual whales prior to mortality, assess potential factors contributing to death, and identify the timing for the onset of chronic illness leading to mortality. Our analysis included tracing longitudinal changes in (1) stable isotope values in baleen (δ13C and δ15N), which allowed us to infer the baleen growth rate and assess the seasonal changes in diet and foraging location in large whales (Figure 2), along with the quantification of (2) two adrenal glucocorticoid steroids that are biomarkers for the whale’s stress response, (3) one thyroid hormone (triiodothyronine, T3) as an indicator of nutritional state, and (4) two sex hormones, progesterone and testosterone, to infer about reproductive status and sexual maturity. By integrating isotopic and hormonal methodologies, our study demonstrates how baleen analysis offers a comprehensive narrative of the endocrine and trophic ecology of individual whales over time.
Figure 2. Gray whales, like other large marine mammals that rely on built-up energy reserves, exhibit distinct seasonal shifts in their feeding habits. During summer, these whales feed at the ocean’s bottom, consuming organisms lower in the food chain, which is reflected in lower nitrogen values in their baleen (summer foraging). In winter, however, they must rely on their own fat reserves, causing an increase in nitrogen values (wintering). In this plot we can observe the oscillations in δ15N over time; this information allows us to estimate the baleen growth rate. Our results suggest that gray whale baleen holds a record of around 1.3 years of stable isotopes and hormone data prior to the time of death (Fernandez Ajo et al. 2024). The red cross in the X-axis, indicate the time of death. Gray whale illustration https://www.fisheries.noaa.gov/species/gray-whale
Our endocrine assessments revealed detailed profiles of stress-related hormones (glucocorticoids, cortisol) and thyroid hormones along the lengths of the baleen. We found increased levels of cortisol in whales that died from unknown causes, starting about eight months prior to their deaths. This suggests these whales were under prolonged stress before dying. In contrast, in the case of a whale killed acutely by a killer whale, cortisol levels were low and constant prior to death, indicating this individual was likely in good health prior to the sudden attack. In terms of thyroid activity, indicated by T3 hormone levels, we found a gradual increase over several months in the whales that died of unknown causes. This pattern is not typically expected, as stress usually suppresses thyroid function. This anomaly could suggest an adaptive response to maintain body temperature and metabolism in potentially malnourished whales. Regarding the sex hormones, as expected for this age class, we found no significant fluctuations or spikes that would indicate sexual maturity in these young whales (Figure 3).
Figure 3. Longitudinal hormone profiles in an individual gray whale that died due to unknown causes (left) and one that died acutely due to orca predation (right). Note the pronounced elevations in cortisol levels (indicative of stress) and T3 prior to death in the case of unknown cause of death, while hormone levels remained low and constant prior to death in the whale acutely killed. Sex hormones do not present any clear oscillations, indicating that these whales were likely sexually immature. The red cross in the X-axis, indicate the time of death. Killer whale (Orcinus orca) illustration https://www.fisheries.noaa.gov/species/killer-whale
Although commercial whaling is currently banned and several whale populations show evidence of recovery, today’s whales are exposed to a variety of other human stressors that cause significant lethal and non-lethal impacts (e.g., entanglement in fishing gear, vessel strikes, shipping noise, climate change, etc.; reviewed in Thomas et al., 2016). The recovery and conservation of large whale populations is particularly important to the oceanic environment due to their key ecological role and unique biological traits (See my previous post). Our research demonstrates the strengths of using baleen as a tool for the retrospective assessments of whale endocrinology and trophic ecology. As the Eastern North Pacific gray whale population faces recurring challenges, indicated by fluctuating numbers and unusual mortality events, innovative techniques like the baleen analysis presented here, are essential to investigate the causes of mortality and inform management, helping us understand not only the immediate causes of death but also broader environmental and ecological changes affecting their survival. Broadly implementing this approach with a greater sample size of baleen collected across a larger spatial and temporal range could significantly improve our strategies for conservation and management of baleen whales.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly message when we post a new blog. Just add your name and email into the subscribe box below.
References
Christiansen, F., Rodríguez-González, F., Martínez-Aguilar, S., Urbán, J., Swartz, S., Warick, H., Vivier, F., & Bejder, L. (2021). Poor body condition associated with an unusual mortality event in gray whales. Marine Ecology Progress Series, 658, 237–252. https://doi.org/10.3354/meps13585
Gulland, F. M. D., & Stockin, K. A. (2020). Harmonizing global strandings response. European Cetacean Society Special Publication Series.
Gulland, F., Pérez-Cortés, H., Urbán, J. R., Rojas-Bracho, L., Ylitalo, G., Weir, J., Norman, S., Muto, M., Rugh, D., Kreuder, C., & Rowles, T. (2005). Eastern North Pacific gray whale (Eschrichtius robustus) unusual mortality event, 1999-2000. U.S. Department of Commerce. NOAA Technical Memorandum. NMFS-AFSC-150., March, 33 pp. http://www.afsc.noaa.gov/publications/AFSC-TM/NOAA-TM-AFSC-150.pdf
Martínez-Aguilar, S., Mariano-Meléndez, E., López-Paz, N., Castillo-Romero, F., Zaragoza-aguilar, G. A., Rivera-Rodriguez, J., Zaragoza-Aguilar, A., Swartz, S., Viloria-Gómora, L., & Urbán, J. R. (2019). Gray whale (Eschrichtius robustus) stranding records in Mexico during the winter breeding season in 2019. Report of the International Whaling Commission. Document SC/68A/CMP/14, May.
Stewart, J. D., Joyce, T. W., Durban, J. W., Calambokidis, J., Fauquier, D., Fearnbach, H., Grebmeier, J. M., Lynn, M., Manizza, M., Perryman, W. L., Tinker, M. T., & Weller, D. W. (2023). Boom-bust cycles in gray whales associated with dynamic and changing Arctic conditions. Science, 382(6667), 207–211. https://doi.org/10.1126/science.adi1847
Torres, L. G., Bird, C. N., Rodríguez-González, F., Christiansen, F., Bejder, L., Lemos, L., Urban R, J., Swartz, S., Willoughby, A., Hewitt, J., & Bierlich, KC. (2022). Range-Wide Comparison of Gray Whale Body Condition Reveals Contrasting Sub-Population Health Characteristics and Vulnerability to Environmental Change. Frontiers in Marine Science, 9. https://doi.org/10.3389/fmars.2022.867258
Urbán, R. (2020). Gray whale stranding records in Mexico during the 2020 winter breeding season. Unpublished Paper SC/68B/CMP/13 Presented to the IWC Scientific Committee, Cambridge.
Dr. Alejandro A. Fernández Ajó, Postdoctoral Scholar, Marine Mammal Institute – OSU Department of Fisheries, Wildlife, & Conservation Sciences, Geospatial Ecology of Marine Megafauna (GEMM) Lab.
A year in a baleen whale life typically involves migrating between polar or subpolar “feeding grounds” in summer and subtropical “breeding grounds” in winter. Calves are typically born during a specific portion of the winter months (Lockyer and Brown, 1981), suggesting a regular alternation between reproductively active and inactive states (Bronson, 1991). Seasonal reproduction in mammals often includes pronounced annual cycles in reproductive hormones triggered by changes in the photoperiod or other environmental cues, along with endogenous circannual cycles (Hau 2007).
Testosterone (T), a key reproductive hormone, is crucial for male spermatogenesis (development of sperm) and influences behaviors such as courtship, mating, and male to male competition. Seasonally breeding mammals exhibit an annual peak in T. The amplitude of T can be influenced by age, with immature males having low T levels that rise sharply at sexual maturity (Beehner et al. 2009; Chen et al. 2009) and then, in some species, declines in the older males (i.e., reproductive senescence; Hunt et al. 2020; Chen et al. 2009). This variability, combined with social cues and exposure to stressors, contributes to individual differences in hormone patterns.
Seasonal testosterone patterns are well-documented in many vertebrate males, including terrestrial mammals, pinnipeds, and odontocetes (Wells, 1984; Kellar et al., 2009; Funasaka et al., 2011, 2018; O’Brien et al., 2016; Richard et al., 2017). However, our understanding of seasonal patterns of testosterone in large whales, especially baleen whales, remains incomplete due to their cryptic nature. Improved understanding of cyclic changes in male reproductive hormones could enhance population management and conservation of whale species. For instance, a clear comprehension of male testosterone cycling in a species can potentially improve the accuracy of sex identification for unknown individuals through hormone ratios. It can also aid in better discriminating sexually active adults from juveniles, understanding the age of sexual maturity (often challenging to determine in males), the potential occurrence of reproductive senescence in older males, and determining the month and location of the conceptive season—which, in turn, may inform estimates of gestation length in females. Insight into these aspects of baleen whale reproductive biology would enhance our ability to understand variation in population abundance and vital rates.
Recent advancements in hormone extraction from non-plasma (blood) samples, such as blow, fecal, blubber, earplugs, and baleen, offer new avenues for studying baleen whale physiology (Hunt et al., 2013). However, obtaining repeated samples from an individual, and over an extended period, from whales to assess hormone patterns is challenging. In this context, earplug endocrine analyses, focusing on cerumen layers (ear wax), have provided insights into sexual maturity in male blue whales (Trumble et al., 2013). However, the temporal resolution (e.g., years) in this sample type limits the detection of seasonal patterns. On the other hand, baleen data provides longitudinal information with sufficient resolution for understanding male reproductive biology and it has been successfully applied to the study of whale species with longer baleen plates (over a decade of an individual’s life), such as the bowhead whale, North Atlantic right whale, and a blue whale (Hunt et al., 2018; Hunt et al., 2020). Additionally, seasonal trends in testosterone have been documented in male humpback whales through blubber biopsy analyses (Cates et al. 2019).
Photos: This is Orange Knuckles (AKA OK). He is one of the males that regularly visit the Oregon coast. He was first observed in 2005, which means he is an adult male and is at least 19 years old (as of 2024). Do you want to learn more about him and other PCFG whales that frequent the Oregon coast? Visit IndividuWhale. Credit: GEMM Lab.
With the GEMM Lab’s GRANITE project, we are delving into an eight-year dataset of individual gray whale morphometrics and fecal hormone data to investigate important aspects of male reproduction in detail. Our non-invasive data collection methods (fecal samples and drone overflights) allow important repeated measurements of the same individual throughout and between foraging seasons. Preliminary results from our analysis reveal a significant association of the day of the year with elevation in T, suggesting that in the late summer the Oregon Coast could be an important area for gray whale social behavior in preparation for reproduction. Furthermore, we are uncovering an association between age and T levels, highlighting the potential for us to identify the age for onset of sexual maturity in males. Additionally, we are exploring the relationship between T levels, exposure to stressors, body condition, and other factors that might influence male reproductive attempts. These data will provide valuable information for conservation and management efforts, aiding in critical habitat identification and reproductive timing for gray whales. Stay tuned for the new results to come!
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly message when we post a new blog. Just add your name and email into the subscribe box below.
References
Beehner JC, Gesquiere L, Seyfarth RM, Cheney DL, Alberts SC, Altmann J. 2009. Testosterone related to age and life-history stages in male baboons and geladas. Horm Behav 56:472-80.
Bronson FH (1991) Mammalian Reproductive Biology. University of Chicago Press, Chicago, IL.
Buck CL, Barnes BM. 2003. Androgen in free-living arctic ground squirrels: seasonal changes and influence of staged male-male aggressive encounters. Horm Behav 43:318-26.
Cates KA, Atkinson S, Gabriele CM, Pack AA, Straley JM, Yin S. 2019. Testosterone trends within and across seasons in male humpback whales (Megaptera novaeangliae) from Hawaii and Alaska. Gen Comp Endocrinol 279:164-73.
Chen H, Ge R-S, Zirkin BR. 2009. Leydig cells: from stem cells to aging. Mol Cell Endocrinol 306:9-16.
Funasaka N, Yoshioka M, Suzuki M, Ueda K, Miyahara H, Uchida S (2011) Seasonal difference of diurnal variations in serum melatonin, cortisol, testosterone, and rectal temperature in Indo-Pacific bottlenose dolphins (Tursiops aduncus). Aquatic Mamm 37: 433–443.
Hau M. 2007. Regulation of male traits by testosterone: implications for the evolution of vertebrate life histories. BioEssays 29:133-44.
Hunt KE, Moore MJ, Rolland RM, Kellar NM, Hall AJ, Kershaw J, Raverty SA, Davis CE, Yeates LC, Fauquier DA. 2013. Overcoming the challenges of studying conservation physiology in large whales: a review of available methods. Cons Physiol 1:cot006.
Hunt KE, Buck CL, Ferguson S, Fernández Ajo A., Heide-Jørgensen MP, Matthews CJD, Male Bowhead Whale Reproductive Histories Inferred from Baleen Testosterone and Stable Isotopes, Integrative Organismal Biology, Volume 4, Issue 1, 2022, obac014 https://doi.org/10.1093/iob/obac014
Kellar N, Trego M, Marks C, Chivers S, Danil K (2009) Blubber testosterone: a potential marker of male reproductive status in shortbeaked common dolphins. Mar Mamm Sci 25: 507–522
Lockyer C, Brown S (1981) The migration of whales. In Aldley D, ed. Animal Migration Society for Experimental Biology Seminar Series, Book 13. Cambridge University Press, Cambridge, England.
O’Brien JK, Steinman KJ, Fetter GA, Robeck TR (2016) Androgen and glucocorticoid production in the male killer whale (Orcinus orca): influence of age, maturity, and environmental factors. Andrology 5: 180–190.
Richard JT, Robeck TR, Osborn SD, Naples L, McDermott A, LaForge R, Romano TA, Sartini BL (2017) Testosterone and progesterone concentrations in blow samples are biologically relevant in belugas (Delphinapterus leucas). Gen Comp Endocrinol 246: 183–193.
Trumble S, Robinson E, Berman-Kowalewski M, Potter C, Usenko S (2013) Blue whale earplug reveals lifetime contaminant exposure and hormone profiles. Proc Nat Acad Sci 110: 16922–16926.
Wells RS (1984) Reproductive behavior and hormonal correlates in Hawaiian spinner dolphins (Stenella longirostris). In Perrin WR, Brownell RL Jr, DeMaster DP, eds. Reproduction in Whales, Dolphins, and Porpoises. Cambridge: Reports of the International Whaling Commission, pp 465–472.
Dr. Alejandro A. Fernández Ajó, Postdoctoral Scholar, Marine Mammal Institute – OSU Department of Fisheries, Wildlife, & Conservation Sciences, Geospatial Ecology of Marine Megafauna (GEMM) Lab.
In a previous post (link to blog), I discussed the crucial importance of acquiring knowledge on the reproductive parameters of individual animals in wild populations for designing effective strategies in conservation biology. Specifically, the ability to quantify the number of pregnancies within a population offers valuable insights into the health of individual females and the population as a whole [1,2]. This knowledge provides tools to describe important life-history parameters, including the age of sexual maturity, frequency of pregnancy, duration of gestation, timing of reproduction, and population fecundity; all of which are essential components for monitoring trends in reproduction and the overall health of a species [3]. Additionally, I explained some of the challenges inherent in obtaining such information when working with massive wild animals that spend most of their time underwater in vast expanses of the oceans. Yes, I am talking about whales.
As a result of the logistical and methodological challenges that involve the study of large whales, detailed knowledge of the life-history and general reproductive biology of whales is sparse for most species and populations. In fact, much of the available information is derived from whaling records [4], which may be outdated for application in population models [5].
If you are an avid reader of the GEMM Lab blog posts, you might be familiar with the gray whale (Eschrichtius robustus), and with the distinct subgroup of gray whales, known as the Pacific Coast Feeding Group (PCFG). PCFG gray whales are characterized by their shorter migration to spend their feeding season in the coastal waters of Northern California, Oregon, and southeastern Alaska [6], relative to the larger Eastern North Pacific gray whale that forage in the Arctic region.
The GEMM Lab has monitored individual gray whales within the PCFG off the Oregon coast since 2016 (check the GRANITE project). Each individual whale presents a unique pigmentation pattern, or unique marks that we can use to identify who is who among the whales who visit the Oregon coast. In this way, we keep a detailed record of re-sightings of known individuals (visit IndividuWhale to learn more), and we have high individual re-sighting rates, resulting in a long-term data series for individual whales which enables us to monitor their health, body condition, and thus further develop and advance our non-invasive study methods.
Drone-based image of a Gray whale defecating. Source: GEMM Lab, NOAA/NSF permit #16111
In our recently manuscript published in the Royal Society Open Science journal, armed with our robust dataset comprising fecal hormone metabolites, drone-based photogrammetry, and individual sightings, we delved into the strengths and weaknesses of various diagnostic tools for non-invasive pregnancy diagnosis. Ultimately, we propose a methodological approach that can help with the challenging and important task of identifying pregnancies in gray whales. In particular, we explored the variability in fecal progesterone metabolites and body morphology relative to observed reproductive status and estimated the pregnancy probability for mature females using statistical models.
In mammals, the progesterone hormone is secreted in the ovaries during the estrous cycle and gestation, making it the predominant hormone responsible for sustaining pregnancy [7]. As the hormones are cleared from the blood into the gut, they are metabolized and eventually excreted in feces; fecal samples represent a cumulative and integrated concentration of hormone metabolites [8;9], which are useful indicators for endocrine assessments of free-swimming whales. Additionally, our previous studies in this population [10] detected differences in body condition (see KC blog for more details about how we measure whales) that suggest that changes in the whale’s body widths could be useful in detecting pregnancies.
Our exploratory analyses show that in individual whales, the levels of fecal progesterone were elevated when pregnant as compared to when the same whale was not pregnant. But when looking at progesterone levels at the population level, these differences were masked with the intrinsic variability of this measurement. In turn, the body morphometrics, in particular the body width at the 50% of the total body length, helped discriminate pregnancies better, and the statistical models that included this width variable, effectively classified pregnant from non-pregnant females with a commendable accuracy. Thus, our morphometric approach showcased its potential as a reliable alternative for pregnancy diagnosis.
Below, a comparison of body widths at 5% increments along total body length (from 20 % to 70 %) in female gray whales of known reproductive status from UAS-based photogrammetry (example photograph shown at top). Pregnant females (PF; in blue), presumed nonpregnant juvenile females (JF; yellow), and lactating females (LF; orange). Fernandez Ajó et al. 2023.
Notably, when we ran the pregnancy prediction models on data from our 2022 season and compared results with observations of whales in 2023, we identified a known whale from our study area “Clouds” accompanied by a calf, indicating that she was pregnant in 2022. Our model predicted Clouds to be pregnant with a 70% probability. This validation lends strong confidence to our approach to diagnosing pregnancy. Conversely, some whales predicted to be pregnant in 2022 were not observed with a calf during the 2023 season. However, the absence of calves accompanying these females is likely due to the relatively high mortality of newborn calves in gray whales due to predation or other causes [11].
Overall, our findings underscore some limitations of fecal progesterone metabolite in accurately identifying pregnant PCFG gray whales. However, while acknowledging the challenges associated with fecal sample collection and hormone analysis, we advocate for ongoing exploration of alternative hormone quantification methods and antibodies. Our study highlights the importance of continued research in refining these techniques. The unique attributes of our study system, including high individual re-sighting rates and non-invasive fecal hormone analysis, position it as a cornerstone for future advancements in understanding gray whale reproductive health. By improving our ability to monitor reproductive metrics in baleen whale populations, we pave the way for more effective conservation strategies, ensuring the resilience of these magnificent creatures in the face of a changing marine ecosystems.
References
[1] Burgess EA, Lanyon JM, Brown JL, Blyde D, Keeley T. 2012 Diagnosing pregnancy in free-ranging dugongs using fecal progesterone metabolite concentrations and body morphometrics: A population application. Gen Comp Endocrinol 177, 82–92. (doi:10.1016/J.YGCEN.2012.02.008)
[2] Slade NA, Tuljapurkar S, Caswell H. 1998 Structured-Population Models in Marine, Terrestrial, and Freshwater Systems. J Wildl Manage 62. (doi:10.2307/3802363)
[3] Madliger CL, Love OP, Hultine KR, Cooke SJ. 2018 The conservation physiology toolbox: status and opportunities. Conserv Physiol 6, 1–16. (doi:10.1093/conphys/coy029)
[4] Rice DW, Wolman AA. 1971 Life history and ecology of the gray whale (Eschrichtius robustus). Stillwater, Oklahoma: American Society of Mammalogists.
[5] Melicai V, Atkinson S, Calambokidis J, Lang A, Scordino J, Mueter F. 2021 Application of endocrine biomarkers to update information on reproductive physiology in gray whale (Eschrichtius robustus). PLoS One 16. (doi:10.1371/journal.pone.0255368)
[6] Calambokidis J, Darling JD, Deecke V, Gearin P, Gosho M, Megill W, et al. Abundance, range and movements of a feeding aggregation of gray whales (Eschrichtius robustus) from California to south-eastern Alaska in 1998. J Cetacean Res Manag 2002;4:267–76.
[7] Bronson, F. H. (1989). Mammalian reproductive biology. University of Chicago Press.
[8] Wasser SK, Hunt KE, Brown JL, Cooper K, Crockett CM, Bechert U, Millspaugh JJ, Larson S, Monfort SL (2000) A generalized fecal glucocorticoid assay for use in a diverse array of nondomestic mammalian and avian species. Gen Comp Endocrinol120:260–275.
[9] Hunt, K.E., Rolland, R.M., Kraus, S.D., Wasser, S.K., 2006. Analysis of fecal glucocorticoids in the North Atlantic right whale (Eubalaena glacialis). Gen. Comp. Endocrinol. 148, 260–272. https://doi.org/10.1016/j.ygcen.2006.03.01215.
[10] Soledade Lemos L, Burnett JD, Chandler TE, Sumich JL, Torres LG. 2020 Intra‐ and inter‐annual variation in gray whale body condition on a foraging ground. Ecosphere 11. (doi:10.1002/ecs2.3094)
[11] James L. Sumich, James T. Harvey, Juvenile Mortality in Gray Whales (Eschrichtius robustus), Journal of Mammalogy, Volume 67, Issue 1, 25 February 1986, Pages 179–182, https://doi.org/10.2307/1381019
On Saturday, June 10, Dr. Roger Payne passed away. Throughout his remarkable life, he made impactful contributions to the study, understanding, and conservation of whales. His passion, research, and advocacy efforts played a pivotal role in reshaping public perception, and thus promoting the conservation of these giants, profoundly influencing generations of researchers in the field of conservation biology, including myself.
Roger in Patagonia where here found his love for Southern Right Whales. Credit: Dr. Mariano Sironi / ICB.
In 1970, Roger and his first wife Katy Paine began the Southern Right Whale (SRW) Research Program in Patagonia, Argentina, which in 1996 was continued by the Whale Conservation Institute of Argentina (the ICB) , becoming the longest continually running research program on a great whale (based on known individuals) in existence. In this study, Dr. Payne recognized that individual whales can be identified by the unique marks on their heads, establishing an important milestone for photo-ID, a technique that forms the bedrock of whale science.
I am proud to say that I am part of his legacy, as a member of the ICB. With the SRW program, I continued advancing research on SRW through my doctoral dissertation by advancing methods in conservation physiology (see blog post) to understand the underlaying mechanisms affecting young whales’ mortality in Patagonia (see blog post ).
Probably, one of the most remarkable contributions of Dr. Payne to the field and to whale conservation was his groundbreaking discovery of the humpback whale song. In the mid-20th century, the world’s whale populations were intensively killed by commercial whalers, threatening their extinction. In the late 1960s, Payne and his collaborators unveiled the melodic symphonies of humpback whales, marking the start of modern whale biology and catalyzing the global conservationist movement “Save the Whales”. These haunting songs connected humans with these enigmatic animals in an emotional manner, raising public opinion and support for whale conservation that ultimately led to the global moratorium on commercial whaling in 1982.
Listen to this story on NPR featuring Roger Payne’s LP, ‘Songs of the Humpback Whale,’ released in 1970, which played a pivotal role in sparking the global environmental movement “Save the Whales”, helping whale populations on the brink of extinction. Photo: Ocean Alliance.
While he continued to believe that science provides essential information about the necessary changes needed to protect whales, Dr. Payne strongly believe in that the paths to accelerate these changes often involve a combination of activism and creative arts.
“…All of the great movements in human history have been based not on data but on emotion and passion, and a dream of a better society and a better life. For unless people connect emotionally with a problem they won’t connect with the numbers and the data that describe its dimensions…”
“…It seems highly likely that the changes we so desperately need will only come by invoking emotions, and that is something that poets, musicians, writers, playwrights, sculptors, painters, dancers, composers—in fact, creative people of every stripe do well, but that scientists do at their peril. For the real challenge here is to get the world to fall so deeply in love with Nature that we will no longer tolerate the destruction of creation, and will risk our careers and our lives to save all plankton, mosses, ferns, trees, flowers, jellyfish, crinoids, nautiloids, crabs, bees, butterflies, beetles, squid, fishes, frogs, turtles, birds, and mammals—in other words, we will fight to save all of the non-human “Other”…”
Roger Payne’s influence and legacy continue to inspire generations of scientists and conservationists. His work expanded our understanding of whales, deepened our empathy for these creatures, and paved the way for international collaborations aimed at protecting marine life and preserving our oceans. Today, there are many of us who, inspired by Roger, dedicate our lives to research, environmental education, and conservation. And following Roger’s teachings, we constantly ask questions to seek answers that allow us to continue learning about whales in a changing world.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation?
Subscribe to our blog and get a message when we post a new blog. Just add your name and email into the subscribe box below.
Ecological research focuses on understanding how species and ecosystems interact and function, as well as understanding what drives changes in these interactions and functions over time. Thus, ecology is a critical component of conservation biology. Although uncertainty is present in any research, it is a pervasive characteristic of ecology and conservation biology, often due to our inability to control the complexity of natural systems. Uncertainty poses challenges to decision-making, policy development, and effective conservation strategies, and therefore needs to be understood and addressed when conducting ecological studies and conservation efforts.
There are several sources of uncertainty in ecological research and conservation biology. One of the primary sources arises from incomplete or limited data (epistemic uncertainty). Ecological systems are complex, and obtaining comprehensive data on all relevant variables and scales is often challenging or impossible. Data may be lacking or unavailable for certain species, habitats, or regions, which can hinder the ability to fully understand ecological dynamics and make accurate predictions. Additionally, ecological data may be uncertain or variable due to measurement errors (see blog post), sampling biases, or changes in data collection methods over time (Regan et al. 2002). Furthermore, another source of uncertainty arises from language (linguistic uncertainty). Linguistic uncertainty can result from lack of agreement in the terms and definitions used in the scientific vocabulary (see blog post), which can often result in ambiguous, vague, or context dependent interpretations (Regan et al 2002). These two source-types of uncertainty can create a complex set of challenges.
Uncertainty in ecological research and conservation biology has important implications for decision-making and policy development. When faced with uncertain information, decision-makers may adopt a cautious approach, leading to delayed or ineffective conservation actions. Alternatively, they may make decisions based on incomplete or biased data, which can lead to unintended consequences or wasted resources. Uncertainty can also affect the public’s perception of ecological issues, leading to skepticism, misinformation, or lack of support for conservation initiatives. In addition, uncertainty can also pose challenges in setting conservation priorities. With limited resources, conservation organizations and policymakers must prioritize efforts to protect species or habitats that are at the greatest risk. However, uncertainties in data or predictions can affect the accuracy of risk assessments, leading to potential misallocation of resources. Finally, uncertainty may also arise when assessing the success of conservation interventions, making it difficult to determine the effectiveness of the conservation actions.
Despite the challenges posed by uncertainty, there are ways to address and mitigate its impacts in ecological research and conservation biology. Here are some strategies that the GEMM Lab implements to navigate these nuances in ecological research:
Improving data quality and quantity: Robust data can provide a more accurate understanding of ecological dynamics and facilitate evidence-based decision-making. In this direction, the GEMM Lab develops comprehensive data collection and monitoring efforts that can help reduce uncertainty. The TOPAZ and GRANITE projects, which study gray whale ecology off the Oregon coast, are good examples in this direction due to continuous research efforts since 2015. With these projects we have developed and standardized data collection and analytical methods, improved data accuracy and precision, and are filling knowledge gaps through targeted research.
Emphasizing adaptive management: Adaptive management is an approach that involves learning from ongoing conservation actions and adjusting strategies based on new information (Allen et al. 2015). This approach recognizes that uncertainties are inherent in ecological systems and promotes flexibility in conservation planning. Monitoring and evaluating conservation interventions, and adjusting management strategies, accordingly, can help mitigate the impacts of uncertainty. With OBSIDIAN, OPAL, and HALO projects the GEMM Lab works towards a better understanding of cetaceans’ distribution and its interactions with the oceanographic conditions (e.g., ocean temperature). These research projects can help to forecast the occurrence of whale aggregations and inform management to reduce conflicts when overlapping with human activities. For instance, results from the OPAL project have been incorporated into Dungeness Crab fishing regulations to reduce entanglement risk to whales, and the GEMM Lab is now investigating the effectiveness of these regulations in the SLATE project.
With these projects, along with the many other research efforts conducted by the GEMM lab and the MMI, we are advancing research in marine ecology, through the development and application the best possible science to generate the needed ecological data for effective conservation and management of the marine environment.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly message when we post a new blog. Just add your name and email into the subscribe box below.
Sources:
Regan, H. M., Colyvan, M., & Burgman, M. A. (2002). A taxonomy and treatment of uncertainty for ecology and conservation biology. Ecological applications, 12(2), 618-628.
Allen, C. R., & Garmestani, A. S. (2015). Adaptive management (pp. 1-10). Springer Netherlands.
Dr. Alejandro A. Fernández Ajó, Postdoctoral Scholar, Marine Mammal Institute – OSU Department of Fisheries, Wildlife, & Conservation Sciences, Geospatial Ecology of Marine Megafauna (GEMM) Lab.
The English language is recognized as the international language of science (Gordin, 2015); I believe this is a useful convention that allows scientists to communicate ideas and gain access to global scientific literature regardless of their origin or native tongue. However, this avenue for sharing knowledge is open only for those proficient in English, and many scientists and users of scientific information, such as policy makers and conservationists, communicate on a daily basis in languages other than English. This inevitably creates barriers to the transfer of knowledge between communities, potentially impacting conservation and management because scientific knowledge is often unavailable in local languages.
Although in non-English speaking countries, local journals are receptive to publishing scientific research in languages other than English (i.e., their local language), oftentimes these local journals are perceived as low-quality and have a relatively low impact factor, making publishing in such journals less attractive to scientists. Therefore, readers with language barriers only have access to limited studies and are often unaware of the most significant research, even when the research is conducted in their region. This situation can result in a void of information relevant for environmental policies and conservation strategies. Ensuring that research findings are available in the local language of the region in which the research is conducted is an important step in science communication, but one that is often neglected.
In addition, scientists with English as a Foreign Language (EFL) confront the added challenge of navigating a second language while writing manuscripts, preparing and presenting oral presentations, and developing outreach communications (Ramirez-Castaneda, 2020). For example, EFL researchers have reported that one of the primary targets of criticism for their manuscripts under review is often the quality of their English rather than the science itself (Drubin and Kellogg, 2012). In academia, most job interviews and PhD applications are conducted in English; and grant and project proposals are often required to be written in English, which can be particularly challenging and can impact the allocation of resources for research and conservation in non-English speaking regions.
I am from Argentina, and I am a native Spanish speaker. I am fortunate to have started learning English at an early age and continue practicing with international collaborations and traveling abroad. Being able to communicate in English has opened many doors for me, but I recognize that I am in a privileged position with respect to many Argentinians and South Americans in general, where the majority of the population receives minimal training in English and bilingualism with English is very low. Thus, socioeconomic status can influence English proficiency, which then determines scientific success and access to knowledge. I believe that the scientific community should be aware of these issues and work towards improving equality in the process of research collaborations. Providing opportunities for students, and enhancing the availability of scientific knowledge for non-English speaking communities, particularly when the research is relevant for such communities.
In this picture I am with an international group of Fulbright scholars during the Spring International Language Program at the University of Arkansas. This is on of many activities organized by the Fulbright program to create bridges across cultures and languages.
Fortunately, there are several examples pointing towards improving equality in the scientific process, access to knowledge, and opportunities for EFL communities in STEM careers. Several journals are now accepting, or considering to accept the publication of papers in multiple languages. One example of this is the journal Integrative Organismal Biology, which provides the option for publishing the paper abstract in multiple languages. In our recent publication, “Male Bowhead Whale Reproductive Histories Inferred from Baleen Testosterone and Stable Isotopes,” we provided an abstract in five different languages, including Inuktitut, one of primary languages of indigenous groups in the area. And, international exchange programs like the Fulbright Foreign Student Program, of which I was a beneficiary between 2018-2020, enable graduate students and young professionals from abroad to study and conduct research in the United States.
In an effort to contribute to addressing these problems, I am working with a group of colleagues from Argentina (María Constanza (Kata) Marchesi and Tomas Marina) to develop graduate level coursework that will be offered at the Universidad Nacional de la Patagonia in Puerto Madryn, Argentina, with the objective to enable students to learn effective communication using English in the scientific environment. Unfortunately, these types of programs focused on EFL proficiency for STEM students are currently rare in Argentina, but my hope is that our work can spur the creation of additional programs for EFL students in STEM across the region.
I want to finish this post with the acknowledgement of the huge support I have form the GEMM Lab, which welcomes diversity, equity, and inclusivity, and promotes a culture of anti-racism, transparency, and acceptance (See the GEMM Lab DEI statement here).
Did you enjoy this blog? Want to learn more about marine life, research and conservation? Subscribe to our blog and get weekly updates and more! Just add your name into the subscribe box below!
References and Additional Readings
Gordin, M. D. (2015). Scientific Babel : How Science Was Done Before and After Global English. Chicago, IL: University of Chicago Press.
Ramírez-Castañeda V (2020) Disadvantages in preparing and publishing scientific papers caused by the dominance of the English language in science: The case of Colombian researchers in biological sciences. PLoS ONE 15(9): e0238372. https://doi.org/10.1371/journal.pone.0238372
Drubin, D. G., and Kellogg, D. R. (2012). English as the universal language of science: opportunities and challenges. Mol. Biol. Cell 23:1399. doi: 10.1091/mbc.E12-02-0108
Amano, T., González-Varo, J. P., & Sutherland, W. J. (2016). Languages are still a major barrier to global science. PLoS Biology, 14(12), e2000933. https://doi.org/10.1371/journal.pbio.2000933
Marden, E., Abbott, R. J., Austerlitz, F., Ortiz-Barrientos, D., Rieseberg, L. H. (2021). Sharing and reporting benefits from biodiversity research. Molecular Ecology, 30(5), 1103–1107. https://doi.org/10.1111/mec.15702
Márquez, M. C., & Porras, A. M. (2020). Science communication in multiple languages Is critical to Its effectiveness. Frontiers in Communication, 5(May). https://doi.org/10.3389/fcomm.2020.00031
Ramírez-Castañeda V (2020) Disadvantages in preparing and publishing scientific papers caused by the dominance of the English language in science: The case of Colombian researchers in biological sciences. PLoS ONE 15(9): e0238372. https://doi.org/10.1371/journal.pone.0238372
Trisos, C. H., Auerbach, J., & Katti, M. (2021). Decoloniality and anti-oppressive practices for a more ethical ecology. Nature Ecology and Evolution, 5(9), 1205–1212. https://doi.org/10.1038/s41559-021-01460-w
Letter to the Editor of Marine Mammal Science: Enhancing the impact and inclusivity of research by embracing multi-lingual science communication (2022) DOI: 10.13140/RG.2.2.29934.08001 http://dx.doi.org/10.13140/RG.2.2.29934.08001
Leal, J. S., Soares, B., Franco, A. C. S., de Sá Ferreira Lima, R. G., Baker, K., & Griffiths, M. (2022). Decolonizing ecological research: a debate between global North geographers and global South field ecologists. https://doi.org/10.31235/osf.io/wbzh2
Dr. Alejandro A. Fernández Ajó, Postdoctoral Scholar, Marine Mammal Institute – OSU Department of Fisheries, Wildlife, & Conservation Sciences, Geospatial Ecology of Marine Megafauna (GEMM) Lab.
I often receive two reactions when asked what I am currently working on; one is “Wow! That is a very cool job, it must be amazing to work with such incredible animals!”, the other is “How do you do that and why is that important?”. So, today I decided to blog about some of the reasons why it is important to develop a pregnancy test for gray whales and how we are doing this.
In a previous blogpost, I described the many ways in which whales play critical roles in sustaining marine ecosystem. Briefly, whales can enhance marine productivity by vertically and horizontally mixing of ocean waters, promoting primary production, and mitigating climate change by sequestering carbon with their large biomass and long life-span (1-3). Even after they die, their carcasses can contribute to biodiversity creating new habitat on the seafloor (4). But, over several decades, the whaling industry drastically removed whales around the globe, with some species and populations depleted to near extinction (5). Consequently, these depleted whale populations now play a diminished role in ocean ecosystem processes and their recovery is currently challenged by an increasing number of modern anthropogenic impacts. Hence, working towards whale conservation is essential for keeping a healthy marine ecosystem.
Working and designing effective strategies for conservation biology often involves gaining knowledge regarding the reproductive parameters of individual animals in wild populations. This information is critical for understanding population trends and the underlaying mechanisms that affect animal welfare and their potential for recovery. However, getting such information from free-living whales can be challenging (see Hunt et al. 2013). While we know that whales typically have long life-spans, lengthy generation times, extended parental care, and high survival rates, detailed knowledge on the life history and general reproductive biology of free-ranging whales is limited for the majority of the whale populations. In fact, much of what we do know about whale reproduction is derived from whaling records. Only recently, conservation physiology approaches (see our previous post here) have contributed alternative and non-invasive methods for monitoring key physiological processes that can help monitor a whale’s reproductive biology and determine reproductive parameters such as sexual maturity and pregnancy (6-9).
In this clip you can see an example of a fecal sample collection from a gray whale off the Oregon coast. We can look at hormones in the fecal samples which are useful indicators for endocrine assessments of free-swimming whales. Fecal sample and footage filmed under NOAA/NMFS permit #16111.
Gray whales (Eschrichtius robustus) in the Eastern North Pacific (ENP) typically undertake annual migrations between their lower latitude breeding grounds in the coastal waters of the Baja California Peninsula, Mexico, and the foraging grounds located on the Bering and Chukchi Seas (10). However, among the ENP whales a distinct subgroup of about 230 whales shorten their migration to feed in the coastal waters of Northern California, Oregon, and southeastern Alaska (11). This group of whales is known as the gray whale Pacific Coast Feeding Group (PCFG).
Since 2016, the GEMM Lab has monitored individual gray whales within the PCFG off the Oregon coast (check the GRANITE project). Gray whales have a distinct mottled skin; and each individual whale presents a unique pigmentation pattern that allows for the individual identification of whales. We can identify who is who among the whales who visit the Oregon coast. In this way, we can keep a detailed record of re-sightings of known individuals (visit our new web site to know more about the lives of individual whales that visit the Oregon coast). We have high individual re-sighting rates, so this unique opportunity helps us keep a long-term data series for individual whales to monitor their health, body condition, and reproductive status over time, and thus further develop and advance our non-invasive study methods.
We are combining behavioral and feeding ecology with drone photogrammetry and endocrinology of the same individual whales to help us understand the relationships between natural and anthropogenic drivers with biological parameters. In this way, following individual whales, we are developing sensitive biomarkers to monitor and infer about the population health, population trends, and identify stressors that impact their recovery and welfare. In particular, we are now working to develop a noninvasive approach to detect pregnancy in gray whales based on fecal hormone analyses.
In this picture you can see “Rose”, a gray whale calf, on top of her mother “Scarlett”. Scarlett is one of the most recognizable whales from the PCFG, due to a large scar on the right side of her back (not visible in this picture). She has been observed along the Pacific NW coast since 1996, so she is at least 26 years old today. We know 3 of her calves. Following individual whales like Scarlett is helping us to better understand the gray whale reproductive biology. Photo by Alejandro Fernandez Ajo taken under NOAA/NMFS permit #21678.
In marine mammals, the progesterone hormone is secreted in the ovaries during the estrous cycle and gestation, and is the predominant hormone responsible for sustaining pregnancy (12). As the hormones are cleared from the blood into the gut, they are metabolized and eventually excreted in feces; fecal samples represent a cumulative and integrated concentration of hormone metabolites (13-14), which are useful indicators for endocrine assessments of free-swimming whales. Several studies show that changes in hormone concentration correlate in meaningful ways with exposure to stressors (15-16) and changes in reproductive status (17-19). We are using our long data series of fecal hormones and individual life histories to advance our understanding on the gray whales’ reproductive biology. We are close to developing a technique that will allow us to detect pregnancy in whales based in fecal hormones analyses and photogrammetry. Stay tuned for results from this pregnancy test!
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly alert when we make a new post! Just add your name into the subscribe box on the left panel.
References:
1- Pershing AJ, Christensen LB, Record NR, Sherwood GD, Stetson PB (2010) The impact of whaling on the ocean carbon cycle: Why bigger was better. PLoS ONE 5(8): e12444.
2- Roman J and McCarthy JJ. 2010. The whale pump: marine mammals enhance primary productivity in a coastal basin. PLoS ONE. 5(10): e13255.
3- Morissette L, Kaschner K, and Gerber LR. 2010. “Whales eat fish”? Demystifying the myth in the Caribbean marine ecosystem. Fish Fish 11: 388–404.
4- Smith CR, Roman J, Nation JB. A metapopulation model for whale-fall specialists: The largest whales are essential to prevent species extinctions. J. Mar. Res. 77, 283–302 (2019).
5- Branch TA, Williams TM. Legacy of industrial whaling. Whales. Whal. Ocean Ecosyst. 2006, 262–278 (2006).
6- Kellar NM, Keliher J, Trego ML, Catelani KN, Hanns C, George JC, et al. Variation of bowhead whale progesterone concentrations across demographic groups and sample matrices. Endanger Species Res 2013; 22:61–72. https://doi.org/10.3354/esr00537.
7- Pallin L, Robbins J, Kellar N, Berube M, Friedlaender A. Validation of a blubber-based endocrine pregnancy test for humpback whales. Conserv Physiol 2018;6:1 11. https://doi.org/10.1093/conphys/coy031PMID:29942518.
8-Hunt KE, Robbins J, Buck CL, Bérubé M, Rolland RM (2019) Evaluation of fecal hormones for noninvasive research on reproduction and stress in humpback whales (Megaptera novaeangliae). Gen Comp Endocrinol 280: 24–34.
9-Melica, V., Atkinson, S., Calambokidis, J., Lang, A., Scordino, J., & Mueter, F. (2021). Application of endocrine biomarkers to update information on reproductive physiology in gray whale (Eschrichtius robustus). Plos one, 16(8), e0255368.
10-Swartz SL. Gray Whale. In: Wursig B, Thewissen JGM, Kovacs KM, editors. Encyclopedia of Marine Mammals (Third Edition). Elsevier;2018,p. 422–8.https://doi.org/10.1016/B978-0-12-804327-1.00140–0.
11-Calambokidis J, Darling JD, Deecke V, Gearin P, Gosho M, Megill W, et al. Abundance, range and movements of a feeding aggregation of gray whales (Eschrichtius robustus) from California to south-eastern Alaska in 1998. J Cetacean Res Manag 2002;4:267–76.
12- Bronson, F. H. (1989). Mammalian reproductive biology. University of Chicago Press.
13-Wasser SK, Hunt KE, Brown JL, Cooper K, Crockett CM, Bechert U, Millspaugh JJ, Larson S, Monfort SL (2000) A generalized fecal glucocorticoid assay for use in a diverse array of nondomestic mammalian and avian species. Gen Comp Endocrinol120:260–275.
14- Hunt, K.E., Rolland, R.M., Kraus, S.D., Wasser, S.K., 2006. Analysis of fecal glucocorticoids in the North Atlantic right whale (Eubalaena glacialis). Gen. Comp. Endocrinol. 148, 260–272. https://doi.org/10.1016/j.ygcen.2006.03.01215.
15- Lemos, L.S., Olsen, A., Smith, A., Burnett, J.D., Chandler, T.E., Larson, S., Hunt, K.E., Torres, L.G., 2021. Stressed and slim or relaxed and chubby? A simultaneous assessment of gray whale body condition and hormone variability. Mar. Mammal Sci. 1–11. https://doi.org/10.1111/mms.12877
16- Rolland, R., McLellan, W., Moore, M., Harms, C., Burgess, E., Hunt, K., 2017. Fecal glucocorticoids and anthropogenic injury and mortality in North Atlantic right whales Eubalaena glacialis. Endanger. Species Res. 34, 417–429. https://doi.org/10.3354/esr00866.
17-Rolland, R.M., Hunt, K.E., Kraus, S.D., Wasser, S.K., 2005. Assessing reproductive status of right whales (Eubalaena glacialis) using fecal hormone metabolites. Gen. Comp. Endocrinol. 142, 308–317. https://doi.org/10.1016/j.ygcen.2005.02.002
18- Valenzuela Molina M, Atkinson S, Mashburn K, Gendron D, Brownell RL. Fecal steroid hormones reveal reproductive state in female blue whales sampled in the Gulf of California, Mexico. Gen Comp Endocrinol 2018;261:127–35.https://doi.org/10.1016/j.ygcen.2018.02.015 PMID:29476760.
19- Hunt, K. E., Robbins, J., Buck, C. L., Bérubé, M., & Rolland, R. M. (2019). Evaluation of fecal hormones for noninvasive research on reproduction and stress in humpback whales (Megaptera novaeangliae). General and Comparative Endocrinology, 280, 24-34.
Dr. Alejandro A. Fernández Ajó, Postdoctoral Scholar, Marine Mammal Institute – OSU Department of Fisheries, Wildlife, & Conservation Sciences, Geospatial Ecology of Marine Megafauna (GEMM) Lab.
I just returned to my home country, Argentina, after over 2 years without leaving the USA due to COVID-19 travel restrictions. Being back with my family, my friends, my culture, and speaking my native language feels great and relaxing. However, I returned to a country struggling to rebound from the coronavirus pandemic. I am afraid this post pandemic scenario places Argentina in a vulnerable situation. The need for economic growth could result in decisions or policies that, in the long term, hurt the country, leaving environmental damage for potential economic growth.
Argentina holds extensive oil and gas deposits, including the world’s second largest gas formation, Vaca Muerta. Although offshore (i.e., in the ocean) oil exploration and exploitation are not yet extensively developed, the intention of offshore gas and oil drilling is on the agenda. In July 2021, a public hearing was held with the purpose to consider the environmental impact assessment for carrying out seismic exploration in the North Argentinian basin off the southern coast of the Buenos Aires province. Over 90% of the participants, including scientists, researchers, technicians from various institutions, non-governmental organizations and representatives of the fishing sector spoke against the project and highlighted the negative impacts that such activity can generate on marine life, and to other socioeconomic activities such as tourism and fishing, not only in Argentina but at the regional level.
Thousands of people marched along the beaches and the main coastal cities of Argentina to protest against the approval for a seismic explorations project in the Argentinian basin. Photo source: prensaobrera.com
Seismic prospections are usually done with the purpose for oil and gas exploitation and less frequently for research purposes. In seismic prospections, ships carry out explosions with airguns, whose sound waves reach the seabed, bounce back and are captured by receivers on the ships to map the petroleum deposits in seafloor and identify potential areas for hydrocarbon extractions. The sound emitted by the seismic airguns can reach extremely loud levels of sounds that travel for thousands of miles underwater. Such extreme high levels of sound can alter the behavior of many marine species, from the smallest planktonic species, to the largest marine mammals, masking their communication, causing physical and physiological stress, interfering with their vital functions, and reducing the local availability of prey (Di Iorio & Clark, 2010; Hildebrand, 2009; Weilgart, 2018).
Here you can listen to a short audio clip of a seismic airgun firing every ~8 seconds, a typical pattern. Close your eyes and imagine you are a whale living in this environment. Now, put the clip on loop and play it for three months straight. This would be the soundscape that whales living in a region of oil and gas exploration hear, as seismic surveys often last 1-4 months (see our previous post “Hearing is believing” for more details).
Despite the public rejection and the mounting evidence about the negative impacts and environmental risks associated with such activities, the government approved the initiation of the seismic prospection off the southern coast of Buenos Aires. In response, thousands of people marched along the beaches and the main coastal cities of Argentina to protest against the oil exploration project. The areas where the seismic surveys will be carried out overlap largely with the southern right whale’s migration routes and feeding areas during their spring and summer (Figure 1). Likewise, the area overlaps with highly productive areas in the ocean that hosts great biodiversity of species of ecological and commercial importance, including the feeding areas of seabirds, turtles and other marine mammals. Additionally, the seismic activity will endanger the health of the beaches of the coast of Buenos Aires and Uruguay where thousands of tourists spend the summer to escape from the large cities.
Figure 1. The map on the left is showing (light blue squares CAN_100, CAN_108, and CAN_114) the areas where seismic prospections are proposes. The map on the right is showing the individual satellite track lines for eleven individual southern right whales (SRW) during the feeding season. You can observe that the proposed area for seismic explorations overlaps with critical feeding habitat for the SRW. Source: Whale Conservation Institute of Argentina (ICB).
The impacts of these activities to marine wildlife are difficult to control and monitor (Elliott et al. 2019, Gordon et al, 2003), especially for large whales that are a very challenging taxa to study (Hunt et al. 2013). We know that the ability to perceive biologically important sounds is critical to marine mammals, and acoustic disturbance through human-generated noise can interfere with their natural functions. Sounds from seismic surveys are intense and have peak frequency bands overlapping those used by baleen whales (Di Lorio & Clark, 2010); however, evidence of interference with baleen whale acoustic communication, and the effects on their health and physiology are sparse. In this context, the GEMM Lab project GRANITE (Gray Whale Response to Ambient Noise Informed by Technology and Ecology), plans to generate information to fulfill these knowledge gaps and provide tools to aid conservation and management decisions in terms of allowable noise level in whale habitats. I am hopeful such information will reach decision makers and influence their decisions, however, sometimes it is frustrating to see how evidence-based information generated with high quality standards are often ignored.
The recent approval of the seismic exploration in Argentina is an example of my frustration. There is no way that the oil industry can guarantee a low-risk of impact on biodiversity and the environment. There are too many examples of environmental catastrophes related to the oil industries at sea that speak for themselves. Moreover, the promotion of such activities goes against the compromises assumed by the country to work to mitigate the effects of Climate Change, and to achieve the reductions of the greenhouse gas emissions to comply with the Paris Agreement. Decades of research help recognized the areas that would be impacted by these seismic activities as key habitat for the life cycle of whales, penguins, seals and more. But, apparently all these scientific data are ignored at the time of weighing the tradeoffs between “economic development” and environmental impacts. As a conservation biologist, I am questioning what can be done in order to be heard and significantly influence such decisions.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly alert when we make a new post! Just add your name into the subscribe box on the left panel.
References:
Di Iorio, L., & Clark, C. W. (2010). Exposure to seismic survey alters blue whale acoustic communication. Biology Letters, 6(1), 51–54. https://doi.org/10.1098/rsbl.2009.0651
Weilgart, L. (2018). The impact of ocean noise pollution on fish and invertebrates. Report for OceanCare, Switzerland.
Elliott, B. W., Read, A. J., Godley, B. J., Nelms, S. E., & Nowacek, D. P. (2019). Critical information gaps remain in understanding impacts of industrial seismic surveys on marine vertebrates. In Endangered Species Research (Vol. 39, pp. 247–254). Inter-Research. https://doi.org/10.3354/esr00968
Gordon, J., Gillespie, D., Potter, J., Frantzis, A., Simmonds, M. P., Swift, R., & Thompson, D. (2003). A review of the effects of seismic surveys on marine mammals. Marine Technology Society Journal, 37(4), 16-34.
Hunt, K. E., Moore, M. J., Rolland, R. M., Kellar, N. M., Hall, A. J., Kershaw, J., Raverty, S. A., Davis, C. E., Yeates, L. C., Fauquier, D. A., Rowles, T. K., & Kraus, S. D. (2013). Overcoming the challenges of studying conservation physiology in large whales: a review of available methods. Conservation Physiology, cot006–cot006. https://doi.org/10.1093/conphys/cot006
Dr. Alejandro Fernández Ajó, OSU Department of Fisheries, Wildlife, & Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
Rises in ocean temperatures can lead to multiple alterations in marine ecosystems, including the increase and the frequency of Harmful Algal Blooms (HABs). HABs are characterized by the rapid growth of toxin-producing species of algae that can be harmful to people, animals, and the local ecology, even causing death in severe cases. Species of marine diatom within the genus Pseudo-nitzschia and Nitzschia can form HABs when they produce domoic acid (DA), a potent neurotoxin responsible for amnesic shellfish poisoning (D’Agostino et al., 2018, 2017).
Figure 1. Southern right whale (E. australis) mother and calf swimming at the gulfs of Peninsula Valdes, Argentina, during a phytoplankton bloom. Photo: Mariano Sironi / Instituto de Conservacion de Ballenas de Argentina.
During HABs, DA is transferred to higher organisms through the pelagic food web and is accumulated by intermediate vectors, such as copepods, euphausiids (i.e., krill), shellfish, and fish. As this neurotoxin affects top predators, DA poisoning poses a risk to the safety and health of humans and wildlife. This neurotoxin has caused mortality in many marine mammal species, including both pinnipeds and cetaceans (Gulland 1999; Lefebvre et al. 1999; Fire et al. 2010, 2021; Broadwater et al. 2018). In addition, the exposure to DA constitutes a stressor that may affect glucocorticoids (hormones involved in the stress response) concentrations.
The glucocorticoids (GCs; cortisol and corticosterone) are adrenal steroid hormones that maintain the essential functions of metabolism and energy balance in mammals. GCs can increase sharply in response to environmental stressors to elicit physiological and behavioral adaptations by individuals to support survival (Sapolsky et al. 2000; Bornier et al. 2009). However, with the chronic exposure to a stressor, this relationship can reverse, with GCs sometimes declining below its baseline levels (Dickens and Romero, 2013; Fernández Ajó et al., 2018). Moreover, DA can interfere with the stress response in mammals, and cause alterations in their physiological response. DA is an excitatory amino acid analog of glutamate (Pulido 2008), a well-known brain neurotransmitter that play an important role in the activation of the adrenal axis (which in turn regulate the production and secretion of the GCs) and regulate many of the pituitary hormones involved in the stress response (Brann and Mahesh 1994; Johnson et al. 2001). Hence, monitoring GC levels in marine mammals can be a potential useful metric for assessing the physiological impacts of exposure to DA.
Glucocorticoids are traditionally measured in plasma, but given that plasma sampling from free-ranging large whales is currently impossible, alternative sample types such as fecal samples, among others, can be utilized to quantify GCs in large whales (Ajó et al., 2021; Burgess et al., 2018, 2016; Fernández Ajó et al., 2020, 2018; Hunt et al., 2019, 2014, 2006; Rolland et al., 2017, 2005)(Figure 2). The analyses of fecal glucocorticoid metabolites (fGCm) is particularly useful for endocrine assessments of free-swimming whales, with several studies showing that fGCm correlate in meaningful ways with presumed stressors. For example, high levels of fGCm in North Atlantic right whales (NARW, Eubalaena glacialis) and in gray whales (Eschrichtius robustus) correlate with poor body condition (Hunt et al., 2006; Lemos et al., 2021), and fGCm increases were associated with whale entanglements and ship strikes (i.e., Lemos et al., 2020; Rolland et al., 2017).
Figure 2. Alternative samples types can be used to study hormones in large whales. 1-2-3 are sample types that can be obtained from free-living whales and provide a more instantaneous and acute measurement of the whales´ physiology. 4-5 can be obtained at necropsy when the whale is found dead at the beach and provide an integrated measure of the whale physiology that can expand through years or even the lifespan of an individual.
In Península Valdés, Argentina, southern right whales (SRW, E. australis) gather in large numbers to mate and nurse their calves during the austral winter months (Bastida and Rodríguez, 2009). SRWs are capital breeders, largely fasting during the breeding season and instead relying on stored blubber fuel reserves. However, they can occasionally feed on calanoid copepods (D’Agostino et al., 2018, 2016), particularly during the phytoplankton blooms that are dominated by diatoms of the genus Pseudo-nitzschia (Sastre et al. 2007; D’Agostino et al. 2015, 2018). Therefore, feeding SRWs in Península Valdés temporally overlap with these Pseudo-nitzschia blooms (D’Agostino et al. 2018, 2015) and represents a test case for assessing the relationship of DA exposure with GC levels (Figure 3).
Figure 3. Southern right whale (E. australis) skim feeding at the Peninsula Valdes breeding ground. Photo: Lucas Beltranino.
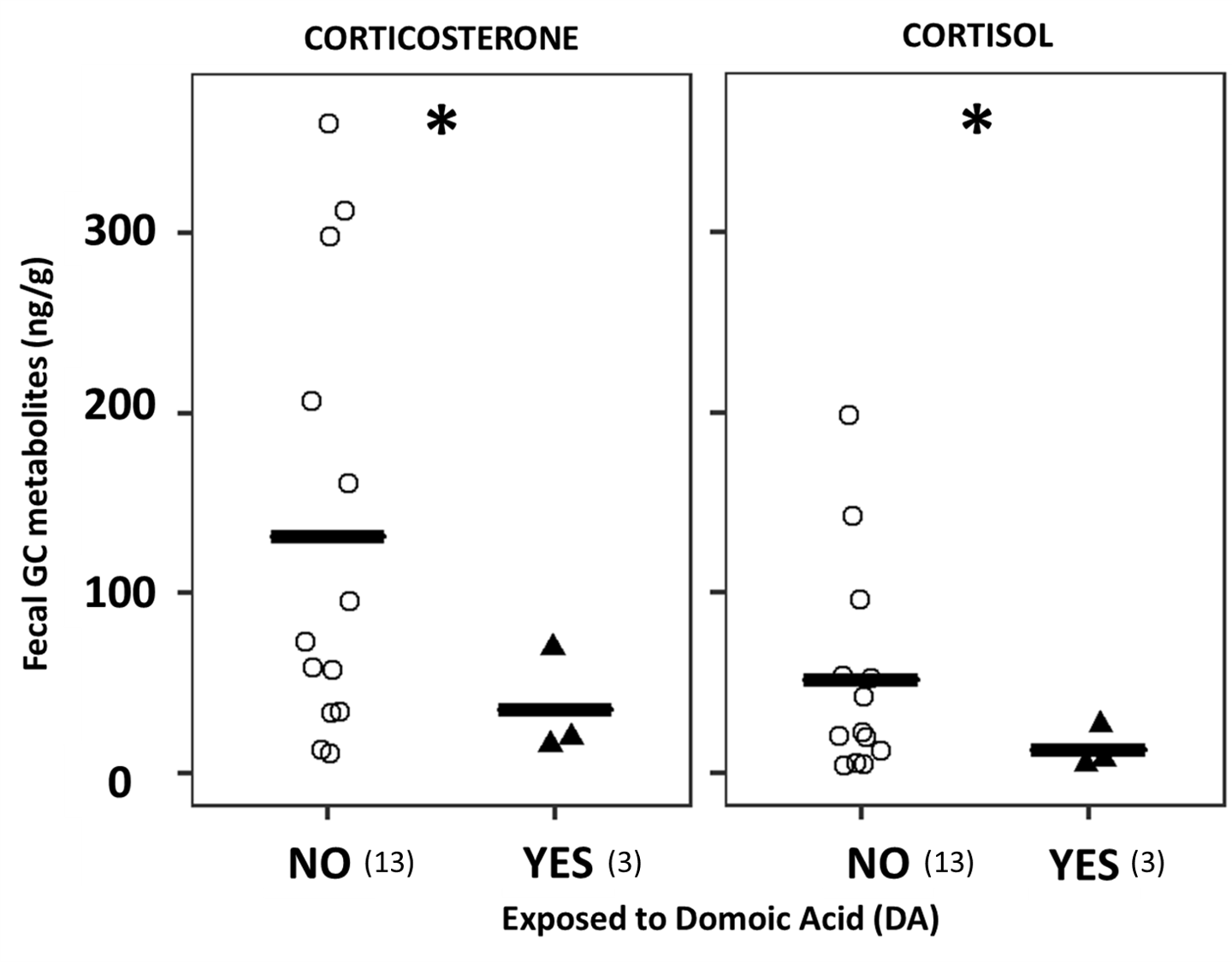
In our recent scientific publication (D’Agostino et al. 2021), we investigate SRW exposure to DA at their breeding ground in Peninsula Valdes and assessed its effects on fecal glucocorticoid concentrations. Although the sample size of this study is unavoidably small due to the difficulties of obtaining fecal samples from whales at their calving grounds where defecation is infrequent, we observed significantly lower fGCm in samples from whales exposed to DA (Figure 4). Our results agree with findings from a previous study in California sea lions (Zalophus californianus) exposed to DA, where these authors found a significant association of DA exposure with reduced serum cortisol (Gulland et al., 2009), which can be tentatively attributed to abnormal function of the adrenal axis due to the exposure.
Figure 4. Fecal glucocorticoid metabolite levels in southern right whales exposed (YES, solid triangles) and not-exposed (NO, open circles) to DA. Left panel: immunoreactive fecal corticosterone metabolites. Right panel: immunoreactive fecal cortisol metabolites. Hormone concentrations are expressed in ng of immunoreactive hormone per gram of dry fecal sample. Significant differences between groups are denoted with an asterisk (P<0.05). The black solid line indicates the mean for each group, and in parenthesis is the sample size for each group. Adapted from D’Agostino et al. 2021.
If ingestion of toxins produced by phytoplankton can result in long-term suppression of baseline GCs, whales and marine mammals in general, could suffer reduced ability to cope with additional stressors. The adrenal function is essential to maintain circulating blood glucose and other aspects of metabolism within normal bounds. Additionally, the ability to elevate GCs facilitates energy mobilization to physiologically cope with a stressful event and to initiate appropriate behavioral responses (i.e., flee from predators, heal wounds). Various toxicants have been shown to reduce adrenal function across taxa (Romero and Wingfield, 2016) and could have negative consequences on the ability of cetaceans to respond and adapt to ongoing environmental and anthropogenic changes. Compounding this problem, whales are exposed to an increasing number of stressors from multiple sources and with cumulative effects and they need to be able to physiologically respond to continue to reproduce and survive.
To our knowledge, this study provides the first quantification of fGCm levels in whales exposed to DA; and we hope this effort starts a growing dataset to which other researchers can add. Sampling and analysis of non-traditional matrices, such as feces, blubber, baleen and others, would likely increase sample sizes and thus our understanding of the interrelationships among DA exposure and age, sex, and reproductive status of cetaceans. Given that chronic exposure to DA could alter the capacity of animals to respond to stress, and indications that HABs are becoming more frequent and intense world-wide (Van Dolah 2000; Masó et al. 2006; Erdner et al. 2008), we believe that research evaluating the health status of marine mammal populations should include the assessment of stress physiology relative to natural and anthropogenic stressors including exposure to toxicants.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get weekly updates and more! Just add your name into the subscribe box on the left panel.
References
Bastida R, Rodríguez D (2009) Ballena franca austral [Southern right whale]. In: Mazzini V (ed) Mamíferos marinos de Patagonia y Antártida [Marine mammals of Patagonia and Antarctica]. Zagier & Urruty Publications, Buenos Aires, pp 72–84.
Bonier F, Moore IT, Martin PR, Robertson RJ (2009) The relationship between fitness and baseline glucocorticoids in a passerine bird. Gen Comp Endocrinol 163:208–213. https:// doi. org/ 10. 1016/j.ygcen. 2008. 12. 013.
Brann DW, Mahesh VB (1994) Excitatory amino acids: function and significance in reproduction and neuroendocrine regulation. Front Neuroendocrinol 15:3–49. https:// doi. org/ 10. 1006/ frne. 1994. 1002.
Broadwater MH, Van Dolah FM, Fire SE (2018) Vulnerabilities of marine mammals to harmful algal blooms. Harmful Algal Blooms 2:191–222.
Burgess, E.A., Hunt, K.E., Kraus, S.D., Rolland, R.M., 2016. Get the most out of blow hormones: Validation of sampling materials, field storage and extraction techniques for whale respiratory vapour samples. Conserv. Physiol. 4, cow024. https://doi.org/10.1093/conphys/cow024
Burgess, E.A., Hunt, K.E., Kraus, S.D., Rolland, R.M., 2018. Quantifying hormones in exhaled breath for physiological assessment of large whales at sea. Sci. Rep. 8, 10031. https://doi.org/10.1038/s41598-018-28200-8
D’Agostino VC, Hoffmeyer MS, Almandoz GO, Sastre V, Degrati M., 2015. Potentially toxic Pseudo-nitzschia species in plankton and fecal samples of Eubalaena australis from Península Valdés calv-ing ground, Argentina. J Sea Res 106:39–43. https:// doi. org/ 10.1016/j. seares.
D’Agostino, V.C., Degrati, M., Santinelli, N., Sastre, V., Dans, S.L., Hoffmeyer, M.S., 2018. The seasonal dynamics of plankton communities relative to the foraging of the southern right whale (Eubalaena australis) in northern Patagonian gulfs, Península Valdés, Argentina. Cont. Shelf Res. 164, 45–57. https://doi.org/10.1016/j.csr.2018.06.003
D’Agostino, V.C., Degrati, M., Sastre, V., Santinelli, N., Krock, B., Krohn, T., Dans, S.L., Hoffmeyer, M.S., 2017. Domoic acid in a marine pelagic food web: Exposure of southern right whales Eubalaena australis to domoic acid on the Península Valdés calving ground, Argentina. Harmful Algae 68, 248–257. https://doi.org/10.1016/j.hal.2017.09.001
D’Agostino, V.C., Hoffmeyer, M.S., Degrati, M., 2016. Faecal analysis of southern right whales (Eubalaena australis) in Península Valdés calving ground, Argentina: Calanus australis, a key prey species. J. Mar. Biol. Assoc. United Kingdom 96, 859–868. https://doi.org/10.1017/S0025315415001897
Dickens, M.J., Romero, L.M., 2013. A consensus endocrine profile for chronically stressed wild animals does not exist. Gen. Comp. Endocrinol. 191, 177–189. https://doi.org/10.1016/j.ygcen.2013.06.014
Erdner DL, Dyble J, Parsons ML, Stevens RC, Hubbard KA, Wrabel ML, Moore SK, Lefebvre KA, Anderson DM, Bienfang P, Bidi-gare RR, Parker MS, Moeller P, Brand LE, Trainer VL (2008) Centers for Oceans and Human Health: a unified approach to the challenge of harmful algal blooms. Environ Health 7:S2. https://doi. org/ 10. 1186/ 1476- 069X-7- S2- S2.
Fernández Ajó, A., Hunt, K.E., Dillon, D., Uhart, M., Sironi, M., Rowntree, V., Buck, C.L., 2021. Optimizing hormone extraction protocols for whale baleen: tackling questions of solvent:sample ratio and variation. Gen. Comp. Endocrinol. 113828. https://doi.org/10.1016/j.ygcen.2021.113828
Fernández Ajó, A.A., Hunt, K.E., Giese, A.C., Sironi, M., Uhart, M., Rowntree, V.J., Marón, C.F., Dillon, D., DiMartino, M., Buck, C.L., 2020. Retrospective analysis of the lifetime endocrine response of southern right whale calves to gull wounding and harassment: A baleen hormone approach. Gen. Comp. Endocrinol. 296, 113536. https://doi.org/10.1016/j.ygcen.2020.113536
Fernández Ajó, A.A., Hunt, K.E., Uhart, M., Rowntree, V., Sironi, M., Marón, C.F., Di Martino, M., Buck, C.L., 2018. Lifetime glucocorticoid profiles in baleen of right whale calves: potential relationships to chronic stress of repeated wounding by Kelp Gulls. Conserv. Physiol. 6, 1–12. https://doi.org/10.1093/conphys/coy045
Fire SE, Bogomolni A, DiGiovanni RA Jr, Early G, Leighfield TA, Matassa K, Miller GA, Moore KM, Moore M, Niemeyer M, Pugliares K (2021) An assessment of temporal, spatial and taxonomic trends in harmful algal toxin exposure in stranded marine mammals from the US New England coast. PLoS ONE 16(1):e0243570. https:// doi. org/ 10. 1371/ journ al. pone. 02435 70
Fire SE, Wang Z, Berman M, Langlois GW, Morton SL, Sekula-Wood E, Benitez-Nelson CR (2010) Trophic transfer of the harmful algal toxin domoic acid as a cause of death in a minke whale (Balaenoptera acutorostrata) stranding in southern California. Aquat Mamm 36(4):342–350. https:// doi. org/ 10. 1578/ AM. 36.4.2010. 342.
Gulland F.M., 1999. Domoic acid toxicity in California sea lions stranded along the central California Coast, May-October 1998. NOAA Tech. Memo. NMFS-OPR-8. USA National Marine Fisheries Service, US Department of Commerce.
Gulland FMD, Hall AJ, Greig DJ, Fram ER, Colegrove KM, Booth RKN, Wasser SK, S.-M.C., 2009. Gulland, Hall – 2012 – Evaluation of circulating eosinophil count and adrenal gland function in California sea lions naturally exposed t. J. Am. Vet. Med. Assoc. 241, 943–949.
Gulland, F., Pérez-Cortés, H., Urbán, J.R., Rojas-Bracho, L., Ylitalo, G., Weir, J., Norman, S., Muto, M., Rugh, D., Kreuder, C., Rowles, T., 2005. Eastern North Pacific gray whale (Eschrichtius robustus) unusual mortality event, 1999-2000. U.S. Dep. Commer. NOAA Tech. Memo. NMFS-AFSC-150. 33 pp.
Hunt, K.E., Robbins, J., Buck, C.L., Bérubé, M., Rolland, R.M., 2019. Evaluation of fecal hormones for noninvasive research on reproduction and stress in humpback whales (Megaptera novaeangliae). Gen. Comp. Endocrinol. 280, 24–34. https://doi.org/10.1016/j.ygcen.2019.04.004
Hunt, K.E., Rolland, R.M., Kraus, S.D., Wasser, S.K., 2006. Analysis of fecal glucocorticoids in the North Atlantic right whale (Eubalaena glacialis). Gen. Comp. Endocrinol. 148, 260–272. https://doi.org/10.1016/j.ygcen.2006.03.012
Hunt, K.E., Stimmelmayr, R., George, C., Hanns, C., Suydam, R., Brower, H., Rolland, R.M., 2014. Baleen hormones: a novel tool for retrospective assessment of stress and reproduction in bowhead whales (Balaena mysticetus). Conserv. Physiol. 2, cou030–cou030. https://doi.org/10.1093/conphys/cou030
Johnson MP, Kelly G, Chamberlain M (2001) Changes in rat serum corticosterone after treatment with metabotropic glutamate receptor agonists or antagonists. J Neuroendocrinol 13:670–677. https:// doi. org/ 10. 1046/j. 1365- 2826. 2001. 00678.x.
Lefebvre KA, Powell CL, Busman M, Doucette GJ, Moeller PDR, Sliver JB, Miller PE, Hughes MP, Singaram S, Silver MW, Tjeer-dema RS (1999) Detection of domoic acid in northern anchovies and California sea lions associated with an unusual mortality event. Nat Toxins 7(3):85–92. https:// doi. org/ 10. 1002/ (SICI) 1522-7189(199905/ 06)7: 3% 3C85:: AID- NT39% 3E3.0. CO;2-Q.
Lefebvre, K.A., Kendrick, P.S., Ladiges, W., Hiolski, E.M., Ferriss, B.E., Smith, D.R., Marcinek, D.J., 2017. Chronic low-level exposure to the common seafood toxin domoic acid causes cognitive deficits in mice. Harmful Algae 64, 20–29. https://doi.org/10.1016/j.hal.2017.03.003
Lemos, L.S., Olsen, A., Smith, A., Burnett, J.D., Chandler, T.E., Larson, S., Hunt, K.E., Torres, L.G., 2021. Stressed and slim or relaxed and chubby? A simultaneous assessment of gray whale body condition and hormone variability. Mar. Mammal Sci. 1–11. https://doi.org/10.1111/mms.12877
Lemos, L.S., Olsen, A., Smith, A., Chandler, T.E., Larson, S., Hunt, K., Torres, L.G., 2020. Assessment of fecal steroid and thyroid hormone metabolites in eastern North Pacific gray whales. Conserv. Physiol. 8. https://doi.org/10.1093/conphys/coaa110
Masó M, Garcés E., 2006. Harmful microalgae blooms (HAB); prob-lematic and conditions that induce them. Mar Pollut Bull 53:620–630. https:// doi. org/ 10. 1016/j. marpo lbul. 2006. 08. 006.
Pulido O.M., 2008. Domoic acid toxicologic pathology: a review. Mar Drugs 6(2):180–219. https:// doi. org/ 10. 3390/ md602 0180.
Rolland, R., McLellan, W., Moore, M., Harms, C., Burgess, E., Hunt, K., 2017. Fecal glucocorticoids and anthropogenic injury and mortality in North Atlantic right whales Eubalaena glacialis. Endanger. Species Res. 34, 417–429. https://doi.org/10.3354/esr00866
Rolland, R.M., Hunt, K.E., Kraus, S.D., Wasser, S.K., 2005. Assessing reproductive status of right whales (Eubalaena glacialis) using fecal hormone metabolites. Gen. Comp. Endocrinol. 142, 308–317. https://doi.org/10.1016/j.ygcen.2005.02.002
Romero, M.L., Wingfield, J.C., 2016. Oxford series in behavioral neuroendocrinology. Tempests, poxes, predators, and people: stress in wild animals and how they cope 1–2.
Sastre V, Santinelli N, Marino G, Solís M, Pujato L, Ferrario M., 2007. First detection of domoic acid produced by Pseudo-nitzschia spe-cies, Chubut coastal waters, Patagonia, Argentina. Harmful Algae News 34:12–14.
Van Dolah FM., 2000. Marine algal toxins: origins, health effects, and their increased occurrence. Environ Health Perspect 108:133–141. https:// doi. org/ 10. 1289/ ehp. 00108 s1133