Dawson Mohney, TOPAZ/JASPER HS Intern, Pacific High School Graduate
My name is Dawson Mohney, I am a high school intern for the 2025 TOPAZ/JASPER team this field season. I first heard about the TOPAZ/JASPER internship from my friend Jonah Lewis, a previous intern from the 2023 field season. Coincidentally, Jonah and I both graduated this year from Pacific High School here on the coast—small world. I have called Port Orford my home for most of my life, and in recent years I discovered that a gray whale research project has been happening in my own backyard. Growing up less than a mile from the Oregon Coast, I’ve spent a lot of time looking out into the water. I always liked how, no matter what happened in my life, the ocean was always there. This interest is what encouraged me to apply for the internship with the hope of discovering more about the ocean, a substantial part of my home and family.
Fig 1: Picture fellow intern Maddie took of me (Dawson) during our trip to Natural Bridges.
A critical part of this project is understanding not only the magnificent gray whales but also the much less apparent zooplankton–after all, the whales need to eat a lot of zooplankton! Many different species of zooplankton—“zoop” for short—call the Oregon coast home. Each day, as we kayak to our 12 sample stations within the gray whale feeding grounds of Mill Rocks and Tichenor’s Cove, I find myself wondering which species of zoop I’ll get to identify later under the microscope.
Throughout the duration of this internship, our team has met to discuss a few research papers published by GEMM Lab members, including research produced from the TOPAZ/JASPER projects. Recently, I read, “Do Gray Whales Count Calories? Comparing Energetic Values of Gray Whale Prey Across Two Different Feeding Grounds in the Eastern North Pacific,” by Hildebrand et al. who describe the caloric content of different zooplankton species. Before reading this paper, I didn’t realize whale prey could vary in nutritional value – much like food for humans. This paper made it clear that each of the different species of zooplankton is just as important as the last, but consuming more of the higher caloric species such as the Neomysis rayii or the Dungeness crab larvae would certainly be a welcome meal. Seeing these “healthy” meals in the area makes me hopeful for the whales.
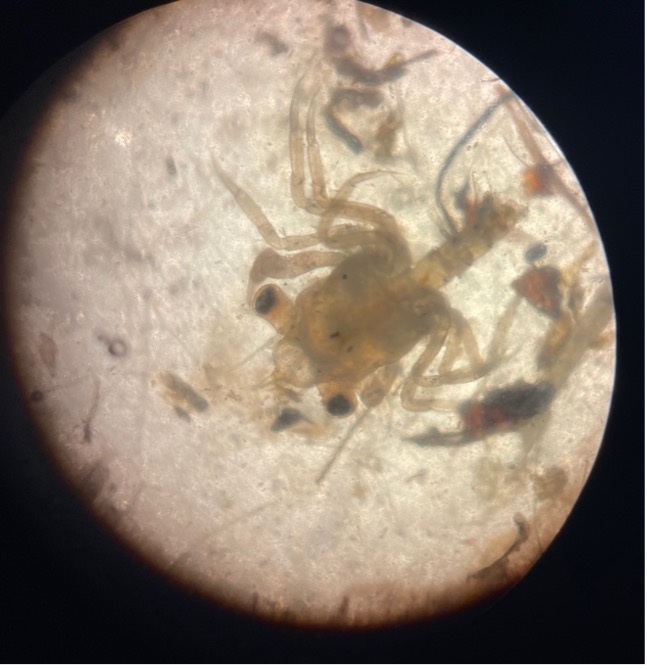
Fig 2: Image of a crab larvae in their megalopae stage.
From reading previous blog posts, the foraging habits of the whales this season appear to be unusual. In prior TOPAZ/JASPER field seasons, gray whales have often been tracked foraging near or around our Mill Rocks and Tichenor Cove study sites. This season, we haven’t tracked a single whale in Mill Rocks and only two in Tichenor Cove. Could there just not be enough good zoop?
Along with this lack of whales, there does seem to be a lack of these “high calorie zoop species”. Our team has most frequently collected samples primarily comprising of Atylus tridens, a lower calorie prey type. In fact, during one of our earlier kayak training days this field season we collected 2,019 individual A. tridens. However, since this day we have collected sparse amounts of zooplankton in our samples, ranging from zero to 121 in a given sample. Our total zoop count thus far is 2,524 zooplankton, a third of the total zooplankton collected last field season.
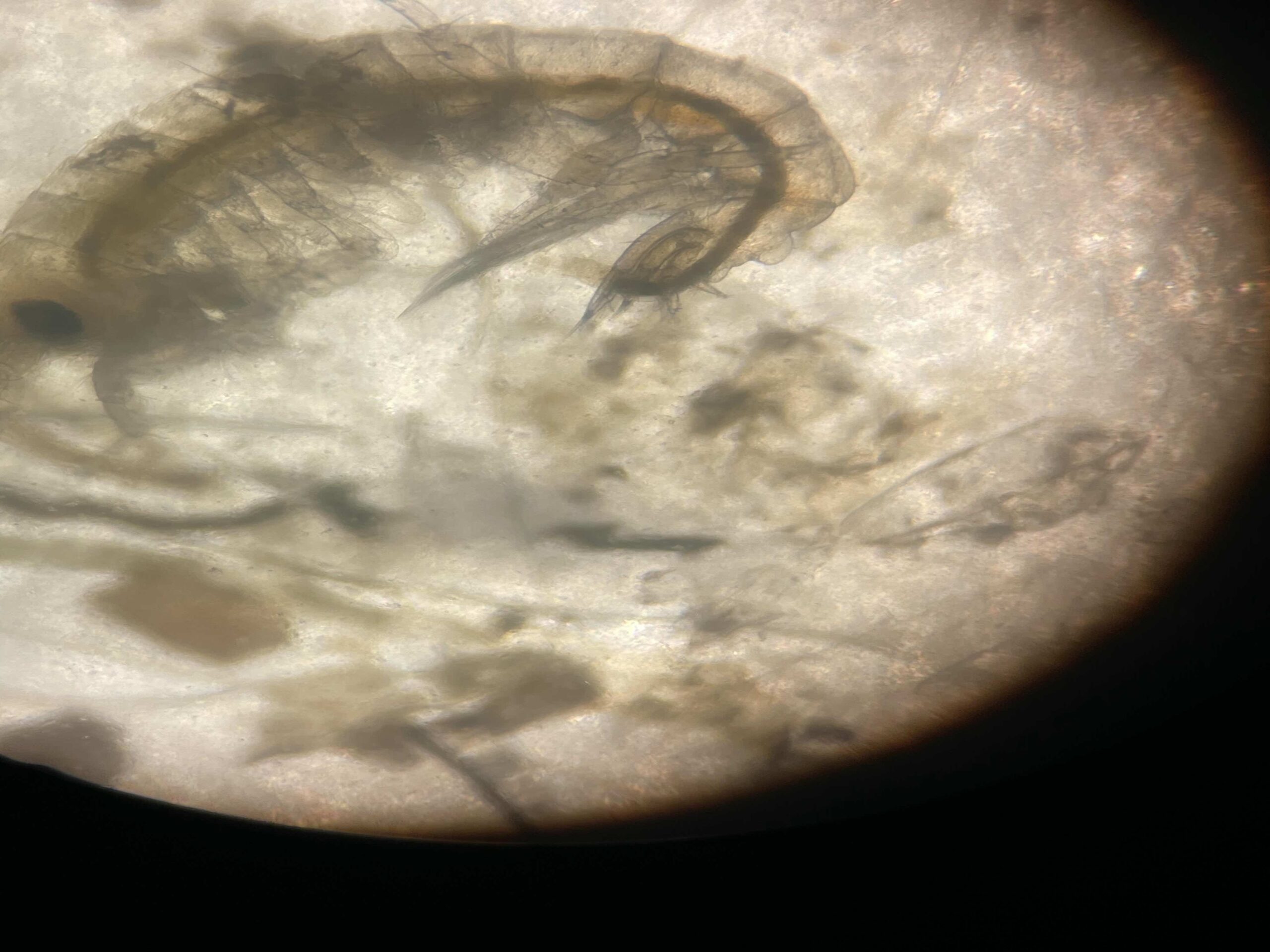
Fig 3: Image of an Atylus tridens under a microscope.
As for whale presence, we have been observing many whales blows near Hell’s Gate as mentioned in last week’s blog written by fellow intern Miranda Fowles. From our cliff site, it has been difficult to know whether these are gray whales or a different kind of whale, leading us to venture out to the Heads to get a better look. The persistence of whales in this area is certainly unusual, and perhaps it can be explained by a larger amount of higher calorie zooplankton species in the Hell’s Gate area.
Fig 4: Dawson tracking blows by Hell’s Gate with the theodolite.
Being part of the TOPAZ/JASPER project, I have become exposed to what the true meaning is behind “fieldwork,” including learning how to be flexible and adapt to new challenges every day. What I have most enjoyed is the team’s ability to overcome any new hurdle together as a unit. My dad often says, “You learn something new every day,” and this internship couldn’t embody this quote more. In just these 5 weeks, it almost feels like my head is now a couple sizes bigger.
Before this experience, I never thought much about how one might track a whale or how different microscopic species could have such a profound impact on a whale’s decision to forage. Now I feel I understand just how important these less than obvious factors are and the effort which goes behind understanding these relationships. I can only hope future opportunities teach me as much as joining the TOPAZ/JASPER legacy has—it’s an experience that, even just a few days into the 2025 field season, I knew would be hard to match.
Fig 4: Dawson (navigator) and Miranda (sampler) during kayak training on their way to Mill Rocks.
Hildebrand, L., Bernard, K. S., & Torres, L. G. (2021). Do Gray Whales Count Calories? Comparing Energetic Values of Gray Whale Prey Across Two Different Feeding Grounds in the Eastern North Pacific. Frontiers in Marine Science, 8, 683634. https://doi.org/10.3389/fmars.2021.683634
Miranda Fowles, GEMM Lab TOPAZ/JASPER Intern, OSU Fisheries and Wildlife Undergraduate
Hello! My name is Miranda Fowles, and I am the OSU intern for the 2025 TOPAZ/JASPER project this summer! I recently earned my bachelor’s degree – almost, I have one more term, but I walked at commencement in June – from Oregon State University in Fisheries, Wildlife and Conservation Sciences and a minor in Spanish. My interest in whales began at a young age during a visit to SeaWorld. While I didn’t enjoy the killer whale shows for their entertainment aspect, this exposure allowed me to see a whale for the first time. From then on, I knew I wanted to contribute to understanding more about these animals, even if I wasn’t always sure how to make that happen. My decision to pursue Fisheries and Wildlife sciences was set from the beginning, however I wondered if there were actually opportunities to study whales.
Last summer, I was a MACO intern and stayed at the Hatfield Marine Science Center where I met last year’s TOPAZ/JASPER REU student, Sophia Kormann, and she raved all about her experience, so I just had to apply for this year’s internship! I remember feeling so nervous for the interview, but Dr. Leigh Torres and Celest Sorrentino’s kindness and inspiration quickly put me to ease. When I found out I was offered the position, I was just more excited than I’d ever been!
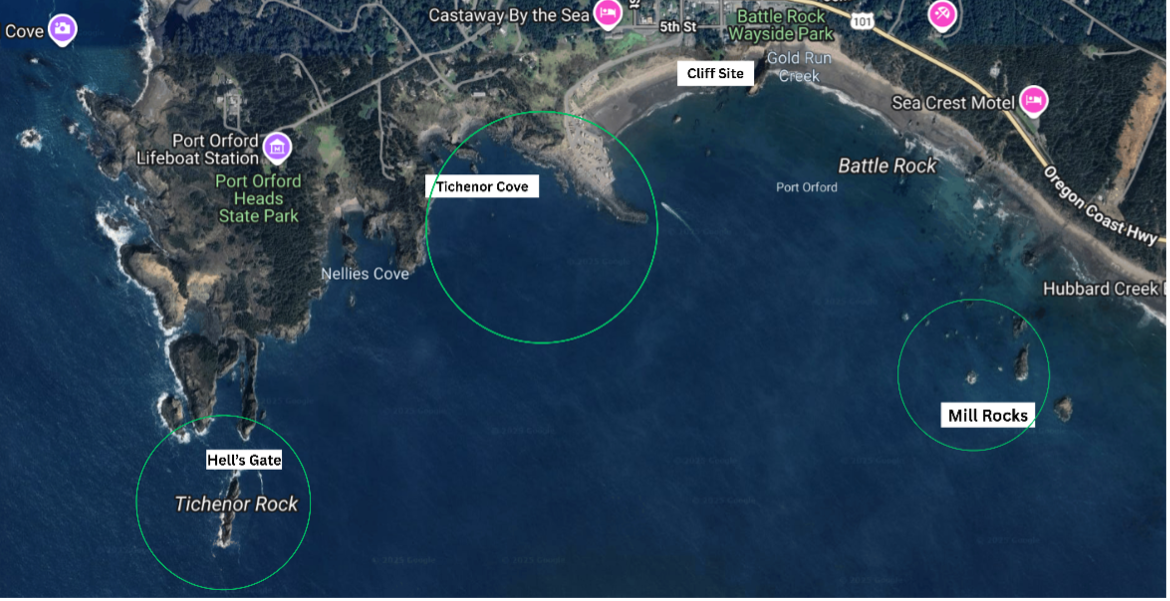
My day-to-day life as a TOPAZ/JASPER intern here at the Port Orford Field Station looks one of two ways: either on the kayak or the cliff site. When we are ocean kayaking, we go to our 12 sampling sites in the Mill Rocks and Tichenor Cove study areas (Fig. 1), where we collect zooplankton samples (Fig. 2) and oceanographic data with our RBR (an oceanographic instrument), as well as GoPro footage. When on the cliff site, we keep our eyes peeled for any whales to take pictures of them and mark their location in the water with a theodolite.
Fig. 1: Map of our study sites (Tichenor Cove and Mill Rocks) and where we have been seeing gray whales (Hell’s Gate) circled in green, and our Cliff Site.
Fig. 2: Miranda Fowles out on the kayak pointing at her zooplankton samples.
A theodolite is an instrument that is used for mapping and engineering; in our case it is used to track where a gray whale blows and surfaces (For more info, please see this blog by previous intern Jonah Lewis). Each time a whale surfaces, we use the theodolite to create a point in space that marks its location. Once we have multiple points, we can draw lines between each point to establish the track of the whale. These tracklines can then be used to make assumptions of the whales’ behavior. For example, if the trackline is straight, and the individual is moving at a consistent speed and direction, we can assume the whale is transiting. Whereas if the trackline is going back and forth in one small area, the whale is likely searching or foraging for food (Hildebrand et al., 2022).
In last week’s blog my peer Nautika Brown showed how photo ID is a critical part in our field methods. When theodolite tracking, we assign a number with each new individual whale observation. If the whale is close enough, we also capture photographs of the whale (Fig. 3) and match it up to its given number, allowing us to link the trackline to an individual whale so we can understand more about individual behavior. Documenting individual specific behavior is important because previous research has shown that age, size and the individual ID of a whale can all influence different foraging tactic use (Bird et al., 2024). Therefore, each season as we collect more and more data, we establish a repertoire of recurring or new behaviors to sieve for trends and patterns.
Fig. 3: Photo of a gray whale surfacing captured from our cliff site.
I find animal behavior to be an integral role in many ecological studies, and I am intrigued to explore this topic more. As marine mammals that spend most of their time underwater, cetaceans are quite an inconspicuous species to study (Bird et al., 2024), but by studying their ecology through photo ID and theodolite tracking we get insight into who they are, how they behave, and where they go.
Up until this point in the season, we have theodolite tracked gray whales for 12 hours and 3 minutes (woohoo). Interestingly, most of these tracks of whales have been near an area called “Hell’s Gate”, which is located around large rocks toward the far west of our study site (Figs. 2 and 4). We can assume, but cannot be sure, that the whales are feeding here because they spend so much time in the area, and return day after day. According to Dr. Torres, the consistent use of this area near Hell’s Gate by gray whales is unusual. In the prior 10 years of the TOPAZ project, few whales have been tracked foraging in this area near Hell’s Gate, but rather most whales have foraged in the Mill Rocks and Tichenor Cove areas. It is interesting to think about why the whales are behaving differently this year. Maybe this is due to variations in prey availability at these different sites. In recent years, Port Orford has been affected by a surge in purple sea urchin density, which have overgrazed the once prominent kelp forests here. A high urchin density decreases the kelp condition, which then leads to less habitat for zooplankton, creating a decline in prey availability for gray whales (Hildebrand et al., 2024). Upon reflection of my time on the kayak, I have noticed minimal kelp and low zooplankton abundance when conducting our zooplankton drops in our Mill Rocks and Tichenor Cove study sites. Additionally, I have also noticed many purple sea urchins in our GoPro videos. With the effects of this trophic cascade in mind, not observing any gray whales in our traditional study sites is understandable. With these gray whales more commonly seen near Hell’s Gate this year, I am curious to know what prey is attracting them there. Perhaps it is a different type of prey species or one that is high in caloric value than what is in the Mill Rocks and Tichenor Cove areas.
Fig. 4: Intern Nautika Brown looking at Hell’s Gate through the binoculars. Hell’s Gate is the passage between the two large boulders in the distance.
From actively observing whales and learning from my mentor, Celest, I have started to understand that behavior is a critical piece to any form of studying gray whales (and all species). By integrating photo-ID and theodolite tracking, we can learn so much about whale behavior, from where they eat, who is spending time where, and how they may adjust their behavior in response to a changing environment. The TOPAZ/JASPER internship has allowed me to truly comprehend what field research is like, how studying the behaviors of an individual is important, and how detail and patience are extremely necessary when collecting data. As this summer is continuing, I wonder if we will continue to see gray whales primarily feeding in the Hell’s Gate area, or if we will start to observe them more in the Mill Rocks and Tichenor Cove sites like previous years. The thrill of seeing gray whales is unlike any other, and I am so ready to see more whales this season!
References:
Bird, C. N., Pirotta, E., New, L., Bierlich, K. C., Donnelly, M., Hildebrand, L., Fernandez Ajó, A., & Torres, L. G. (2024). Growing into it: Evidence of an ontogenetic shift in grey whale use of foraging tactics. Animal Behaviour, 214, 121–135. https://doi.org/10.1016/j.anbehav.2024.06.004
Hildebrand, L., Derville, S., Hildebrand, I., & Torres, L. G. (2024). Exploring indirect effects of a classic trophic cascade between urchins and kelp on zooplankton and whales. Scientific Reports, 14(1), 9815. https://doi.org/10.1038/s41598-024-59964-x
Hildebrand, L, Sullivan, F. A., Orben, R. A., Derville. S., Torres L. G. (2022) Trade-offs in prey quantity and quality in gray whale foraging. Mar Ecol Prog Ser 695:189-201 https://doi-org.oregonstate.idm.oclc.org/10.3354/meps14115
Nautika Brown, GEMM Lab TOPAZ/JASPER Intern, recent Lake Roosevelt high school graduate
Hi everyone! I’m Nautika Brown, a recent graduate at Lake Roosevelt High School in a small town on the Colville Indian Reservation in Washington.
Growing up in beautiful Eastern Washington, I spent most all my days outside and, from the time I could swim, I was in the water. When I was little, I used to wish I was a fish so I could live underwater and swim every day of my life. And since then, I have always been fascinated by all animals that could live in and around water. This very fascination is what sparked the idea of becoming a marine biologist. Animals AND water, perfect!
(Left): Nautika holding a fish she caught back home in Buffalo Lake.
(Right) Nautika with a new type of catch (purple sea urchin) while conducting a zooplankton drop at station MR 18.
Although, as you might assume, living on a reservation surrounded by wheat fields and a few lakes, there weren’t a lot of opportunities to explore my passion. Hence, when I came across a flyer for the 2025 TOPAZ/JASPER internship just a few days before the deadline, I submitted my application as soon as I could. I was so thrilled, I couldn’t imagine getting the chance to kayak with whales on the ocean! It was all I could talk about for weeks on end.
Since starting my internship here in Port Orford, I have learned so many new things. During our first couple weeks at the field station, we went through a few different classes and trainings, one of them being a presentation on photo identification by GEMM Lab PhD candidate Lisa Hildebrand. Prior to this presentation, I had no idea photos were so important in marine mammal science. During this presentation, I learned about the many different identifiers of a whale and how you can apply them when looking at photos to identify a specific individual. For example, Lisa’s rule of three’s: to confidently ascertain an individual’s ID, at least 3 consistent characteristics between photos must be matched. At the end of this presentation, we even played a guessing game to test our new photo ID’ing skills. (I did pretty well – not to brag or anything.)
Now with my new photo ID skills, I was excited to capture a photo of a gray whale. On our second day of training, we did spot a whale—but thanks to my newly learned photo-ID skills, I quickly realized it wasn’t the gray whale I was expecting. When the whale first surfaced, I noticed the lack of dorsal knuckles and its distinctly darker body—clear signs it wasn’t a gray whale, but a humpback whale! While it is common to see gray whales from shore along the Oregon coast as they feed in the very nearshore habitat, humpback whales are typically found in much deeper waters, further from shore. Over the last week we have seen a humpback whale within our study site across several days—and we’re not the only ones! When chatting with the local fisherman pre and post kayak, a few have expressed their own excitement about seeing a humpback so close to shore as well. Throughout our conversations, the question of why a humpback would be so close to shore weighed on our minds, leading me to do my own online research.
To investigate whether these humpback sightings have been of the same individual or multiple different whales, I decided to review the photos we have captured to try and determine a match. Once I conducted a first pass of the photos, I downloaded 10 of the most clear and definite shots and compared the photos using Lisa’s rule of threes. After reviewing the photos, I noticed that the humpback whale’s dorsal hump resembled one from a previous sighting, but I couldn’t find any other distinguishing markings on its body. While I couldn’t confirm we have been observing the same humpback whale, I gained a deeper understanding of the importance of clear, high-quality photos in photo-ID work.
(Left) Nautika getting ready to take pictures of whales with camera on our cliff site.
(Right) Picture of humpback whale caught on camera on our 2nd day of training
After reading a few articles about humpback whale migration through Oregon, I found a few potential reasons behind this whale’s occurrence close to the shores of Port Orford. During the summer months, humpbacks travel to colder, more nutrient-dense places to feed, often near the shelf break (where the depth of the ocean suddenly gets deeper, around 200 m). Interestingly, the shelf break near Port Orford is not far from shore, and is a known hotspot for foraging humpback whales in the summer (Derville et al. 2022). Humpback whales filter-feed on krill and small fish, so perhaps enough prey has moved into the waters near Port Orford to attract a humpback so close to shore. Another reason for this humpback to be close to shore could be the effects of climate change. As the waters warm, food distribution changes, causing multiple species, including humpbacks, to change their feeding grounds and migration routes (read more here). Although the humpback sightings are outside the range of our kayak zooplankton sampling stations, it would be interesting to see what prey is in the water that is keeping them around.
So far, I have learned the importance of photo identification in marine mammal science and the many ways it can be used. I’m especially grateful for Lisa’s fun and insightful presentation at the start of the season and even more surprised by how quickly I was able to put those photo-ID skills into practice. With three weeks left in the field season, I’m excited to keep building on what I’ve learned and to keep growing my skills. And speaking of building, I’m also curious to see how my “kayak muscles” are shaping up by the end of this amazing TOPAZ/JASPER internship!
(Left) Nautika and Celest on kayak heading Mill Rocks stations.
(Right) Miranda and Nautika wrapping up kayak training with a celebratory team dab
Derville, S., D.R. Barlow, C. Hayslip, and L.G. Torres, Seasonal, Annual, and Decadal Distribution of Three Rorqual Whale Species Relative to Dynamic Ocean Conditions Off Oregon, USA. Frontiers in Marine Science, 2022. 9: p. 868566.
Celest Sorrentino, GEMM Lab Master’s student, OSU Department of Fisheries, Wildlife and Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
As a loyal and trusted GEMM Lab blog reader, I am sure you know just what time of year it is: the beginning of the 11th annual TOPAZ/JASPER field season where we study whales and their prey while also training the next generation of scientists. The start of the season has been kicked into high tail already and we have many updates to share. Fear not, dear reader, as I am here to release you from relentlessly refreshing your inbox for the long-awaited introduction of the TOPAZ/JASPER team that is taking the project into their second decade.
But first, to appreciate the present milestone, it’s worth revisiting the legacy of those who guided us to this moment. The TOPAZ/JASPER projects began in 2015, with PI. Dr. Leigh Torres and master’s student Florence Sullivan (2015-2018), and continued forward with Lisa Hildebrand (2018-2021), and Allison Dawn (2022-2024). Now, as a new droplet in this stream of brilliant leaders before me, I feel immense gratitude to be the master’s student leading the TOPAZ/JASPER team this summer. Having been trained by Allison Dawn with Team Protein in 2024, and full unwavering support from Leigh and each leader before me, I enter this new role with confidence and excitement for the next six gray-whale-and-zooplankton filled weeks of data collection. Now, let’s meet the young scientist interns for 2025!
(Left picture) Maddie (right) with Nautika (top) and Celest (left) during their kayak training.
(Right picture) Photo Maddie took of a humpback in the Port Orford Bay.
Madison (Maddie) Honomichl is a senior wrapping up her last semester of undergrad at CSU Monterrey Bay this fall to gain a degree in Marine Science. As the GEMM Lab’s REU intern this summer, Maddie began her internship in June by joining me in Newport to learn more about gray whale and pymgy blue whale mother-calf relationships. Without spoiling too much (you’ll hear more from her in her blog post in just a few weeks!) her project focuses on capturing mother-calf blow synchrony of gray and blue whales in drone footage. Now in Port Orford, her gifted talent for photography has been excellent in helping capture photos of traveling whales on the cliff.
(Left picture) Nautika finding a purple urchin after a successful zooplankton drop at our station MR 18.
(Right picture) Miranda(front) and Nautika(rear) after their first kayak training, where Nautika accidentally fell into the water but got back on the kayak in record breaking time, still in good spirits to dab!
Nautika Brown is one of our high school interns from Coulee Dam, Washington. Having just graduated, Nautika’s ambition and passion for studying wildlife lead her to apply to our TOPAZ/JASPER project and we are so happy she did. Accidentally hilarious, she has made everything from kayak training to zooplankton identification that much more enjoyable—reminding the team to have some fun while still getting the job done.
(Left picture) Dawson leading the team with the heavy theodolite stand up to the cliff.
(Right picture) The team hyper locked in on tracking a humpback whale in the bay, working together to describe the position of the whale for Dawson on the theodolite.
Dawson Mohney is our Port Orford local, having recently graduated from Pacific High School in May. Though he might not know the best spots around town, Dawson’s demeanor mirrors that of Port Orford itself: kind, welcoming, and always helpful. Always up for any task, he is the first to ask if anyone needs help with carrying equipment up to the cliff or cooking a ground beef refried beans mash for team dinner. Come fall Dawson is excited to start his first semester at Southwestern Oregon Community college.
(Left picture) Miranda enjoying an outdoor stroll of Port Orford beaches.
(Right picture) Miranda stoked on catching so many atylus tridens for her first kayak training day!
Miranda Fowles is a recent graduate at Oregon State University having completed her major in Fisheries, Wildlife, and Conservation Sciences with a minor in Spanish. Originally from Seattle, her childhood memories include kayaking with her family, so ocean kayaking has come naturally. Miranda’s genuine curiosity shines through in her eagerness to ask questions about whale life histories and their social dynamics. She’s expressed a clear passion for continuing her journey in marine science and academia.
We are now T-minus 2 days until the last of the team’s training period, and we couldn’t be more thrilled for the 4 more weeks to come. Through unexpected wildlife sightings and spontaneous team jokes, our team has only grown stronger and more connected. For all of the interns, this experience is not only their first experience with marine fieldwork, but also their longest. Training days have been both rewarding and physically strengthening; we’ve watched harbor seals lounging between Mill Rocks and tracked a particularly active humpback whale that keeps surfacing in the bay—all while developing what we now call our “ultimate kayak muscles.” By the time lunch rolls around, it feels like an ultimate power recharge, to continue forward with data processing. As any marine field scientist will tell you: there’s something deeply satisfying about coming back to shore and sinking your teeth into a handmade sandwich.
And speaking of our absolute craving for sandwiches, this team has unexpectedly brought back the 2010s dab—with such enthusiasm that it was only right to fuse “dab” with our love for chips-in-sandwiches. With this, I share with your our new, very official team name:
Team Dabwich.
With the right amount of salty, silly, and scienc-y, Team Dabwich is ready to crush the 11th TOPAZ/JASPER field season.
Team Dabwich dabbing right before a successful kayak training ヽ(⌐_⌐ゞ)!
By Rachel Kaplan, PhD candidate, Oregon State University College of Earth, Ocean, and Atmospheric Sciences and Department of Fisheries, Wildlife, and Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
At the beginning of a graduate program, it’s common for people to tell you how quickly the time will pass, but hard to imagine that will really be the case. Suddenly, I’ve been working on my PhD for almost five years, and I’ll defend in just over two weeks. As I look back, I am amazed by how much I have learned and grown during this time, and how all the different parts of my graduate school experience have woven together. I began my program in 2020 with an intense “bootcamp” of oceanographic coursework, and am ending in 2025 with new analytical skills, a few publications, and a ton of new thoughts about whales and the zooplankton krill, the subjects of my research. My PhD work encapsulates all those different elements in an exploration of ecological relationships between baleen whale predators and their krill prey – which I now see as an expression of oceanographic and atmospheric processes.
Figure 1. One of my favorite sightings during my PhD fieldwork was a group of seven fin whales in Antarctica, on Christmas 2024. Photo: Rachel Kaplan
Oceanographic processes drive prey quantity and quality across time and space, shaping the preyscape encountered by predators on their foraging grounds and driving habitat use (Fleming et al., 2016; Ryan et al., 2022). Aspects of prey including distribution, energy density, and biomass therefore represent mechanistic links between ocean and atmospheric conditions (e.g., El Niño Southern Oscillation cycles, circulation patterns, and upwelling processes) and diverse aspects of marine predator ecology, including spatiotemporal distributions, foraging behaviors, reproductive success, population size, and health. Both predator and prey species are impacted by environmental variability and climate change (e.g., Hauser et al., 2017; Atkinson et al., 2019; Perryman et al., 2021), and events like marine heatwaves and harmful algal blooms can force ecosystem changes on short, seasonal time scales (e.g. McCabe et al., 2016; Fisher et al., 2020). However, many marine species have some degree of plasticity that allows them to still accomplish life history events in the face of ecosystem variability (e.g., Lawrence, 1976; Oestreich, 2022), which may provide the capacity to adapt to climate change processes.
Observing and describing predator-prey relationships is complex due to the scale-dependent nature of these relationships (Levin, 1992). Each chapter of my dissertation considered krill, a globally-important prey type, from the perspective of baleen whales, which are krill predators. Chapter 2 used a comparative analysis to identify the optimal spatial scale at which to observe baleen whale-krill relationships on the Northern California Current (NCC) foraging grounds. We found correlations at a 5 km scale to be strongest, which can provide a useful starting point for further studies in the NCC and other systems. Chapter 3 used this spatial scale to compare several aspects of krill prey quality and quantity as predictors of humpback whale (Megaptera novaeangliae) distributions in the NCC. The best performing metric was a species, season, and spatially informed krill swarm biomass variable – yet the comparable performance of a simple acoustic abundance metric indicated that it can act as a reliable proxy for biomass. This finding may be advantageous for future research, as measuring the acoustic proxy is less computationally intensive and relies on fewer datastreams. Interestingly, one of this study’s best-performing models was based on only the proportion of Thysanoessa spinifera in krill swarms, which is also a highly accessible variable due to effective krill species distribution modeling in the NCC (Derville et al., 2024). Integrating the acoustic abundance proxy and krill species distribution predictions, two relatively simple metrics, could support predictions of humpback whale distributions in the NCC and inform whale-prey research in other ecosystems.
Figure 2. Collecting samples of individual krill gave us the opportunity to learn about their quality as prey for whales in the Northern California Current. Photo: Courtney Flatt
Studies relating predator foraging to prey characteristics often rely on metrics such as prey biomass or energy density (Schrimpf et al., 2012; Savoca et al., 2021; Cade et al., 2022), but the tendency of krill to form aggregations introduces dimensionality to krill prey quality. Chapter 4 showed that elements of krill swarm structure (particularly depth, proportion of T. spinifera, and metrics describing how krill occupy space within swarms) may be mechanistic drivers of variable blue, fin and humpback whale distribution patterns on the NCC foraging grounds. These findings suggest that krill swarm characteristics may be important links between baleen whales and the foraging environment. Swarm characteristics may be considered a component of krill prey quality for baleen whales, and future research could illuminate direct causal relationships between oceanographic conditions, krill swarming responses, and niche expression in baleen whale predators.
The relationships between baleen whale distributions and krill quantity and quality explored in the first chapters of my dissertation may also shed light on other aspects of baleen whale ecology. The final chapter considers overwintering trends in global baleen whale populations, and examines the wintertime Western Antarctic Peninsula (WAP) as a case study. Extended humpback whale presence on the WAP feeding grounds may be driven by the profitable feeding areas and elevated energy content of krill during the winter months, and may reflect the high energetic needs of certain demographic subgroups (e.g. lactating females, juveniles). Wintertime humpback whale presence may also reflect adaptation to multifaceted competitive pressure on krill resources that are declining due to climate change (Atkinson et al., 2019), including consumption by growing baleen whale populations (Johnston et al., 2011) and a fishery whose catch limits may be impacting krill predators (Watters et al., 2020; Savoca et al., 2024). This work demonstrates how investigating prey quality during the winter months can contextualize baleen whale overwintering on the foraging grounds. It also provides a meaningful violation of the canonical baleen whale migration paradigm central to marine mammal science, which may lessen the efficacy of whale monitoring programs and management policies.
Figure 3. We were surprised to see humpback whales like this one in Antarctica during the winter months — which raised a number of questions about overwintering of baleen whales on foraging grounds around the world. Photo: Giulia Wood
Management efforts that aim to mitigate risk to whales often hinge on predictive modeling of whale distributions. Species distribution models (SDMs) can provide managers with spatially and temporally explicit predictions of protected species occurrences (Wikgren et al., 2014; Santora et al., 2020), but species distributions in rapidly changing ecosystems are difficult to predict (Muhling et al., 2020). Findings from this dissertation may inform modeling efforts by suggesting meaningful predictor variables for SDMs, such as krill species on the NCC foraging grounds and swarm energy density at the WAP. This work also speaks to meaningful spatial scales for analyzing predator-prey relationships (i.e., 5 km), and relevant elements of temporal variability (e.g., seasonal cycles of krill energy density).
Just as marine predator-prey relationships are shaped by ocean processes, they likewise have consequences for those processes. For example, krill and other zooplankton are capable of generating large-scale mixing that can overcome stratification of water masses and alter water column structure (Noss and Lorke, 2014). Baleen whales influence global carbon cycles due to the huge amount of prey they consume (Savoca et al., 2021; Pearson et al., 2023) and transport important nutrients along the “great whale conveyer belt” during their vast migrations (Roman et al., 2025). Baleen whales seek krill as an essential prey resource on foraging grounds around the globe, and the impact of this trophic interaction scales up, with implications for ecosystem functioning and management. Continued research into the spatiotemporally dynamic relationships between krill and baleen whales improves our understanding of ocean functioning, and can improve our capacity to live as part of this system.
References
Atkinson, A., Hill, S. L., Pakhomov, E. A., Siegel, V., Reiss, C. S., Loeb, V. J., Steinberg, D. K., et al. 2019. Krill (Euphausia superba) distribution contracts southward during rapid regional warming. Nature Climate Change, 9: 142–147.
Cade, D. E., Kahane-Rapport, S. R., Wallis, B., Goldbogen, J. A., and Friedlaender, A. S. 2022. Evidence for Size-Selective Predation by Antarctic Humpback Whales. Frontiers in Marine Science, 9: 747788.
Derville, S., Fisher, J. L., Kaplan, R. L., Bernard, K. S., Phillips, E. M., and Torres, L. G. 2024. A predictive krill distribution model for Euphausia pacifica and Thysanoessa spinifera using scaled acoustic backscatter in the Northern California Current. Progress in Oceanography: 103388.
Fisher, J. L., Menkel, J., Copeman, L., Shaw, C. T., Feinberg, L. R., and Peterson, W. T. 2020. Comparison of condition metrics and lipid content between Euphausia pacifica and Thysanoessa spinifera in the northern California Current, USA. Progress in Oceanography, 188.
Fleming, A. H., Clark, C. T., Calambokidis, J., and Barlow, J. 2016. Humpback whale diets respond to variance in ocean climate and ecosystem conditions in the California Current. Glob Chang Biol, 22: 1214–24.
Hauser, D. D. W., Laidre, K. L., Stafford, K. M., Stern, H. L., Suydam, R. S., and Richard, P. R. 2017. Decadal shifts in autumn migration timing by Pacific Arctic beluga whales are related to delayed annual sea ice formation. Global Change Biology, 23: 2206–2217.
Johnston, S. J., Zerbini, A. N., and Butterworth, D. S. 2011. A Bayesian approach to assess the status of Southern Hemipshere humpback whales (Megaptera novaeangliae) with an application to Breeding Stock G. J. Cetacean Res. Manage.: 309–317. International Whaling Commission.
Lawrence, J. M. 1976. Patterns of Lipid Storage in Post-Metamorphic Marine Invertebrates. American Zoologist, 16: 747–762. Oxford University Press (OUP).
Levin, S. A. 1992. The Problem of Pattern and Scale in Ecology: The Robert H. MacArthur Award Lecture. Ecology, 73: 1943–1967.
McCabe, R. M., Hickey, B. M., Kudela, R. M., Lefebvre, K. A., Adams, N. G., Bill, B. D., Gulland, F. M., et al. 2016. An unprecedented coastwide toxic algal bloom linked to anomalous ocean conditions. Geophys Res Lett, 43: 10366–10376.
Muhling, B. A., Brodie, S., Smith, J. A., Tommasi, D., Gaitan, C. F., Hazen, E. L., Jacox, M. G., et al. 2020. Predictability of Species Distributions Deteriorates Under Novel Environmental Conditions in the California Current System. Frontiers in Marine Science, 7.
Noss, C., and Lorke, A. 2014. Direct observation of biomixing by vertically migrating zooplankton. Limnology and Oceanography, 59: 724–732. Wiley.
Oestreich, W. 2022. Acoustic signature reveals blue whales tune life‐history transitions to oceanographic conditions. Functional Ecology. https://besjournals.onlinelibrary.wiley.com/doi/10.1111/1365-2435.14013 (Accessed 20 September 2024).
Pearson, H. C., Savoca, M. S., Costa, D. P., Lomas, M. W., Molina, R., Pershing, A. J., Smith, C. R., et al. 2023. Whales in the carbon cycle: can recovery remove carbon dioxide? Trends in Ecology & Evolution, 38: 238–249.
Perryman, W. L., Joyce, T., Weller, D. W., and Durban, J. W. 2021. Environmental factors influencing eastern North Pacific gray whale calf production 1994–2016. Marine Mammal Science, 37: 448–462. Wiley.
Roman, J., Abraham, A. J., Kiszka, J. J., Costa, D. P., Doughty, C. E., Friedlaender, A., Hückstädt, L. A., et al. 2025. Migrating baleen whales transport high-latitude nutrients to tropical and subtropical ecosystems. Nature Communications, 16: 2125. Nature Publishing Group.
Ryan, J. P., Benoit-Bird, K. J., Oestreich, W. K., Leary, P., Smith, K. B., Waluk, C. M., Cade, D. E., et al. 2022. Oceanic giants dance to atmospheric rhythms: Ephemeral wind-driven resource tracking by blue whales. Ecology Letters, 25: 2435–2447.
Santora, J. A., Mantua, N. J., Schroeder, I. D., Field, J. C., Hazen, E. L., Bograd, S. J., Sydeman, W. J., et al. 2020. Habitat compression and ecosystem shifts as potential links between marine heatwave and record whale entanglements. Nat Commun, 11: 536.
Savoca, M. S., Czapanskiy, M. F., Kahane-Rapport, S. R., Gough, W. T., Fahlbusch, J. A., Bierlich, K. C., Segre, P. S., et al. 2021. Baleen whale prey consumption based on high-resolution foraging measurements. Nature, 599: 85–90.
Savoca, M. S., Kumar, M., Sylvester, Z., Czapanskiy, M. F., Meyer, B., Goldbogen, J. A., and Brooks, C. M. 2024. Whale recovery and the emerging human-wildlife conflict over Antarctic krill. Nature Communications, 15: 7708. Nature Publishing Group.
Schrimpf, M., Parrish, J., and Pearson, S. 2012. Trade-offs in prey quality and quantity revealed through the behavioral compensation of breeding seabirds. Marine Ecology Progress Series, 460: 247–259.
Watters, G. M., Hinke, J. T., and Reiss, C. S. 2020. Long-term observations from Antarctica demonstrate that mismatched scales of fisheries management and predator-prey interaction lead to erroneous conclusions about precaution. Scientific Reports, 10: 2314.
Wikgren, B., Kite-Powell, H., and Kraus, S. 2014. Modeling the distribution of the North Atlantic right whale Eubalaena glacialis off coastal Maine by areal co-kriging. Endangered Species Research, 24: 21–31.
Dr. Enrico Pirotta (CREEM, University of St Andrews) and Dr. Leigh Torres (GEMM Lab, MMI, OSU)
The health of animals affects their ability to survive and reproduce, which, in turn, drives the dynamics of populations, including whether their abundance trends up or down. Thus, understanding the links between health and reproduction can help us evaluate the impact of human activities and climate change on wildlife, and effectively guide our management and conservation efforts. In long-lived species, such as whales, once a decline in population abundance is detected, it can be too late to reverse the trend, so early warning signals are needed to indicate how these populations are faring.
We worked on this complex issue in a study that was recently published in the Journal of Animal Ecology. In this paper, we developed a new statistical approach to link three key components of the health of a Pacific Coast Feeding Group (PCFG) gray whale (namely, its body size, body condition, and stress levels) to a female’s ability to give birth to a calf. We were able to inform these metrics of whale health using an eight-year dataset derived from the GRANITE project of aerial images from drones for measurements of body size and condition, and fecal samples for glucocorticoid hormone analysis as an indicator of stress. We combined these data with observations of females with or without calves throughout the PCFG range over our study period.
We found that for a female to successfully have a calf, she needs to be both large and fat, as these factors indicate if the female has enough energy stored to support reproduction that year (Fig. 1). Remarkably, we also found indication that females with particularly high stress hormone levels may not get pregnant in the first place, which is the first demonstration of a link between stress physiology and vital rates in a baleen whale, to our knowledge.
Figure 1. Taken from Pirotta et al. (2025), Fig. 5. Combined relationship of PCFG gray whale length and nutritional state (combination of body size and condition) in the previous year with calving probability, colored by whether the model estimated an individual to have calved or not at a given reproductive opportunity.
Our study’s findings are concerning given our previous research indicating that gray whales in this PCFG sub-group have been growing to shorter lengths over the last couple of decades (Pirotta et al. 2023), are thinner than animals in the broader Eastern North Pacific gray whale population (Torres et al, 2022), and show an increase in stress-related hormones when exposed to human activities (Lemos et al, 2022; Pirotta et al. 2023). Furthermore, in our recent study we also documented that there are fewer young individuals than expected for a growing or stable population (Fig. 2), which can be an indicator of a population in decline since there may not be many individuals entering the reproductive adult age groups. Altogether, our results act as early warning signals that the PCFG may be facing a possible population decline currently or in the near future.
Figure 2. Taken from Pirotta et al. (2025), Fig. 1. Age structure diagram for 139 PCFG gray whales in our dataset. Each bar represents the number of individuals of a given age in 2023, with the color indicating the proportion of individuals of that age for which age is known (vs. estimated from a minimum estimate following Pirotta, Bierlich, et al., 2024). The red line reports a smooth kernel density estimate of the distribution.
These findings are sobering news for Oregon residents and tourists who enjoy watching these whales along our coast every summer and fall. We have gotten to know many of these whales so well – like Scarlett, Equal, Clouds, Lunita, and Pacman, who you can meet on our IndividuWhale website – that we wonder how they will adapt and survive as their once reliable habitat and prey-base changes. We hope our work sparks collective and multifaceted efforts to reduce impacts on these unique PCFG whales, and that we can continue the GRANITE project for many more years to come to monitor these whales and learn from their response to change.
This work exemplifies the incredible value of long-term studies, interdisciplinary methods, and effective collaboration. Through many years of research on this gray whale group, we have collected detailed data on diverse aspects of their behavior, ecology and life history that are critical to understanding their response to disturbance and environmental change, which are both escalating in the study region. We are incredibly grateful to the following members of the PCFG Consortium for contributing sightings and calf observation data that supported this study: Jeff Jacobsen, Carrie Newell, NOAA Fisheries (Peter Mahoney and Jeff Harris), Cascadia Research Collective (Alie Perez), Department of Fisheries and Oceans, Canada (Thomas Doniol-Valcroze and Erin Foster), Mark Sawyer and Ashley Hoyland, Wendy Szaniszlo, Brian Gisborne, Era Horton.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly alert when we make a new post! Just add your name into the subscribe box below!
References:
Lemos, Leila S., Joseph H. Haxel, Amy Olsen, Jonathan D. Burnett, Angela Smith, Todd E. Chandler, Sharon L. Nieukirk, Shawn E. Larson, Kathleen E. Hunt, and Leigh G. Torres. “Effects of Vessel Traffic and Ocean Noise on Gray Whale Stress Hormones.” Scientific Reports 12, no. 1 (2022): 18580. https://dx.doi.org/10.1038/s41598-022-14510-5.
Pirotta, Enrico, Alejandro Fernandez Ajó, K. C. Bierlich, Clara N Bird, C Loren Buck, Samara M Haver, Joseph H Haxel, Lisa Hildebrand, Kathleen E Hunt, Leila S Lemos, Leslie New, and Leigh G Torres. “Assessing Variation in Faecal Glucocorticoid Concentrations in Gray Whales Exposed to Anthropogenic Stressors.” Conservation Physiology 11, no. 1 (2023). https://dx.doi.org/10.1093/conphys/coad082.
Torres, Leigh G., Clara N. Bird, Fabian Rodríguez-González, Fredrik Christiansen, Lars Bejder, Leila Lemos, Jorge Urban R, et al. “Range-Wide Comparison of Gray Whale Body Condition Reveals Contrasting Sub-Population Health Characteristics and Vulnerability to Environmental Change.” Frontiers in Marine Science 9 (2022). https://doi.org/10.3389/fmars.2022.867258. https://www.frontiersin.org/article/10.3389/fmars.2022.867258
Dr. Clara Bird, Postdoctoral Scholar, OSU Department of Fisheries, Wildlife, and Conservation Sciences, GEMM Lab & LABIRINTO
Cycles can be found everywhere in nature and our lives. From tides and seasons to school years and art projects, we’re constantly experiencing cycles of varying scales. Spring on the Oregon coast brings several important cyclical events: more daylight, the oceanographic spring transition, and the return of our beloved gray whales – just to name a few. On my own personal scale, I’ve been thinking about the cycles we experience as scientists a lot lately, since I’ve recently transitioned out of graduate school and into my current position as a postdoctoral scholar.
Starting this new postdoc has been a bit jarring, as it’s felt like starting over. Even though I’m still working at the Marine Mammal Institute and still studying gray whales, I’ve been learning new skills, knowledge and theory, which pushes me to re-start the cycle of the scientific method, the process we follow in research (Figure 1). Broadly, we start by observing a system and asking a question about a potential pattern or event we see. We then come up with a hypothesis (or two or ten) to address our question(s). The next steps are to collect the data we need to answer our question(s) and test our hypotheses, analyze that data (i.e. run some statistical models), and draw some conclusions from the analysis results. While it seems quite linear, the process of data collection and analysis always leads to more questions than answers, and we inevitably start the cycle all over again.
Figure 1. Schematic depicting the scientific method
Throughout my scientific training I’ve gained experience in all these phases, but I’ve also learned just how many add-ons and do-overs there are in this process (Figure 2). Developing questions and hypotheses often requires a long and winding path through the literature, depending on how much you already know. These steps are often some of the first and biggest steps in graduate school. You need to learn as much as you can about the field and questions you are interested in, as this will inform what has already been done, where the knowledge gaps are, and the hypotheses you’re developing. For example, we often back up a hypothesis with references to studies that have answered our question in different systems. The learning curve is steep, and it’s important to not understate the work that goes into this phase. Early in my career, I remember hearing that “asking the good questions” is a critical skill for research. At the time that sounded like some vague, innate characteristic, and working to gain this ability felt ambiguous and overwhelming. I was absolutely wrong. Like most skills, knowing how to ask good questions is more about experience than intelligence. Here, experience is a combination of reading the literature and practice formulating questions based on the literature.
Figure 2. A more realistic version of the scientific method
Beyond this lesson, I also had to learn that what qualifies a question as “good” also depends on the funding source. In many research institutions, including those in the U.S., scientists are responsible for finding the funding to run their research projects. Funding a project includes salary for the scientists (e.g., professors, grad students, post docs), the cost of collecting and analyzing the data (e.g., travel, equipment, boat time), and the cost of publishing and sharing our findings (e.g., publication costs). The programs we solicit funding from often have their own priorities, so a big part of the research cycle is finding a funding source that is interested in the kinds of questions you want to ask and then adjusting your own questions and hypotheses to align with the funding source’s priorities and budget. The actual application includes writing a proposal where we (1) summarize all the background research justifying the novelty and value of the questions we want to ask and backing up our hypotheses and (2) describe how we plan on answering those questions. Funding is competitive and we typically apply multiple times before being successful. Furthermore, we often apply to multiple funding sources to support the same project. Since each source has its own focus, this ends up being an exercise in coming up with multiple ways to frame and justify a project.
Once we have funding (which can be years after the start of the cycle), we can finally start collecting, analyzing, and interpreting the data. But each of these steps has its own sub-cycles and complexities. Data collection can take years and involve all kinds of troubleshooting equipment issues, logistics, and methods. Depending on your question, data processing and analysis may involve developing your own method. For example, our lab asks a lot of questions about the morphology and body condition of whales. But before we could answer those questions, we first had to work out the best way to accurately measure whales from drone imagery while accounting for measurement uncertainty (read more here). This separate cycle of method development involved so many sub-projects and new software tools that Dr. KC Bierlich now leads the Marine Mammal Institute’s Center of Drone Excellence (CODEX).
Data analysis and interpretation brings us back to the literature review part of the cycle. But now we are looking for examples of how similar data have been analyzed previously and for studies to which we can compare our results. Then, after testing out different models and triple checking our analysis, we’re finally ready to share our findings. We share our results through conference presentations, publications (after the peer review cycle), outreach talks, and press releases that lead to media pieces and interviews.
In addition to the excitement of sharing our findings with the world, we’re simultaneously hyper-aware of all the caveats and limitations of our work. We’re always left with a long list of follow-up questions, thus starting the cycle again. From a zoomed-out perspective these results can form a clean, linear story. But zooming in reveals the reality of years and years of multiple overlapping cycles that have had to pass roadblocks and restart countless times. For example, after nine years for research, the GRANITE project has produced an impressive suite of results addressing questions related to Pacific Coast Feeding Group gray whale morphology, health, hormones, space use, and behavior. It took years of data collection, proposal writing, training, and multiple researchers working through their own project cycles to get here (and we’re not done).
Transitioning out of graduate school has meant expanding my scope of attention to multiple cycles running in parallel, re-starting the literature review process for new projects, and spending a lot more time in the proposal writing sub-cycle. While it’s felt overwhelming at times, I’ve also enjoyed digging into new topics and skills. It’s an interesting balance of experiencing the discomfort that comes with being a beginner while simultaneously drawing comfort from the knowledge that I’ve experienced this cycle before and know how to learn something new.
A consequence of learning the scientific process is growing accustomed to this cyclical nature. As scientists we know that it’s a slow process, that every result is just the start of a new cycle, and that future work building on a result may agree or disagree with the previous finding. But the way scientific findings are shared with the public doesn’t necessarily reflect the process. Catchy headlines and brief summaries often present findings as definitive and satisfying conclusions to a story. Behind those headlines are years of set up, data collection, analysis, and a suite of caveats that we want to dig into in the future. The results of any given study reflect our best current knowledge at that point in the cycle. By design, that knowledge will grow and change as we move forward.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly alert when we make a new post! Just add your name into the subscribe box below!
When most people think about monitoring the health of a 40-ton gray whale, they picture blubber thickness, dive patterns, or perhaps growth rates. But what if some of the most telling signs are found not in the whale’s bulk, but right on the surface–embedded in its skin, and even crawling across it?
As part of the GRANITEproject, my research focuses on using a long-term photographic dataset (>347,000 photos from 10 years!) to evaluate epidermal indicators of stress and health in Pacific Coast Feeding Group (PCFG) gray whales (Eschrichtius robustus) foraging off the Oregon coast. My central questions ask:
Can we use features visible on the skin like epidermal diseases, lesion severity, scarring from orcas, boats, and fishing gear, and potentially cyamid loads as biomarkers of physiological stress or nutritional status?
How do these skin-based indicators correlate with environmental variables, prey availability, fecal hormones, and overall body condition?
By tracking these patterns across individuals and years, my goal is to understand how gray whales are responding to a changing ocean and whether their skin can tell us more about what’s under the surface.
What are cyamids?
Cyamids, more commonly known as “whale lice”, are small crustaceans that live exclusively on marine mammals. Despite their nickname, cyamids are not true lice—they’re actually amphipod ectoparasites, more closely related to beach hoppers than anything you’d find in your hair. For gray whales (Eschrichtius robustus), these tiny passengers are a constant presence throughout their lives.
Figure 1. Gray whale blow hole area covered in barnacles and cyamids. The circle inset shows a zoomed in area where you can see the orange cyamids aggregating near the more yellow barnacles.
Each whale can host thousands of cyamids at a time, with individuals often clustering in specific areas of the body that provide physical refuge from the currents: around the blowhole, in the crevices of flukes, along the rostrum, genital slits, and especially around wounds or skin irregularities (Figure 1). Unlike barnacles, which attach directly to the skin and remain stationary while they feed on nutrients in the passing water, cyamids grasp onto the whale’s body using claw-like appendages, feeding on sloughed skin and bodily fluids. This relationship is generally not thought to be harmful to the whale, but high cyamid loads can be indicative of poor health, injury, or compromised immune function.
There are several species of cyamids, and many are host-specific—meaning they’ve evolved alongside particular whale species. In gray whales, the most common is Cyamus scammoni, which specializes on gray whales and is rarely found elsewhere. Other species found on gray whales include Cyamus kessleri, and the rarer Cyamus ceti. Cyamids are transmitted primarily from mother to calf, which helps explain their host fidelity, but horizontal transmission (between unrelated individuals) may also occur during close contact which can explain some rare occurrences of cyamids that are found outside of their general host species. In fact, Cyamus ceti was only found once on gray whales in 1861 but is generally thought to be specific to bowhead whales, giving us potential insight into interspecies interactions (bowhead and gray whales can spatially overlap on Arctic foraging grounds).
Figure 2. Cyamus scammoni close up photographs of (A) aggregation, (B) juvenile stage, (C) dorsal side of an adult, and (D) ventral side of an adult showcasing the cyamids corkscrew shaped gills (Takeda et al. 2005)
Because cyamids are permanent residents of the whale’s skin, they offer a unique window into both individual whale life histories and broader ecological trends. Their location, abundance, and distribution can potentially inform us about wound healing, residency duration in foraging areas, and even stress or health status—which makes them an unexpectedly valuable focal point in drone and photograph-based monitoring efforts like in the GRANITE project.
Cyamid Life History
Cyamids are obligate ectoparasites, meaning they spend their entire life on a whale and cannot survive independently in the open ocean. Unlike free-swimming crustaceans, cyamids are permanently attached to the skin or embedded within crevices of the whale’s body, often clinging to roughened areas, scars, embedded barnacles, or calloused skin where they can anchor themselves more securely.
They begin life as tiny juveniles, hatching from eggs carried in the brood pouch of a female cyamid. Rather than undergoing a larval phase in the water column like many marine invertebrates, cyamids develop directly into miniature versions of adults and remain on the whale from birth. This direct development is essential because there’s no safe habitat for a larval cyamid in the open ocean: the host whale is both nursery and home.
Most transmission occurs from mother to calf during the close physical contact of early life. Calves born in the warm lagoons of Baja California, Mexico where gray whales calve and nurse during the winter inherit their cyamid colonies during nursing, rubbing, and swimming alongside their mothers. These early colonizers will multiply as the calf grows and can remain with the whale for years, forming the basis of a persistent, host-specific population.
For Cyamus scammoni specifically (our gray whale specific cyamid), adults will breed in the summer just before the southbound migration. Females will have around 1,000 eggs in their brood pouch, although only about a 60% are fertilized (Leung, 1976). These eggs will hatch in the fall while the gray whales take on their southbound migration but they will stay in the safety of the brood pouch for around 2 to 3 months. The juveniles will be released in the winter, when gray whales arrive in the Baja lagoons where they will then find shelter within the crevices of their host gray whale. Juveniles reach maturity during the northbound migration and will be a full-grown brood upon arrival to summer grounds. While the cycle takes about 8 months to complete, there are juveniles found along the gray whales year-round, leading us to believe that there is likely overlap between broods. For our less abundant Cyamus kessleri, the life cycle is very similar, but the juveniles reach maturity before the gray whales northbound migration to summer feeding grounds. Also, there are around 300 eggs in the Cyamus kessleri brood pouches that have a higher rate of fertilization (75-80%) than Cyamus scammoni (60%) (Leung, 1976)
In short, the life of a cyamid is fully bound to the life of a whale. Every migration, dive, foraging event, and scar the whale experiences becomes part of the cyamid’s environment. By studying them, we gain another lens through which to interpret the health, behavior, and ecology of gray whales on the Oregon Coast.
Uses in Cetacean Health Assessments
As we’ve established, cyamids have unique life histories as ectoparasites and may be valuable indicators in cetacean health assessments across multiple whale species. Because they often congregate around wounds, lesions, and areas of poor skin integrity, their presence and distribution can reveal important clues about a whale’s physical condition, injury history, and immune response. However, studies that have made these connections have variable results.
In species like North Atlantic right whales (Pettis et al. 2004, Pirotta et al. 2023), harbor porpoises (Lehnert et al. 2021), and gray whales (Raverty et al. 2024), researchers have used visual surveys and photographic analysis to quantify cyamid loads in living, stranded, and hunted whales. Researchers can score cyamid presence by identifying attachment sites (e.g. blowhole, scar, dorsal ridge) and estimating the relative coverage by using standardized reference images to maintain consistency. In these studies, whales with heavy cyamid coverage, especially in sensitive regions like the blowhole, mouthline, and genital area, often show signs of poor health or stress, such as emaciation, scarring from entanglement, or chronic skin conditions. Cyamid coverage is sometimes used alongside body condition indices and lesion scoring to build a more complete health profile (Pirotta et al. 2023). There are also studies that show no connections, or even positive connections between body condition and cyamid coverage (Von Duyke et al. 2016).
While cyamids are often associated with injured, inflamed, or otherwise damaged skin, there is no evidence that points towards cyamids directly damaging the skin themselves. However, more work needs to be done to assess their role in the healing processes. Additionally, it’s been noted that more work is needed on the role of cyamids and disease spread (Overstreet et al. 2009). For the PCFG, there is an iconic whale we call “Scarlett” (also known as “Scarback”) who has a large scar on the right side of her back that is highly identifiable due to the orange swarm of cyamids that are constantly surrounding the edges of the wound. She has managed to survive and thrive, producing many calves over the years, but questions remain: How are the cyamids affecting the healing process? Are they increasing or decreasing the risk of infection? How does the frequency of large injuries like this on whales contribute to the cyamid population over evolutionary time?
Figure 3. Right side of PCFG icon, “Scarlett” showing her massive scar covered with orange aggregations of cyamids.
Because whales are complex, highly mobile, long-lived creatures with a constant population of cyamid hitchhikers their skin condition is likely representative of specific to life history, phylogeography, and demographic traits of individuals. While we know that cyamids generally eat sloughed or damaged skin on the whale, what this behavior and symbiosis means for each whale’s individual physiology can be highly complex. Through our high-resolution drone and lateral imagery of the same individuals over time paired with other data sources, such as body condition and prey availability, cyamid scores can offer key insights into how environmental stressors and foraging success affect individual and population-level whale health.
These tiny crustaceans, clinging to the folds and scars of their hosts, might seem like background noise in a study focused on body condition or foraging ecology—but they’re far from incidental. In my research, I’ve come to see cyamids as part of the bigger story:silent indicators of stress, recovery, movement, and resilience. By pairing imagery of PCFG gray whale skin with data on prey availability and environmental conditions, I’m working to understand how foraging success and anthropogenic stressors (such as vessel traffic and entanglements) manifest not just in a whale’s body condition, but in the skin itself. The presence, distribution, and density of cyamids may offer yet another layer of insight into how gray whales are coping with changing ocean conditions. It’s a reminder that even the smallest details, like a patch of whale lice, can help us ask bigger questions about the health, resilience, and future of these cetaceans.
References
Callahan, C.M., n.d. MOLECULAR SYSTEMATICS AND POPULATION GENETICS OF WHALE LICE (AMPHIPODA: CYAMIDAE) LIVING ON GRAY WHALE ISLANDS.
Lehnert, K., IJsseldijk, L.L., Uy, M.L., Boyi, J.O., van Schalkwijk, L., Tollenaar, E.A.P., Gröne, A., Wohlsein, P., Siebert, U., 2021. Whale lice (Isocyamus deltobranchium & Isocyamus delphinii; Cyamidae) prevalence in odontocetes off the German and Dutch coasts – morphological and molecular characterization and health implications. International Journal for Parasitology: Parasites and Wildlife 15, 22–30. https://doi.org/10.1016/j.ijppaw.2021.02.015
Leung, Y., 1976. Life cycle of cyamus scammoni (amphipoda: cyamidae), ectoparasite of gray whale, with a remark on the associated species. Scientific Reports of the Whales Research Institute 28, 153–160.
Overstreet, R.M., Jovonovich, J., Ma, H., 2009. Parasitic crustaceans as vectors of viruses, with an emphasis on three penaeid viruses. Integrative and Comparative Biology 49, 127–141. https://doi.org/10.1093/icb/icp033
Pettis, H.M., Rolland, R.M., Hamilton, P.K., Brault, S., Knowlton, A.R., Kraus, S.D., 2004. Visual health assessment of North Atlantic right whales (Eubalaena glacialis) using photographs. Can. J. Zool. 82, 8–19. https://doi.org/10.1139/z03-207
Pirotta, E., Schick, R.S., Hamilton, P.K., Harris, C.M., Hewitt, J., Knowlton, A.R., Kraus, S.D., Meyer-Gutbrod, E., Moore, M.J., Pettis, H.M., Photopoulou, T., Rolland, R.M., Tyack, P.L., Thomas, L., 2023. Estimating the effects of stressors on the health, survival and reproduction of a critically endangered, long-lived species. Oikos 2023, e09801. https://doi.org/10.1111/oik.09801
Raverty, S., Duignan, P., Greig, D., Huggins, J.L., Huntington, K.B., Garner, M., Calambokidis, J., Cottrell, P., Danil, K., D’Alessandro, D., Duffield, D., Flannery, M., Gulland, F.M., Halaska, B., Lambourn, D.M., Lehnhart, T., Urbán R., J., Rowles, T., Rice, J., Savage, K., Wilkinson, K., Greenman, J., Viezbicke, J., Cottrell, B., Goley, P.D., Martinez, M., Fauquier, D., 2024. Gray whale (Eschrichtius robustus) post-mortem findings from December 2018 through 2021 during the Unusual Mortality Event in the Eastern North Pacific. PLoS One 19, e0295861. https://doi.org/10.1371/journal.pone.0295861
Stimmelmayr, R., Gulland, F.M.D., 2020. Gray Whale (Eschrichtius robustus) Health and Disease: Review and Future Directions. Front. Mar. Sci. 7. https://doi.org/10.3389/fmars.2020.588820
Takeda, M., Ogino, M., n.d. Record of a Whale Louse, Cyamus scammoni Dall (Crustacea: Amphipoda: Cyamidae), from the Gray Whale Strayed into Tokyo Bay, the Pacific Coast of Japan.
Von Duyke, A.L., Stimmelmayr, R., Sheffield, G., Sformo, T., Suydam, R., Givens, G.H., George, J.C., 2016. Prevalence and Abundance of Cyamid “Whale Lice” (Cyamus ceti) on Subsistence Harvested Bowhead Whales (Balaena mysticetus). Arctic 69, 331–340.
Würsig, B., Thewissen, J.G.M., Kovacs, K.M., 2017. Encyclopedia of Marine Mammals. Elsevier Science & Technology, Chantilly, UNITED STATES.
Baleen whales must navigate a seemingly featureless world to locate the resources they need to survive. The task of finding prey to feed on in the vast seascapes relies on the use of several sensory modalities that operate at different scales (Torres 2017; Figure 1). For example, baleen whale vision is believed to be rather limited, with the ability to see objects about 10-100 meters away. Yet, baleen whale somatosensory perception of oceanographic stimuli is thought to be on the order of 100-1000s kilometers. This diversity in sensory ability has led scientists to believe that whales, in fact all animals, perceive cues and make decisions at several scales. As ecologists, we endeavor to understand why and when animals are found (or not found) in certain locations as this knowledge allows us to better manage and conserve animal populations. With this information we can aim to minimize potential anthropogenic disturbance and protect important resource areas, such as foraging or nursing grounds. In order to accomplish this goal, we ourselves must conduct studies and test hypotheses at several scales (Levin 1992; Hobbs 2003). As someone who tackles spatial foraging ecology questions, I am particularly interested in understanding whale behavior and movement in the context of feeding. Since accurately measuring predator and prey distribution at the same scales can be challenging, we often resort to environmental variables to serve as proxies for prey, whereby we look for correlations between environmental variables and whales to understand and predict the distribution of our population.
Figure 1. Schematic of hypothetical interchange of sensory modalities used by baleen whales to locate prey at variable scales. X-axis represents log distance to prey from micro (left) to macro (right). Y-axis represents the relative use of each sensory modality between 0 (no contribution) to 10 (highest contribution). Each line and color represent a different sensory modality. Figure taken and caption adapted from Torres 2017.
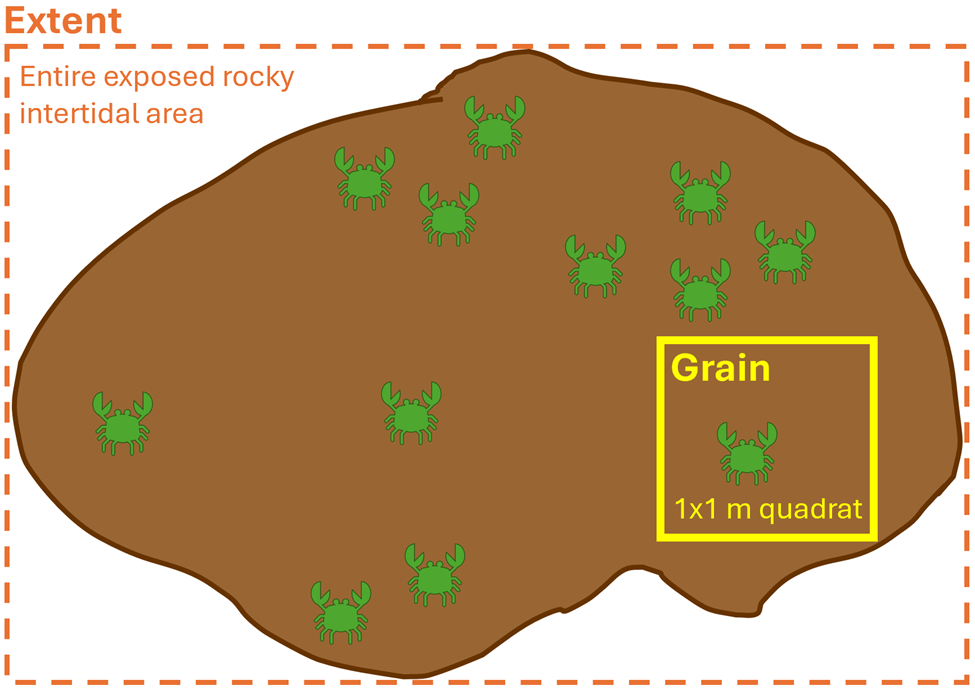
What do I mean when I use the word ‘scale’? The term scale is typically explained by two components: grain and extent (Wiens 1989). The grain is the finest resolution measured; in other words, how detailed we are measuring. The extent is the overall coverage of what we are measuring. These components can be applied to both spatial scale and temporal scale. For example, spatially, if we were using a 1×1 meter sampling quadrat to count the number of crabs on a rocky shore, then our grain would be the 1 m2 quadrat and the extent would be the entire exposed rocky intertidal area that we are surveying (Figure 2). Temporally, if we placed a temperature logger at the mouth of Yaquina Bay that took a temperature recording every minute for two years, then our grain would be one minute and the extent would be two years. So, when designing a study, it is imperative for us to decide on the spatiotemporal scales of the ecological questions we are asking and the hypotheses we are testing, as it will inform what data we need to collect. When making this decision, it is important to think about the scale at which the ecological process happens, as opposed to the scale at which we can observe the process (Levin 1992). In other words, we need to think from the perspective of our study species, as opposed to from our own human perspective. Making informed and ecologically reasonable decisions regarding the choice of scale relies on having prior knowledge of an animal’s biology, such as knowing that baleen whales might see a prey patch that is 50 meters away, but it may also somatosensorily perceive an oceanic front where zooplankton prey aggregate from 500 kilometers away.
Figure 2. Schematic of spatial scale where the extent (depicted by dashed orange box) is the entire exposed rocky intertidal area being surveyed and the grain (solid yellow box) is the 1×1 m quadrat being used to count crabs.
There is a wealth of studies that have explored space use patterns of wildlife relative to environmental variables to better understand foraging behavior. I want to share a couple from the marine mammal realm with you that I find particularly fascinating. In their 2018 study, González García and colleagues used opportunistic sightings of blue whales around the Azorean islands of Portugal and modeled their distribution patterns relative to physiographic and oceanographic variables summarized at different spatial (fine [1-10 km] and meso [10-100 km]) and temporal (daily, weekly, monthly) scales. The two variables that were most correlated with blue whale occurrence was distance from the coast and eddy kinetic energy (a measure of mesoscale variability of ocean dynamics). Both of these variables were interestingly found to be scale invariant, meaning that no matter which spatial and temporal scale was investigated, the relationship between blue whales and these two variables stayed the same; blue whale occurrence increased with increasing distance from the coast and was maximal at an eddy kinetic energy value of 0.007 cm2/s2 (Figure 3).
Figure 3. Functional response curves between presence of Azorean blue whales and distance to the coast (panel 1 on left) and eddy kinetic energy (panel 2 on right). The top row of each panel represents the low spatial scale and the bottom row represents the high spatial scale. Each column represents a different temporal scale (from left to right: daily, weekly, monthly). Note that the general shape of the relationship remains similar across all spatiotemporal scales and that the peak of the curves tend to occur at the same values for distance to coast and eddy kinetic energy across all scales. Figures taken from González García et al. 2018.
However, not all studies find scale invariant relationships. For example, Cotté and co-authors (2009) found that habitat use of Mediterranean fin whales was very much scale dependent. At a large scale (700-1,000 km and annual), fin whales were more densely aggregated during the summer in the Western Mediterranean where there was consistently colder water than in the winter. However, at a meso scale (20-100 km and weekly-monthly), fin whale densities were highest in areas where there were steep changes in temperature, as opposed to consistently cold temperatures. The authors explain that these differences in fin whale density and temperature at different scales are likely due to whale movement being driven by annually persistent prey abundance at the large scale, but at the meso scale, where prey aggregations are less predictable, the fin whales’ distribution becomes more driven by areas of physical ocean mixing.
As I investigate the environmental drivers of individual gray whale space use using our 8-year GRANITE (Gray whale Response to Ambient Noise Informed by Technology and Ecology) dataset, these studies (and many more) are at the top of my mind to interpret the patterns we are detecting. Our goal is to quantify and describe what environmental conditions (1) lead to a higher probability of a gray whale being seen in our central Oregon coast study area (~70 km) at a daily scale, and (2) influence space use patterns (activity range, residency, activity center) of different individual whales at annual scales. Our results show both consistency and variation in the environmental drivers of gray whales across these scales, leading me to deeply consider how gray whales make decisions at different points in their lives, based on information gained through various senses, to maximize their chances of capturing food. Previous work from the GEMM Lab on the relationships between gray whales and prey, at both fine (read more here) and large (read more here) scales have guided my work by providing specific hypotheses regarding environmental variables and lag times for me to test. Investigating the environmental drivers of animal space use and behavior is exciting work as it reveals that no single environmental variable determines animal distribution, but rather that multiple processes are happening concomitantly that animals respond to at different scales continually. It is only by studying animal space use patterns across spatiotemporal scales that we can begin to understand their complex decision-making patterns.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog below and get a monthly message when we post a new blog. Just add your name and email into the subscribe box below.
References
Cotté, C., Guinet, C., Taupier-Letage, I., Mate, B., & Petiau, E. (2009). Scale-dependent habitat use by a large free-ranging predator, the Mediterranean fin whale. Deep Sea Research Part I: Oceanographic Research Papers, 56(5), 801-811.
González García, L., Pierce, G. J., Autret, E., & Torres-Palenzuela, J. M. (2018). Multi-scale habitat preference analyses for Azorean blue whales. PLoS One, 13(9), e0201786.
Hobbs, N. T. (2003). Challenges and opportunities in integrating ecological knowledge across scales. Forest Ecology and Management, 181(1-2), 223-238.
Levin, S. A. (1992). The problem of pattern and scale in ecology: the Robert H. MacArthur award lecture. Ecology, 73(6), 1943-1967.
Torres, L. G. (2017). A sense of scale: Foraging cetaceans’ use of scale‐dependent multimodal sensory systems. Marine Mammal Science, 33(4), 1170-1193.
Wiens, J. A. (1989). Spatial scaling in ecology. Functional ecology, 3(4), 385-397.
As the sun set on February 16th, the R/V Star Keys pulled into Wellington Harbour, marking the end of the 2025 SAPPHIRE field season. The crew and science team returned to shore after a packed, productive, and successful three weeks at sea studying the impacts of environmental change on blue whales and krill in the South Taranaki Bight, Aotearoa New Zealand.
A blue whale comes up for air in the South Taranaki Bight.
In stark contrast to the 2024 field season, which featured dense and seemingly endless layers of gelatinous salps in the water and no krill or blue whales in the South Taranaki Bight, the 2025 field season was filled with blue whales and krill. In our three weeks aboard our research vessel Star Keys this year, we observed 66 blue whales, most of which were lunge feeding at the surface on dense patches of krill. We also collected krill for on-board respiration experiments and to be frozen to measure their lengths, weights, and caloric content. We recovered two hydrophones that recorded blue whale calls for the past year, and replaced them with two more. We collected identification photos, skin and blubber tissue samples for genetic and hormone analysis, and flew drones over almost all whales we encountered to measure body condition and morphology. We conducted water column profiles to measure the oceanography of the region, and mapped the prey field as we surveyed using a scientific echosounder.
Map of our survey effort (gray tracklines), blue whale sightings (red circles), and hydrophone locations (purple stars).
Around the world, we are currently bearing witness to environmental change. Our survey last year in 2024 was a reminder of the challenges these blue whales face to survive and thrive in an increasingly unpredictable ocean. This year was a poignant example of the vibrant marine life that exists here in the South Taranaki Bight when ocean conditions align more closely with what is expected, and of the incredible resilience of these animals as they navigate changing waters. These contrasting conditions over multiple years are key to our understanding as we study the impacts of climate change on krill and blue whales through the SAPPHIRE project.
Drone image of a blue whale coming to the surface.
The fieldwork we do to collect these data is motivated by scientific questions, management needs, and fascination with this ecosystem. But ultimately, what makes fieldwork possible and memorable is the people. We are deeply grateful for the many partners on the SAPPHIRE project. The 2025 science team was made up of Leigh Torres, Dawn Barlow, KC Bierlich, Kim Bernard (Oregon State University), Mike Ogle, and Ros Cole (Department of Conservation). The outstanding crew of the R/V Star Keys (Western Work Boats), Josh Fowden, Dave Futter, and Jordy Maiden-Drum, kept us safe, sailing, fed, and happy for three intense weeks. We are also grateful for our shore support, including our colleagues at Cornell University’s Yang Center for Conservation Bioacoustics, NIWA, the Marine Mammal Institute at Oregon State University, and the University of Auckland. Importantly, we deeply appreciate our many stakeholders who help us share, learn, and make our findings meaningful, including the Department of Conservation, the people of Aotearoa, and iwi across our study region, especially Ngāruahine who hosted us at the Rangatapu Marae for a profound hui with a powerful pōwhiri and critical wānanga of knowledge sharing.
Drone image of a blue whale mom and calf pair.
Now the next phase of the work begins. We have many terabytes of data to process, analyze, interpret, and share. We will certainly have our hands full. But while we are at our computers back in Oregon, we will be holding the memories of this field season close: The brilliant turquoise glow of a blue whale just below the surface, the sound of the deep exhalation as the whale comes up for air, and the awe of looking into a blue whale’s eye as it engulfs a dense swarm of krill; The golden sunset lighting and moon rise over Cape Farewell, and Mount Taranaki towering over the blue waters of the South Taranaki Bight; The giddy exclamations or silent awe of those of us privileged to spend time in these waters observing these animals, and the visions that linger just behind our eyelids as we fell into an exhausted sleep. We will see what the next year holds for the SAPPHIRE team and the blue whales and krill of the South Taranaki Bight.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly alert when we make a new post! Just add your name into the subscribe box below!