Clara Bird, PhD Student, OSU Department of Fisheries, Wildlife, and Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
The start of a new school year is always an exciting time. Like high school, it means seeing friends again and the anticipation of preparing to learn something new. Even now, as a grad student less focused on coursework, the start of the academic year involves setting project timelines and goals, most of which include learning. As I’ve been reflecting on these goals, one of my dad’s favorite sayings has been at the forefront of my mind. As an overachieving and perfectionist kid, I often got caught up in the pursuit of perfect grades, so the phrase “just learn the stuff” was my dad’s reminder to focus on what matters. Getting good grades didn’t matter if I wasn’t learning. While my younger self found the phrase rather frustrating, I have come to appreciate and find comfort in it.
Given that my research is focused on behavioral ecology, I’ve also spent a lot of time thinking about how gray whales learn. Learning is important, but also costly. It involves an investment of energy (a physiological cost, Christie & Schrater, 2015; Jaumann et al., 2013), and an investment of time (an opportunity cost). Understanding the costs and benefits of learning can help inform conservation efforts because how an individual learns today affects the knowledge and tactics that the individual will use in the future.
Like humans, individual animals can learn a variety of tactics in a variety of ways. In behavioral ecology we classify the different types of learning based on the teacher’s role (even though they may not be consciously teaching). For example, vertical transmission is a calf learning from its mom, and horizontal transmission is an individual learning from other conspecifics (individuals of the same species) (Sargeant & Mann, 2009). An individual must be careful when choosing who to learn from because not all strategies will be equally efficient. So, it stands to reason than an individual should choose to learn from a successful individual. Signals of success can include factors such as size and age. An individual’s parent is an example of success because they were able to reproduce (Barrett et al., 2017). Learning in a population can be studied by assessing which individuals are learning, who they are learning from, and which learned behaviors become the most common.
An example of such a study is Barrett et al. (2017) where researchers conducted an experiment on capuchin monkeys in Costa Rica. This study centered around the Panama ́fruit, which is extremely difficult to open and there are several documented capuchin foraging tactics for processing and consuming the fruit (Figure 1). For this study, the researchers worked with a group of monkeys who lived in a habitat where the fruit was not found, but the group included several older members who had learned Panamá fruit foraging tactics prior to joining this group. During a 75-day experiment, the researchers placed fruits near the group (while they weren’t looking) and then recorded the tactics used to process the fruit and who used each tactic. Their results showed that the most efficient tactic became the most common tactic over time, and that age-bias was a contributing factor, meaning that individuals were more like to copy older members of the group.
Figure 1. Figure from Barrett et al. (2017) showing a capuchin monkey eating a Panamá fruit using the canine seam technique.
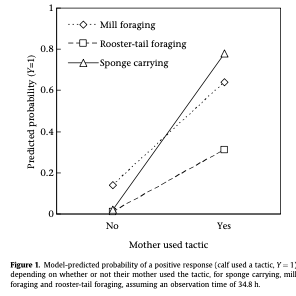
Social learning has also been documented in dolphin societies. A long-term study on wild bottlenose dolphins in Shark Bay, Australia assessed how habitat characteristics and the foraging behaviors used by moms and other conspecifics affected the foraging tactics used by calves (Sargeant & Mann, 2009). Interestingly, although various factors predicted what foraging tactic was used, the dominant factor was vertical transmission where the calf used the tactic learned from its mom (Figure 2). Overall, this study highlights the importance of considering a variety of factors because behavioral diversity and learning are context dependent.
Figure 2. Figure from Sargeant & Mann (2009) showing that the probability of a calf using a tactic was higher if the mother used that tactic.
Social learning is something that I am extremely interested in studying in our study population of gray whales in Oregon. While studies on social learning for such long-lived animals require a longer study period than of the span of our current dataset, I still find it important to consider the role learning may play. One day I would love to delve into the different factors of learning by these gray whales and answer questions such as those addressed in the studies I described above. Which foraging tactics are learned? How much of a factor is vertical transmission? Considering that gray whale calves spend the first few months of the foraging season with their mothers I would expect that there is at least some degree of vertical transmission present. Furthermore, how do environmental conditions affect learning? What tactics are learned in good vs. poor years of prey availability? Does it matter which tactic is learned first? While the chances that I’ll get to address these questions in the next few years are low, I do think that investigating how tactic diversity changes across age groups could be a good place to start. As I’ve discussed in a previous blog, my first dissertation chapter will focus on quantifying the degree of individual specialization present in my study group. After reading about age-biased learning, I am curious to see if older whales, as a group, use fewer tactics and if those tactics are the most energetically efficient.
The importance of understanding learning is related to that of studying individual specialization, which can allows us to estimate how behavioral tactics might change in popularity over time and space. We could then combine this with knowledge of how tactics are related to morphology and habitat and the associated energetic costs of each tactic. This knowledge would allow us to estimate the impacts of environmental change on individuals and the population. While my dissertation research only aims to provide a few puzzle pieces in this very large and complicated gray whale ecology puzzle, I am excited to see what I find. Writing this blog has both inspired new questions and served as a good reminder to be more patient with myself because I am still, “just learning the stuff”.
Today let me take you on a journey into the tropical waters of the Indo-Pacific Ocean, far from Oregon’s beautiful coasts. Although I have been working as a postdoc on the OPAL project for a year, the pandemic has prevented me from moving to the US as planned. Like so many around the globe, I have been working remotely from my study area (Oregon coastal waters), imagining my study species (blue, fin and humpback whales) gently swimming and feeding along the productive California Current system. One day, I’ll get to see these amazing animals for real, that’s for sure.
But in the meantime, I have taken this year as an opportunity to work with the GEMM lab, while continuing to enjoy the marvels of New Caledonia, a French overseas territory where I have lived for more than 6 years now. Among the animals that I get to approach and observe regularly in the coral reef lagoons that surround the island, the dugong (Dugon dugon) is perhaps the most emblematic and intriguing. This marine mammal is listed as vulnerable in the IUCN Red list of threatened species and has been the focus of important research and conservation efforts in New Caledonia over the last two decades1–3. During my previous post-doctoral position at the French Institute of Research for Sustainable Development, I contributed to some recent research involving satellite tracking of dugongs in the region. This work has led to a publication, now in review4, and will be the topic of my oral presentation at the 7th International Bio-Logging Science Symposium hosted in Hawaii in a couple weeks.
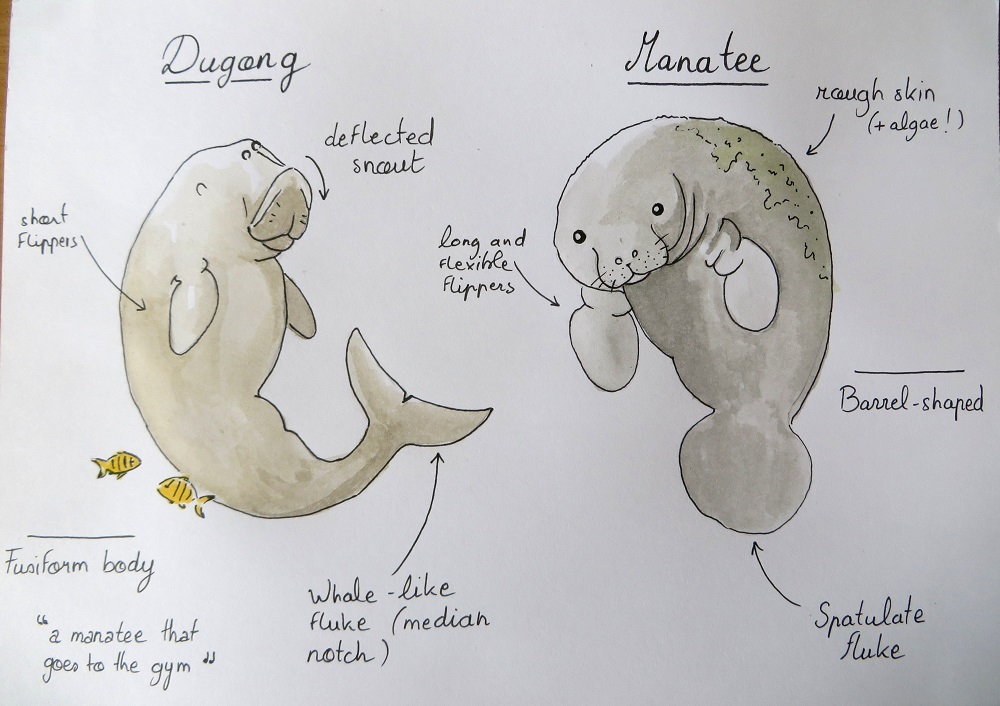
While I was analyzing dugong satellite tracks, writing this paper with my colleagues and preparing for the symposium, I learned a lot about these strange “sea cows”. Dugongs belong to the Sirenian marine mammal order, just like manatees (West Indian, Amazonian and West African species), which they are often mistaken for (watch out: Google Images will misleadingly suggest hundreds of manatee pictures if you make a “dugong” keyword search). The physiology and anatomy of dugongs is actually quite different from that of manatees (Figure 1). They also live in a different part of the world as they are broadly distributed in the Indo-Pacific coastal and island waters. Dugongs form separate populations, some of which are very isolated and at high risk of extirpation. They are found in 37 different countries, with Australia being home to the largest populations by far (exceeding 70,000 individuals5).
Figure 1: Manatee vs Dugong, can you tell them apart? Among other things, dugongs and manatees have a very different body shape. As the famous Sirenian specialist Helene Marsh said, a dugong essentially looks like “a manatee that goes to the gym”5! Illustration by S. Derville.
Sea cow or sea elephant?
Through the tree of evolution, the dugong and manatee’s closest relative is not the one you would think… other marine mammals like cetaceans or pinnipeds. Indeed, molecular genetic analyses have placed the Sirenians in the Afrotheria Superorder of mammals. Therefore, it appears that dugongs are more closely related to elephant and golden moles than to whales and dolphins!
As a memory aid to help remember this ancient origin, we may notice that both elephants and dugongs have tusks. Mature male and female dugongs have erupted tusks, although the females’ only erupt rarely and at a very old age. Interestingly, tusks are used by scientists to determine age. Analyses of growth layers in bisected dugong tusks have revealed that dugongs are long-lived, with a maximum longevity record of 73 years (estimated from a female individual found in Western Australia5).
An (almost) vegetarian marine mammal
Dugongs and manatees are the only predominantly herbivorous aquatic mammals. Given that manatees use both marine and fresh water ecosystems they tend to have a broader diet, eating many kinds of submerged, floating or emergent algae and seagrass (even bank growth!). On the other hand, dugongs are a strictly marine species and primarily feed on seagrass, which may look very similar to seaweeds, but are in fact marine flowering plants. Seagrass tend to form underwater shallow meadows that are among the most productive ecosystems in the world6. In fact, dugong grazing influences the biomass, species composition and nutritional quality of seagrass meadows7,8. Just like we take care of our gardens, dugongs regulate seagrass ecosystems. But there is more. Recent research conducted in the Great Barrier Reef indicates that seagrass seeds that have been digested by dugongs germinate at a faster rate9. As well as playing a role in dispersal10, it appears that dugongs are pooping seeds with enhanced germination potential, hence participating to seagrass meadow resilience.
Figure 2: Dugong mother and calf feeding on a dense seagrass bed (a) and solitary adult foraging in a very sparce seagrass bed (b). Seagrass grows in many different types of meadows, which may vary in density, species composition and substrate. For instance, seagrass species of the Halophila genus are among the preferred dugong’s meals although may be very thinly distributed (c). Photo credit: Serge Andréfouët, New Caledonia.
Unlike manatees, dugongs cannot feed over the whole water column and are strictly bottom feeders. They use their deflected snout (Figure 1) to search the seabed for their favorite food (Figure 2). The feeding trails left by dugongs in dense seagrass meadows are easily detectable from above, just like the sediment clouds that they generate when searching muddy bottoms. Although seagrass is undoubtedly the main component of the dugong’s diet, they may incidentally (or not) ingest algae and invertebrates5.
A legendary animal
The etymology for the word Sirenian comes from the mermaids, or “sirens” of the Greek mythology. These aquatic creatures with the upper body of a female human would sing to lure sailors towards the shore… and towards a certain death. The morphology of dugongs and manatees shares some resemblance with mermaids, at least enough for desperate and lonely sailors to think so!
In addition to having a scientific name rooted in legends, dugongs are also important to contemporary human cultures. In tropical islands and coastal communities, marine megafauna species such as dugongs are considered heritage, due to the strong bond that their people have forged with the ocean5. Dugongs may play an important cultural role because they can be part of the socio-symbolic organization of societies, associated with the imaginary world, or simply because they are seen as companions of the sea, which people frequently encounter. For New Caledonia’s indigenous people, the Kanaks, dugongs can be totem to tribes. Like other large marine species (whales, sharks), the dugong is also considered as an embodiment of ancestors11.
Dugongs have been hunted throughout their range since prehistoric times. Archaeological excavations such as those conducted on the island of Akab in the United Arab Emirates12, indicate that dugong hunting played a role in ancient rituals, in addition to providing a large quantity of meat. The cultural value of dugongs is recognized by multiple countries, which have therefore authorized indigenous dugong hunting, sometimes under quotas. For instance, in Australia, dugongs may be legally hunted by Aboriginal and Torres Strait Islander people (Figure 3) under section 211 of the Native Title Act 1993.
In New Caledonia, the dugong has been protected since 1962 and its hunting is only authorized in one province, with a dispensation for traditional Kanak celebrations13. However, in view of the critical situation in which the New Caledonian dugong population finds itself, estimated at around 700 individuals in 2008-201214, no hunting exemptions have been issued since 2004.
Figure 3: “Naath” (dugong hunting platform), hand colored linocut by Torres Strait Islander artist Dennis Nona. The art piece represents traditional dugong hunting where the hunter is guided by the phosphorescent glow the dugong would leave in the water at night.
What future for dugongs?
Despite legislations to forbid dugong meat consumption outside specific traditional permits, poaching persists, in New Caledonia and in many of the “low-income” countries that are home to dugongs. As climate change and demography intensifies risks to food security, scientists and stakeholders fear for dugongs. Moreover, dugongs entirely rely on seagrass ecosystems that are also disappearing at an alarming rate (7% per year6) as a result of coastal development, pollution and overfishing.
Can we preserve dugongs in regions of high climate vulnerability and where people still have low levels of access to basic needs? Can dugongs play the role of “umbrellas” for the conservation of the ecosystem they live in? I do not have the answer to these questions but I certainly believe that people’s well-being and environmental conservation are tightly intertwined. I hope that rising transdisciplinary approaches such as those supported by the “One Health” framework will help reconnect human populations to their environment, and achieve the goal of optimal health for everyone, humans and animals.
References
1. Garrigue, C., Patenaude, N. & Marsh, H. Distribution and abundance of the dugong in New Caledonia, southwest Pacific. Mar. Mammal Sci. 24, 81–90 (2008).
2. Cleguer, C., Grech, A., Garrigue, C. & Marsh, H. Spatial mismatch between marine protected areas and dugongs in New Caledonia. Biol. Conserv. 184, 154–162 (2015).
3. Cleguer, C., Garrigue, C. & Marsh, H. Dugong (Dugong dugon) movements and habitat use in a coral reef lagoonal ecosystem. Endanger. Species Res. 43, 167–181 (2020).
4. Derville, S., Cleguer, C. & Garrigue, C. Ecoregional and temporal dynamics of dugong habitat use in a complex coral reef lagoon ecosystem. Sci. Rep. (In review)
5. Marsh, H., O’Shea, T. J. & Reynolds, J. E. I. Ecology and conservation of the Sirenia: dugongs and manatees, Vol 18. (Cambridge University Press, Cambridge, 2011).
6. Unsworth, R. K. F. & Cullen-Unsworth, L. C. Seagrass meadows. Curr. Biol. 27, R443–R445 (2017).
7. Aragones, L. V., Lawler, I. R., Foley, W. J. & Marsh, H. Dugong grazing and turtle cropping: Grazing optimization in tropical seagrass systems? Oecologia 149, 635–647 (2006).
8. Preen, A. Impacts of dugong foraging on seagrass habitats: observational and experimental evidence for cultivation grazing. Mar. Ecol. Prog. Ser. 124, 201–213 (1995).
9. Tol, S. J., Jarvis, J. C., York, P. H., Congdon, B. C. & Coles, R. G. Mutualistic relationships in marine angiosperms: Enhanced germination of seeds by mega-herbivores. Biotropica (2021) doi:10.1111/btp.13001.
10. Tol, S. J. et al. Long distance biotic dispersal of tropical seagrass seeds by marine mega-herbivores. Sci. Rep. 7, 1–8 (2017).
11. Dupont, A. Évaluation de la place du dugong dans la société néo-calédonienne. (Mémoire Master. Encadré par L. Gardes (Agence des Aires Marines Protégées) et C. Sabinot (IRD), 2015).
12. Méry, S., Charpentier, V., Auxiette, G. & Pelle, E. A dugong bone mound: The Neolithic ritual site on Akab in Umm al-Quwain, United Arab Emirates. Antiquity 83, 696–708 (2009).
13. Leblic, I. Vivre de la mer, vivre de la terre… en pays kanak. Savoirs et techniques des pêcheurs kanak du sud de la Nouvelle-Calédonie. (Société des Océanistes, 2008).
14. Hagihara, R. et al. Compensating for geographic variation in detection probability with water depth improves abundance estimates of coastal marine megafauna. PLoS One 13, e0191476 (2018).
Returning to a place you once lived always shows how much you and the world around you have changed, offering a new perspective on the time away and where you are now. I’m writing this from my old office at Bigelow Laboratory for Ocean Sciences in East Boothbay, Maine, where I worked before moving out to Oregon to join the GEMM Lab and start graduate school at OSU. Being back in Maine has made me reflect on how much I’ve learned over the last year, and given me the opportunity to think about what’s ahead.
As a science communications specialist at Bigelow for three years, much of my work involved quickly getting up to speed on new research and writing articles for a general audience about important ocean processes. My first year of grad school has both deepened and broadened my perspective on the ocean, prodding me to think at telescoping temporal and spatial scales. I can tell that I think about the ocean differently now.
In 2019, writing this feature-length article about impacts of changing climate on North Atlantic right whales and their prey was my first introduction to research using environmental models to help mitigate entanglement issues. Now, I’m excited to be pursuing research with these themes as part of the GEMM Lab’s Project OPAL.
Over the last year, my coursework in ocean ecology and biogeochemistry surveyed the physical and chemical workings of the ocean, marine ecosystem dynamics, and the global cycles that control much of life on earth. Through lab activities and fieldwork, I began learning about whales and the marine system off the coast of Oregon, and how to ask questions that occupy the intersection between whales and their environment.
This work and learning have made me think in a new way about whales as agents of biogeochemical cycling: how do they shuttle nutrients across large distances and affect global cycles? In what ways is the biogeography of whales an expression of the global patterns of light availability and nutrient fluxes that support their prey? How is it possible to detangle and encapsulate all of the relevant variability of a natural system into a mathematical model?
All these questions were churning in my mind at the start of this trip, as I spent the bus ride from Boston to Maine reading papers for our monthly GEMM lab meeting. I also remembered the first meeting that I joined, when I was so intimidated that I couldn’t imagine discussing research with this impressive group. This time, I was just as in awe as ever of the lab, but a bit more confident in wielding acronyms and sharing ideas.
I actually attended my first GEMM Lab meeting while still working in Maine, in July 2020. I was joined by my friends’ one-year-old daughter, who alternately tried to chime in on the meeting and shut my laptop. Now, she is a chatty two-year-old kid and newly a big sister. The new baby became part of my PhD this week too, snoozing in my lap as I edited an abstract.
Only 16 days old and already helping write an abstract!
Often, it’s only seeing my friends’ children grow that shows me how much time has passed. This time, I can feel it in myself, as well. I’m excited to have made it through the first year of coursework and to be learning to formulate research questions and think about ocean systems in new ways. I’m happy to be back in this place that inspired me to pursue a PhD, and to be able to share my own work and knowledge with former colleagues.
I gained so much during my time here at Bigelow: the communication and outreach skills in my job, inspiration from the scientific curiosity and passion of my colleagues, and the support of all these people who reassured me that I would get into grad school and that doing a PhD is a good idea. I’m so happy to be able to carry this support and momentum forward with me through the rest of grad school, and excited to return to Oregon and keep going.
By Allison Dawn, Master’s student, OSU Department of Fisheries, Wildlife and Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
Part 1: Team “Heck Yeah!”
Allison, inspecting a prey sample from the morning’s in situ collection:
“Look! It’s dungeness crab larvae.”
Damien, squinting to see the tiny megalopa floating in the water:
“Oh, heck yeah!”
This simple exchange is just a snapshot of the shared excitement the 2021 Port Orford project interns and I have felt since we have arrived at the field station. Whether it was the first time we set up the theodolite correctly, watched our first GoPro footage of the water column, or when we jumped from the dinner table to grab the camera after Lisa spotted a whale foraging in Mill Rocks – this year’s team is full of enthusiasm. Thus, I’m excited to introduce you all to the 2021 summer field team name: Team “Heck Yeah!”.
While the name is unconventional, the phrase “heck yeah” is not just an exclamation we use when excited. We also use it when we know a task will be a challenge to learn, like staying within the correct GPS position for kayak sampling stations during windy conditions. We know some tasks will be difficult, but we now say “heck yeah!” to meet challenges with a focused, positive determination. This positivity in the face of challenges is a great skill to have, especially because these first two weeks have been one steep learning curve for all of us.
If you read my previous blog, you know that I (Allison) am taking over leadership of the Port Orford project next summer. On top of learning the important day-to-day data collection tasks along with the interns, I have also been learning from Lisa how to lead this project. I am very grateful to have her here to guide me, as I have been able to witness that even when you are a seasoned project leader (this is Lisa’s 4th year!) curve balls can and do happen. It has been great to watch as she has adapted to each unexpected challenge so far, and I am learning first-hand how to best prepare for the Port Orford field season – preparation that includes expecting the unexpected.
Before I discuss in more detail the adaptations we have made and what we are seeing so far in the field, I would like to introduce the interns that comprise our JASPER program this year. Adaptability is a key skill to have when dealing with the unexpected, and after two weeks of training in abnormally windy conditions, Team “Heck Yeah!” is ready for this field season. Though it has only been two weeks, I already feel confident it will continue to be a great season and am grateful to have each one of them aboard our team.
Part 2: The 2021 Port Orford Gray Whale Foraging Ecology Team
First, I would like to introduce Jasen White. Jasen is a first generation, non-traditional undergraduate student at Oregon State University. He is a senior majoring in Fisheries and Wildlife Science and has a passion for spatial ecology and resource management. His professional management background in the U.S. Navy combined with his technical skill set has made him a great addition to the team. He has proven to be dependable and has already brought some new ideas to the project, from handling/maintaining gear to task efficiency. While we have yet to see many whales, Jasen is most looking forward to honing his theodolite skills. As we all have learned, theodolite tracking is more involved than simply looking through a scope. I appreciate the motivation he and the other interns have to become experts in each task.
Figure 1. Jasen photographing “Moby Dick”, a PCFG whale familiar to the GEMM lab, who was spotted foraging near the jetty. Port Orford, OR. July 2021.
The next member of our intern team is Nadia Leal. Nadia is also an undergraduate at OSU and is majoring in Marine Mammal Biology and minoring in Marine Conservation Management. She is an aspiring conservation biologist and moved all the way from Nevada to study at Oregon State University. As a previous soccer athlete in high school, teamwork and determination are two of the many skills Nadia has contributed to the Port Orford team this year. Even during the most difficult training days, her positive energy has been infectious to everyone. So far, Nadia’s favorite aspect about living at the field station is how closely she gets to interact with the community. She is fully enjoying the unique networking opportunities here – this internship has allowed her to connect with the team at Port Orford Sustainable Seafood and with other like-minded interns, such as Maddie English, a student conducting research here as part of OSU’s 2021 Marine Studies Initiative.
Figure 2. Nadia using high-powered binoculars to systematically scan the sea for gray whales. Port Orford, OR. July 2021.
Last but not least is our high school intern, Damian Amerman-Smith. Damian is a rising senior at Pacific High School and plans to attend college fall 2022 to pursue a career in engineering. Due to his experience here so far, he is now considering applying to OSU! Damian is a Port Orford local and in addition to sharing his sense of humor and unique stories, he also has demonstrated curiosity, engagement, and attention to detail to every aspect of our work thus far. Damian’s favorite task at this point in the season is observing whale activity from the cliff. He picked up on nuances of the theodolite quickly and has enjoyed learning the various methods for systematic wildlife observation.
Figure 3. Damian learning how to use Pythagoras, the tracking software that accompanies the theodolite. Port Orford, OR. July 2021.
Part 3: Wind, Prey, and Looking Ahead
Now that you are familiar with the 2021 Port Orford team, it is time to discuss updates from the field! It has been particularly windy during the start of this season, and we also have had few whale encounters.
Flexibility during unfavorable weather conditions is important, and for that reason it has made the training period all that more intense. We only have two weeks to master our new skill set before we collect four weeks of standardized data, and therefore we must be sure to choose our days off wisely. Good weather days have been sparse, so we make sure they are used for field work. However, just because the forecast predicts poor conditions does not mean we immediately have the day off. Forecasts are not always accurate, and for the past week the predicted versus actual conditions were not aligned. Hence, we wake up at the usual time and often drive to the Port early to assess the weather conditions in person before deciding what is feasible that day.
Yet, with adaptability, determination, and a close eye on conditions, we have managed to persist with our kayak sampling and theodolite observations. Due to the collective persistence of Team “Heck Yeah!”, we have been able to collect exciting prey data.
Figure 4. Mysid swarm captured on GoPro video at Tichenor Cove, Station 4. Port Orford, OR. July 2021.
It is important to note, however, that increased winds may not be ultimately bad. As Dawn has written about before on the blog, winds often increase upwelling conditions, bringing nutrients to the surface and thus increase prey abundance. In fact, we have seen an uptick in prey abundance in the past few days. In particular, we have captured increased amounts of zooplankton prey at two sampling stations in Tichenor Cove. We have seen Dungeness crab larvae, mysid swarms, and we even counted 438 amphipods that were collected in our prey net last Friday!
When I look back at the past two weeks, I am amazed to recount everything we have learned: First Aid/CPR, kayak paddle & rescue, theodolite tracking, in situ kayak prey sampling, data management, as well as gray whale and zooplankton prey ID. I know I speak for all of us when I say that these training weeks were challenging yet rewarding, and we retained our eagerness to put our new skillset to task. The future is bright as we hope to see a less wind and more whales over the next four weeks. Until then, we are saying “Heck Yeah!” to each challenge and continue to refine our skills so we are prepared when weather conditions are favorable. Stay tuned for more updates from Team “Heck Yeah!”.
Figure 5. Team “Heck Yeah!” excited for the field season. Port Orford, OR. July 2021.
Recently, when expected to choose a wildlife species for behavioral observation for one of my Oregon State University graduate courses, I immediately chose harbor seals as my focus. Harbor seals (Fig 1) are an abundant species and in proximity to the Hatfield Marine Science Center (HMSC) (Steingass et al., 2019) where I will be spending much of my time this summer, making logistics easy. Studying pinnipeds (marine mammals with a finned foot, seals, walrus, and sea lions) is appealing due to their undeniably cute physique, floppy nature on land, and super agile nature in the water. I am working to iron out my methods for this study, which I hope to work through in this initial phase of my research project.
Figure 1. Harbor seal hauling out to rest on rocks off Oregon Coast near HMSC.
Behaviors:
At times it can appear that the most interesting harbor seal behaviors occur under water, and the haul out time is simply time for resting. During mating season, most adult seal behaviors take place in the water, such as the incredible vocal acoustics displayed by the males to attract the females (Matthews et al., 2018). However, I hypothesize that young pups can capitalize on haul out time by practicing becoming adults (while the adults are taking that time to rest) and therefore I plan to observe their haul out behaviors in their first summer of life. Specifically, I will document seal pup vocal behavior to evaluate how they are learning to use sound. I am beginning this study in late July, which is just after pupping season (Granquist et al., 2016). This should give me the opportunity to find pups along the Oregon coast near HMSC, so I intend to visit several locations where harbor seals are known to frequently haul out. Knowing that field work and animal behavior is unpredictable, there is no telling what behaviors I will observe on a given day, or if I will see seals at all. Some days I could come home with lots of seal data and great photos, and other days I could come home with little to report. This will be my first hurdle combined with my time limit (strictly completing this observation in the next five weeks). I intend to schedule at least eight hours of field observation at haul-out sites over the next two weeks and will adjust my schedule based on my success in data collection at that point.
Figure 2. Harbor Seals hauling out on rocks not too far from HMSC.
Timing:
Prior knowledge on harbor seal haul-out sites along the Oregon coast is clearly important for this project’s success, but I must also pay close attention to the tide cycles. During low tides, haul out locations are exposed and occupied by seals. When the tide is high, the seals are less likely to haul-out (Patterson et al., 2008). Furthermore, according to a recent study conducted on harbor seals residing on the Oregon coast, these seals spend on average 71% of their time in the water and will haul-out for the remainder of their time (Steingass et al., 2019). Therefore, it is crucial to maximize my observation time of hauled out pups wisely.
Concerning timing, I also need to observe locations and periods without too many tourists who can get near the haul-out site. As I learned recently, when children show up and start throwing rocks into the water near where harbor seals are swimming, the seals will recede from the area and no longer be available for observation. As an experiment, I waited for the noisy crowds with unchecked children to leave and only myself, my trusty sidekick (my daughter), and one quiet photographer were left on the beach. Once that happened, we noticed more and more seal heads popping up out of the water. Then they came closer and closer to the beach, splashing around doing somersaults visibly on the surface of the water. It was quite a show. I will either need to account for the presence of humans when evaluating seal behavior or assess only periods without disturbance. Seal pups are easily disturbed by humans, so I will keep a non-invasive distance while positioning myself to hear the vocals.
Figure 3. Hauled-out adult harbor seal on the Oregon coast near HMSC.
Data Collection and Analysis Approach:
The aspect of this project I am still working out is how to quantify pup vocalizations and their associated behaviors. As I mentioned, I will go out each week for eight hours and record each time I notice a pup exhibiting vocal behavior. I will categorize and describe the sound produced by the pup, and document any associated behavior of the pup or behavioral responses from nearby adult seals. Prior research has found that harbor seals are much attuned to vocal behavior. Mother harbor seals learn to quickly distinguish their own pup’s call within a few days of their birth (Sauve et al. 2015). I hypothesize that pups themselves can discern and use vocalizations, and I am excited to watch them develop over the course of my field observations.
Figure 4. Seal pup on the far-left rock, watching the adults as they appear to rest.
References
Granquist, S.M., & Hauksson, E. (2016). Seasonal, meteorological, tidal, and diurnal effects on haul-out patterns of harbour seals (Phoca vitulina) in Iceland. Polar Biology, 39 (12), 2347-2359.
Matthews, L.P., Blades, B., Parks, S. (2018). Female harbor seal (Phoca vitulina) behavioral response to playbacks of underwater male acoustic advertisement displays. PeerJ, 6, e4547.
Patterson, J., Acevedo-Gutierrez, A. (2008). Tidal influence on the haul-out behavior of harbor seals (Phoca vitulina) At all time levels. Northwestern Naturalist, 89 (1), 17-23.
Sauve, C., Beauplet, G., Hammil, M., Charrier, I. (2015). Mother-pup vocal recognition in harbour seals: influence of maternal behavior, pup voice and habitat sound properties. Animal Behavior, 105 (July 2015), 109-120
Steingass, S., Horning, M., Bishop, A. (2019). Space use of Pacific harbor seals (Phoca vitulina richardii) from two haulout locations along the Oregon coast. PloS one. 14 (7), e0219484.
By Marc Donnelly, Carleton College, GEMM Lab REU Intern
My name is Marc Donnelly (He/Him) and this summer I have the pleasure of working with Clara Bird and Dr. Leigh Torres on a project, within the GRANITE project, that maps the habitat use of Pacific Coast Feeding Group (PCFG) gray whales. This summer, as a National Science Foundation (NSF) Research Experience for Undergraduates (REU) intern, I have the opportunity to learn about the scientific process in action, build relationships with researchers, and pursue my own research project. I am relatively new to the world of research and even more green in the field of marine science. In September, I will start my fourth and last year as an Environmental Studies major at Carleton College in Minnesota, but for the next seven weeks my bread and butter is whales. I could not be more excited about the road ahead. I have read about gray whales, examined pictures of them, and even studied videos of the oblong beauties (Video 1). But the one thing I have not done, and probably will not do this summer, is see one… or a boat for that matter, or a single piece of field equipment. Not in person anyway. This is because I am a remote intern. And before I continue, let me clarify that I am not lamenting unfortunate circumstances. These past three weeks have already been enjoyable, fulfilling and exciting and I expect the summer will only get better. Yet, like with so many people in the past 16 months, my expected role has undergone some changes, so I want to take this opportunity to share my experience so far.
Video 1. Examples of gray whale behaviors observed from the drone
Especially when thinking about engaging with hands-on work, a fundamental aspect of any research program is place. REUs are competitive and sought after positions because they supply undergrads, who have to balance coursework with the desire to fully immerse themselves in a research community, with the opportunity to pursue a genuine research experience. Just being in a room surrounded by peers, grad students, and seasoned scientists who are all bubbling over with excitement and ideas is a fundamentally different (and might I add more motivating) experience than classroom lectures. Location is enough by itself to facilitate the connection between a burgeoning scientist and their research community as well as their work itself. Conducting hands-on fieldwork is also a common, highly sought-after aspect of an REU. However, visiting study sites, collecting data, and experiencing your study organism first hand, are all activities that become impossible when working remotely. So what do you do when you lack the benefits of location?
Well, if you had any sense, you would start by furnishing your apartment, finding a mattress that you can sleep on and a table and a chair that you can work at. But if you are like me, 21 and excited to be living away from home in a new city, you might be so overcome with the idea of adventure that you forget sleeping is important. Beyond furniture, the course of my summer has primarily been in the hands of my new mentors at Oregon State University (OSU), the institution funding my position in the GEMM lab, and thankfully they had a much more robust plan for my summer than I did. Data analysis and an in depth literature review have filled the void where my marine mammal companions could have been. This situation does not mean that analyzing data and diving into the literature are not part of in-person internships as well, or that I am not able to build any sort of connection with the gray whales I study, but my computer screen has certainly taken a more central role in my work. This summer, Geographic Information Systems (GIS) software is my weapon of choice. My goal is to create habitat maps of the coastal waters off Newport where gray whales feed that includes characteristics ranging from the type of surface on the bottom of the ocean (i.e., sand, reef, rock, etc) to more ephemeral features like kelp and prey density. This list of features I will map is dynamic based on the purpose and time scale of the map (month, year, static); so suffice it to say I will be making a lot of maps this summer. Once I have produced these habitat maps, the team and I can compare them with our whale sightings to better understand if and how gray whales use certain areas. This work will help us develop a baseline for gray whale ecology, which will ultimately be used to inform disturbance models and conservation efforts.
Figure 1: Me finally getting a table and chair for work.
After finding a way to move work online, the next step is to somehow engineer a social environment that provides people with a sense of community. As explained to the interns during our first professional development workshop, forming these connections are not just important for combating feelings of isolation, but they may also serve as fruitful professional relationships in the near or distant future. After three terms of online classes and vain attempts at forming meaningful connections via awkward breakout rooms and forced group projects, I was preemptively lowering my expectations for how this summer might unfold. I should not have worried; both the GEMM lab and the greater REU cohort have been extraordinary. It has been such a privilege and joy to meet so many compassionate and involved people. Every week there are numerous opportunities for interns to engage with various groups associated with OSU. From one-on-one meetings with my mentors to laid back “coffee breaks” with folks from Oregon Sea Grant, engrossing interactions abound. I even had the chance to attend the Marine Mammal Institute Monthly Meeting, or MMIMM, which for a newcomer to the world of marine science is both a fascinating and intimidating thing to watch unfold.
One of my favorite virtual gatherings of all was our monthly GEMM lab meeting, this month our activity involved brief presentations introducing ourselves and our research. If you are a fan of this blog and have had the opportunity to explore the happenings of the GEMM lab through this page, then you probably have some context to understand the excitement and curiosity I felt while listening about the current GEMM projects through my zoom screen. I was simultaneously humbled and comforted by the impressiveness of the work being undertaken by this group of researchers. Even though I was just being exposed to five-minute overviews of people’s work, it was daunting to compare my own limited knowledge to that of the other people on the call. Most of them have been studying marine megafauna for five years or more and their passion coupled with their grasp on their work was remarkable. I was also comforted by the descriptions of all these wonderful and intriguing projects because it gave me a sense of achievement. This feeling may sound silly, but just by virtue of being on a zoom call with such passionate scientists I felt relieved. Relieved because it seemed as though this community is what I had been working towards for the last few years. Not necessarily the GEMM lab in particular, but a community of inspiring people who care about each other, their work, and improving the world.
Despite the fact that they do not know what I look like from the shoulders down, my GEMM lab cohort has welcomed me into their midst and provided me with the tools and environment I need to connect and learn. I am grateful.
Dr. KC Bierlich, Postdoctoral Scholar, OSU Department of Fisheries, Wildlife, & Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
The morphology and body size of an animal is one of the most fundamental factors for understanding a species ecology. For instance, fish body size and fin shape can influence its habitat use, foraging behavior, prey type, physiological performance, and predator avoidance strategies (Fig 1). Morphology and body size can thus reflect details of an individual’s current health, likelihood of survival, and potential reproductive success, which directly influences a species life history patterns, such as reproductive status, growth rate, and energetic requirements. Collecting accurate morphological measurements of individuals is often essential for monitoring populations, and recent studies have demonstrated how animal morphology has profound implications for conservation and management decisions, especially for populations inhabiting anthropogenically-altered environments (Miles 2020) (Fig. 1). For example, in a study on the critically endangered European eel, De Meyer et al. (2020) found that different skulls sizes were associated with different ecomorphs (a local variety of a species whose appearance is determined by its ecological environment), which predicted different diet types and resulted with some ecomorphs having a greater exposure to pollutants and toxins than others. However, obtaining manual measurements of wild animal populations can be logistically challenging, limited by accessibility, cost, danger, and animal disturbance. These challenges are especially true for large elusive animals, such as whales that are often in remote locations, spend little time at the surface of the water, and their large size can preclude safe capture and live handling.
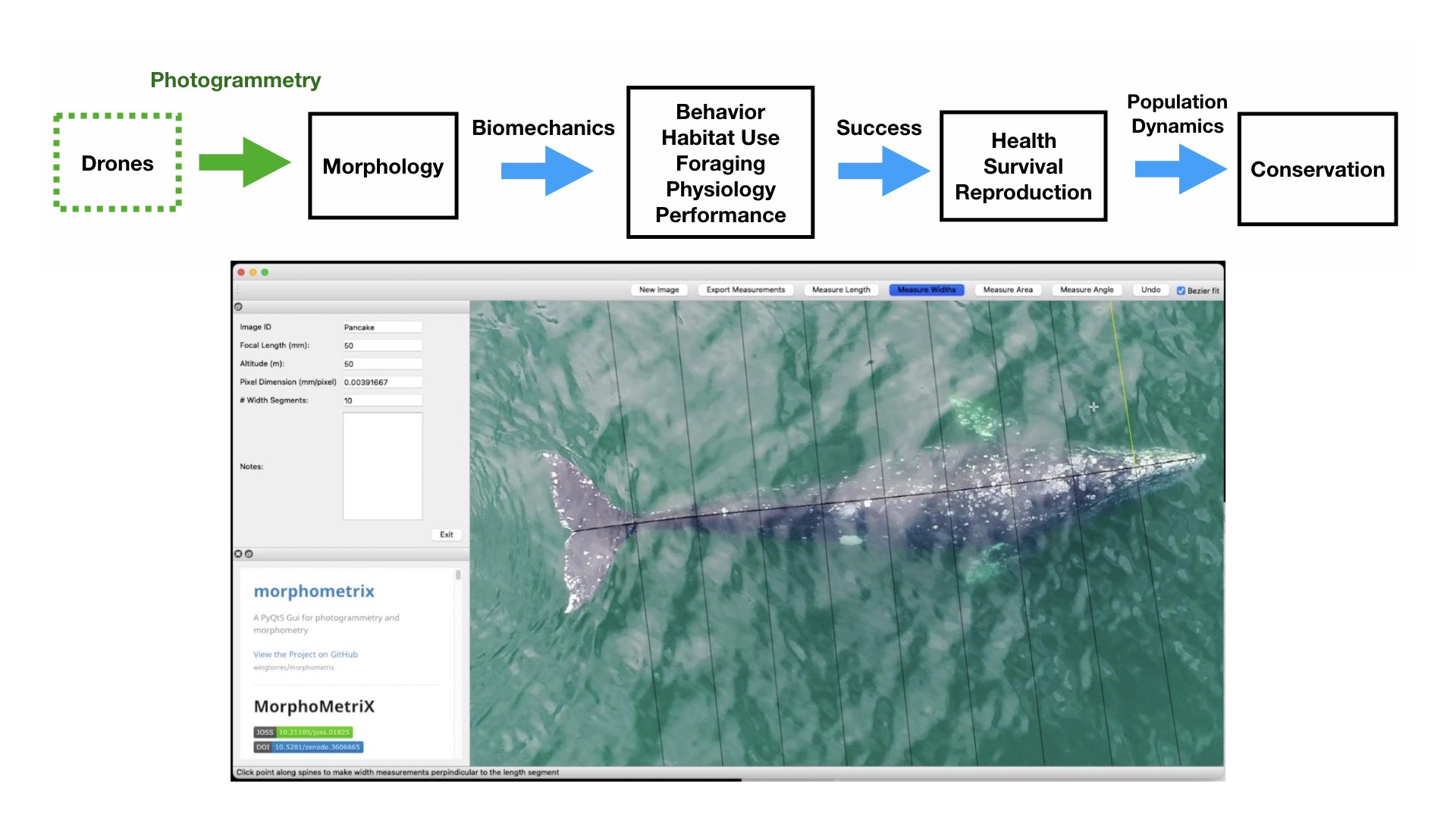
Figure 1. Top) A pathway framework depicting how the morphology of an animal influences its habitat use, behavior, foraging, physiology, and performance. These traits all affect how successful an animal is in its environment and can reflect an individual’s current health, likelihood of survival, and potential reproductive success. This individual success can then be scaled up to assess overall population health, which in turn can have direct implications for conservation. Bottom) an example of morphological differences in fish body size and fin shape from Walker et al. (2013). Fineness ratio (f) = length of body ÷ max body width.
Photogrammetry is a non-invasive method for obtaining accurate morphological measurements of animals from photographs. The two main types of photogrammetry methods used in wildlife biology are 1) single camera photogrammetry, where a known scale factor is applied to a single image to measure 2D distances and angles and 2) stereo-photogrammetry, where two or more images (from a single or multiple cameras) are used to recreate 3D models. These techniques have been used on domestic animals to measure body condition and estimate weight of dairy cows and lactating Mediterranean buffaloes (Negretti et al., 2008; Gaudioso et al., 2014) and on wild animals to measure sexual dimorphism in Western gorillas (Breuer et al., 2007), shoulder heights of elephants (Schrader et al., 2006), nutritional status of Japanese macaques (Kurita et al., 2012), and the body condition of brown bears (Shirane et al., 2020). Over 70 years ago, Leedy (1948) encouraged wildlife biologists to use aerial photogrammetry from aircraft for censusing wild animal populations and their habitats, where photographs can be collected at nadir (straight down) or an oblique angle, and the scale can be calculated by dividing the focal length of the camera by the altitude or by using a ratio of selected points in an image of a known size. Indeed, aerial photogrammetry has been wildly adopted by wildlife biologists and has proven useful in obtaining measurements in large vertebrates, such as elephants and whales.
Whitehead & Payne (1978) first demonstrated the utility of using aerial photogrammetry from airplanes and helicopters as a non-invasive technique for estimating the body length of southern right whales. Prior to this technique, measurements of whales were traditionally limited to assessing carcasses collected from scientific whaling operations, or opportunistically from commercial whaling, subsistence hunting, stranding events, and bycatch. Importantly, aerial photogrammetry provides a method to collect measurements of whales without killing them. This approach has been widely adopted to obtain body length measurements on a variety of whale and dolphin species, including bowhead whales (Cubbage & Calambokidis, 1987), southern right whales (Best & Rüther, 1992), fin whales (Ratnaswamy and Wynn, 1993), common dolphins (Perryman and Lynn, 1993), spinner dolphins (Perryman & Westlake 1998), and killer whales (Fearnbach et al. 2012). Aerial photogrammetry has also been used to measure body widths to estimate nutritive condition related to reproduction in gray whales (Perryman and Lynn, 2002) and northern and southern right whales (Miller et al., 2012). However, these studies collected photographs from airplanes and helicopters, which can be costly, limited by weather and infrastructure to support aircraft research efforts and, importantly, presents a potential risk to wildlife biologists (Sasse 2003).
The recent advancement and commercialization of unoccupied aircraft systems (UAS, or drones) has revolutionized the ability to obtain morphological measurements from high resolution aerial photogrammetry across a variety of ecosystems (Fig. 2). Drones ultimately bring five transformative qualities to conservation science compared to airplanes and helicopters: affordability, immediacy, quality, efficiency, and safety of data collection. Durban et al. (2015) first demonstrated the utility of using drones for non-invasively obtaining morphological measurements of killer whales in remote environments. Since then, drone-based morphological measurements have been applied to a wide range of studies that have increased our understanding on different whale populations. For example, Leslie et al. (2020) used drone-based measurements of the skull to distinguish a unique sub-species of blue whales off the coast of Chile. Groskreutz et al. (2019) demonstrated how long-term nutritional stress has limited body growth in northern resident killer whales, while Stewart et al. (2021) found a decrease in body length of North Atlantic Right whales since 1981 that was associated with entanglements from fishing gear and may be a contributing factor to the decrease in reproductive success for this endangered population.
Drone imagery is commonly used to estimate the body condition of baleen whales by measuring the body length and width of individuals. Recently, the GEMM Lab used body length and width measurements to quantify intra- and inter-seasonal changes in body condition across individual gray whales (Lemos et al., 2020). Drones have also been used to measure body condition loss in humpback whales during the breeding season (Christiansen et al., 2016) and to compare the healthy southern right whales to the skinnier, endangered North Atlantic right whales (Christiansen et al., 2020). Drone-based assessments of body condition have even been used to measure how calf growth rate is directly related to maternal loss during suckling (Christiansen et al., 2018), and even estimate body mass (Christiansen et al., 2019).
Drone-based morphological measurements can also be combined with whale-borne inertial sensing tag data to study the functional morphology across several different baleen whale species. Kahane-Rapport et al. (2020) used drone measurements of tagged whales to analyze the biomechanics of how larger whales require longer times for filtering the water through their baleen when feeding. Gough et al. (2019) used size measurements from drones and swimming speeds from tags to determine that a whale’s “walking speed” is 2 meters per second – whether the largest of the whales, a blue whale, or the smallest of the baleen whales, an Antarctic minke whale. Size measurements and tag data were combined by Segre et al. (2019) to quantify the energetic costs of different sized whales when breaching.
Taken together, drones have revolutionized our ability to obtain morphological measurements of whales, greatly increasing our capacity to better understand how these animals function and perform in their environments. These advancements in marine science are particularly important as these methods provide greater opportunity to monitor the health of populations, especially as they face increased threats from anthropogenic stressors (such as vessel traffic, ocean noise, pollution, fishing entanglement, etc.) and climate change.
Drone-based photogrammetry is one of the main focuses of the GEMM Lab’s project on Gray whale Response to Ambient Noise Informed by Technology and Ecology (GRANITE). This summer we have been collecting drone videos to measure the body condition of gray whales feeding off the coast of Newport, Oregon (Fig. 2). As we try to understand the physiological stress response of gray whales to noise and other potential stressors, we have to account for the impacts of overall nutritional state of each individual whale’s physiology, which we infer from these body condition estimates.
Figure 2. Drones can help collect images of whales to obtain morphological measurements using photogrammetry and help us fill knowledge gaps for how these animals interact in their environment and to assess their current health. Bottom photo is an image collected by the GEMM Lab of a gray whale being measured in MorphoMetriX software to estimate its body condition.
References
Best, P. B., & Rüther, H. (1992). Aerial photogrammetry of southern right whales, Eubalaena australis. Journal of Zoology, 228(4), 595-614.
Breuer, T., Robbins, M. M., & Boesch, C. (2007). Using photogrammetry and color scoring to assess sexual dimorphism in wild western gorillas (Gorilla gorilla). American Journal of Physical Anthropology, 134(3), 369–382. https://doi.org/10.1002/ajpa.20678
Christiansen, F., Vivier, F., Charlton, C., Ward, R., Amerson, A., Burnell, S., & Bejder, L. (2018). Maternal body size and condition determine calf growth rates in southern right whales. Marine Ecology Progress Series, 592, 267–281.
Christiansen, F. (2020). A population comparison of right whale body condition reveals poor state of North Atlantic right whale, 1–43.
Christiansen, F., Dujon, A. M., Sprogis, K. R., Arnould, J. P. Y., & Bejder, L. (2016). Noninvasive unmanned aerial vehicle provides estimates of the energetic cost of reproduction in humpback whales. Ecosphere, 7(10), e01468–18.
Christiansen, F., Sironi, M., Moore, M. J., Di Martino, M., Ricciardi, M., Warick, H. A., … Uhart, M. M. (2019). Estimating body mass of free-living whales using aerial photogrammetry and 3D volumetrics. Methods in Ecology and Evolution, 10(12), 2034–2044.
Cubbage, J. C., & Calambokidis, J. (1987). Size-class segregation of bowhead whales discerned through aerial stereo-photogrammetry. Marine Mammal Science, 3(2), 179–185.
De Meyer, J., Verhelst, P., & Adriaens, D. (2020). Saving the European Eel: How Morphological Research Can Help in Effective Conservation Management. Integrative and Comparative Biology, 23, 347–349.
Gaudioso, V., Sanz-Ablanedo, E., Lomillos, J. M., Alonso, M. E., Javares-Morillo, L., & Rodr\’\iguez, P. (2014). “Photozoometer”: A new photogrammetric system for obtaining morphometric measurements of elusive animals, 1–10.
Gough, W. T., Segre, P. S., Bierlich, K. C., Cade, D. E., Potvin, J., Fish, F. E., … Goldbogen, J. A. (2019). Scaling of swimming performance in baleen whales. Journal of Experimental Biology, 222(20), jeb204172–11.
Groskreutz, M. J., Durban, J. W., Fearnbach, H., Barrett-Lennard, L. G., Towers, J. R., & Ford, J. K. B. (2019). Decadal changes in adult size of salmon-eating killer whales in the eastern North Pacific. Endangered Species Research, 40, 1
Kahane-Rapport, S. R., Savoca, M. S., Cade, D. E., Segre, P. S., Bierlich, K. C., Calambokidis, J., … Goldbogen, J. A. (2020). Lunge filter feeding biomechanics constrain rorqual foraging ecology across scale. Journal of Experimental Biology, 223(20), jeb224196–8.
Leedy, D. L. (1948). Aerial Photographs, Their Interpretation and Suggested Uses in Wildlife Management. The Journal of Wildlife Management, 12(2), 191.
Lemos, L. S., Burnett, J. D., Chandler, T. E., Sumich, J. L., and Torres, L. G. (2020). Intra- and inter-annual variation in gray whale body condition on a foraging ground. Ecosphere 11.
Leslie, M. S., Perkins-Taylor, C. M., Durban, J. W., Moore, M. J., Miller, C. A., Chanarat, P., … Apprill, A. (2020). Body size data collected non-invasively from drone images indicate a morphologically distinct Chilean blue whale (Balaenoptera musculus) taxon. Endangered Species Research, 43, 291–304.
Miles, D. B. (2020). Can Morphology Predict the Conservation Status of Iguanian Lizards? Integrative and Comparative Biology.
Miller, C. A., Best, P. B., Perryman, W. L., Baumgartner, M. F., & Moore, M. J. (2012). Body shape changes associated with reproductive status, nutritive condition and growth in right whales Eubalaena glacialis and E. australis. Marine Ecology Progress Series, 459, 135–156.
Negretti, P., Bianconi, G., Bartocci, S., Terramoccia, S., & Verna, M. (2008). Determination of live weight and body condition score in lactating Mediterranean buffalo by Visual Image Analysis. Livestock Science, 113(1), 1–7. https://doi.org/10.1016/j.livsci.2007.05.018
Ratnaswamy, M. J., & Winn, H. E. (1993). Photogrammetric Estimates of Allometry and Calf Production in Fin Whales, \emph{Balaenoptera physalus}. American Society of Mammalogists, 74, 323–330.
Perryman, W. L., & Lynn, M. S. (1993). Idendification of geographic forms of common dolphin(\emph{Delphinus Delphis}) from aerial photogrammetry. Marine Mammal Science, 9(2), 119–137.
Perryman, W. L., & Lynn, M. S. (2002). Evaluation of nutritive condition and reproductive status of migrating gray whales (\emph{Eschrichtius robustus}) based on analysisof photogrammetric data. Journal Cetacean Research and Management, 4(2), 155–164.
Perryman, W. L., & Westlake, R. L. (1998). A new geographic form of the spinner dolphin, stenella longirostris, detected with aerial photogrammetry. Marine Mammal Science, 14(1), 38–50.
Sasse, B. (2003). Job-Related Mortality of Wildlife Workers in the United States, 1937- 2000, 1015–1020.
Segre, P. S., Potvin, J., Cade, D. E., Calambokidis, J., Di Clemente, J., Fish, F. E., … & Goldbogen, J. A. (2020). Energetic and physical limitations on the breaching performance of large whales. Elife, 9, e51760.
Shirane, Y., Mori, F., Yamanaka, M., Nakanishi, M., Ishinazaka, T., Mano, T., … Shimozuru, M. (2020). Development of a noninvasive photograph-based method for the evaluation of body condition in free-ranging brown bears. PeerJ, 8, e9982. https://doi.org/10.7717/peerj.9982
Shrader, A. M., M, F. S., & Van Aarde, R. J. (2006). Digital photogrammetry and laser rangefinder techniques to measure African elephants, 1–7.
Stewart, J. D., Durban, J. W., Knowlton, A. R., Lynn, M. S., Fearnbach, H., Barbaro, J., … & Moore, M. J. (2021). Decreasing body lengths in North Atlantic right whales. Current Biology.
Walker, J. A., Alfaro, M. E., Noble, M. M., & Fulton, C. J. (2013). Body fineness ratio as a predictor of maximum prolonged-swimming speed in coral reef fishes. PloS one, 8(10), e75422.
About 10 months have passed since I started working on OPAL, a project that aims to identify the co-occurrence between whales and fishing effort in Oregon to reduce entanglement risk. During this period, you would be surprised to know how little ecology I have actually done and how much time has been devoted to data processing! I compiled several million GPS trackline positions, processed hundreds of marine mammal observations, wrote several thousand lines of R code, downloaded and extracted a couple Gb of environmental data… before finally reaching the modeling phase of the OPAL project. And with it, finally comes the time to look more closely at the ecology and behavior of my species of interest. While the previous steps of the project were pretty much devoid of ecological reasoning, the literature homework now comes in handy to guide my choices regarding habitat use models, such as selecting environmental predictors of whale occurrence, deciding on what seasons should be modeled, and choosing the spatio-temporal scale at which the data should be aggregated.
Whale diversity on the US west coast
The productive waters off the US west coast host a great diversity of cetaceans. Eight species of baleen whales are reported to occur there by NOAA fisheries: blue whales, Bryde’s whales, fin whales, gray whales, humpback whales, minke whales, North Pacific right whales and sei whales. Among them, no less than five are listed as Endangered under the Endangered Species Act. Whether they are only passing by or spending months feeding in the region, the timing and location where these animals are observed varies greatly by species and by population.
During the 113 hours of aerial survey effort and 264 hours of boat-based search conducted for the OPAL project, 563 groups of baleen whales have been observed to-date (up to mid-May 2021 to be exact… more data coming soon!). Among the observations where animals could be identified to the species level, humpback whales are preponderant, as they represent about half of the whale groups observed (n = 293). Blue (n = 41) and gray whales (n = 46) come next, the latter being observed in more nearshore waters. Finally, a few fin whale groups were observed (n = 28). The other baleen whale species reported by NOAA in the US west coast species list were very rarely or not observed at all during OPAL surveys.
The OPAL aerial surveys conducted in partnership with the United States Coast Guard (USCG) were specifically designed to study whales occurring on the continental shelf along the coast of Oregon. Hence, most of this survey effort is located in waters from 800 m to 30 m deep, which may explain the relatively low number of gray whales detected. Indeed, gray whales observed in Oregon may either be migrating along the coast to and from their breeding grounds in Baja California, or be part of the small Pacific Coast Feeding Group that forage in Oregon nearshore and shallow waters during the summer. This group of whales is one the main GEMM lab’s research focus, being at the core of no less than three ongoing research projects: AMBER, GRANITE, and TOPAZ.
So today, let’s turn our eyes to the sea horizon and talk about some other members of the baleen whale community: rorquals. Conveniently, the three species of baleen whales (gray whales aside) most commonly observed during OPAL surveys are all part of the rorqual family, a.k.a Balaenopteridae: humpback whales, blue whales and fin whales (Figure 1). They are morphologically characterized by the pleated throat grooves that allow them to engulf large quantities of food and water, for instance when lunge-feeding. Known cases of hybridization between these three species demonstrate their close relatedness (Jefferson et al., 2021). They all have worldwide distributions and display unequally understood migratory behaviors, seasonally traveling between warm tropical breeding grounds and temperate-polar feeding grounds. They occur in great numbers in productive waters such as the upwelling system of the California Current.
The three accomplices
Figure 1: Aerial view of three rorquals species: a humpback whale (left), a fin whale (center), and a blue whale (right). Photo credit: Leigh Torres and Craig Hayslip. Photos taken off the Oregon coast under NOAA/NMFS permit during USCG helicopter flights conducted as part of the OPAL project
Humpback whales (Megaptera novaeangliae) are easily differentiated from other rorquals because of their long pectoral fins (up to one third of their body length!), which inspired their scientific name, Megaptera, « big-winged » (Figure 1). Individuals observed in Oregon mostly belong to a mix of two Distinct Population Segments (DPS): the threatened Mexico and endangered Central American DPS. Although humpback whales from different DPS do not show any morphological differences, they are genetically distinct because they have been mating separately in distinct breeding grounds for generations and generations. This genetic differentiation has great implications in terms of conservation since the Central American DPS is recovering at a lesser rate than the Mexican and is therefore subject to different management measures (recovery plan, monitoring plan, designated critical habitats). Humpback whales migrate and feed off the US west coast, with a peak in abundance in the mid to late summer. Compared to other rorquals that are found in the open ocean, humpback whales are mostly observed on the continental shelf (Becker et al., 2019). They are considered to have a relatively generalist diet, as they feed on a mix of krill (Euphausiids) and fishes (e.g. anchovy, sardines) and are capable of switching their feeding behavior depending on relative prey availability (Fleming, Clark, Calambokidis, & Barlow, 2016; Fossette et al., 2017).
Blue whales (Balaenoptera musculus) are the largest animals ever known (max length 33 m, Jefferson et al., 2008), and sadly the most at risk of global extinction among our three species of interest (listed as « endangered » in the IUCN red list). They have a distinctive mottled blue and light gray skin, a slender body and a broad U-shaped head (or as some say « like a gothic arch », Figure 1). Blue whales tend to be open ocean animals, but they regroup seasonally to feed in highly productive nearshore areas such as the Southern California Bight (Becker et al. 2019, Abrahms et al. 2019). Blue whales migrating or feeding along the US west coast belong to the Eastern North Pacific stock and are subject to great research and conservation efforts. Contrary to their other rorqual counterparts, blue whales are quite picky eaters, as they exclusively feed on krill. This difference in diet leads to resource partitioning facilitating rorqual coexistence in the California Current (Fossette et al., 2017). These differences in feeding strategies have important implications for designing predictive models of habitat use.
Fin whales (Balaenoptera physalus) are nicknamed « greyhounds of the sea » due to their exceptional swim speed (max 46 km/h). They are a little smaller than blue whales (max length 27 m, Jefferson, Webber, & Pitman, 2008) but share a similar sleek and streamlined shape. Their coloration is their most distinctive feature: the left lower jaw being mostly dark while the right is white. V-shaped light-gray « chevrons » color their back, behind the head (Figure 1). The California/Oregon/Washington is one of the three stocks recognized in the North Pacific (NOAA Fisheries, 2018). Within this region, there is genetic evidence for a geographic separation north and south of Point Conception, CA (Archer et al., 2013). Like other rorquals, they are migratory, but their seasonal distribution is relatively less well understood as they appear to spend a lot of time in open oceans. For instance, a meta-analysis for the North Pacific found little evidence for fin whales using distinct calving areas (Mizroch, Rice, Zwiefelhofer, Waite, & Perryman, 2009). In the California Current System, satellite tracking has provided great insights into their space-use patterns. In the Southern California Bight, fin whales show year-round residency and seasonal shifts in habitat use as they move further offshore and north during the spring/summer (Scales et al., 2017). The Northern California Current offshore waters appeared to be used during the summer months by the whales tagged in the Southern California Bight. Yet, fin whales are observed year-round in Oregon (NOAA Fisheries, 2018).
Towards predictive models of rorqual distribution
Enough observations have now been collected as part of the OPAL project to be able to model the habitat use of some of these rorqual species. Based on 12 topographic (i.e., depth, slope, distance to canyons) and physical variables (temperature, chlorophyll-a, water column stratification, etc.), I have made my first attempt at predicting seasonal distribution patterns of humpback whales and blue whales in Oregon. These models will be improved in the coming months, with more data pouring in and refined parametrizations, but they already bring insights into the shared habitat use patterns of these species, as well as their specificities.
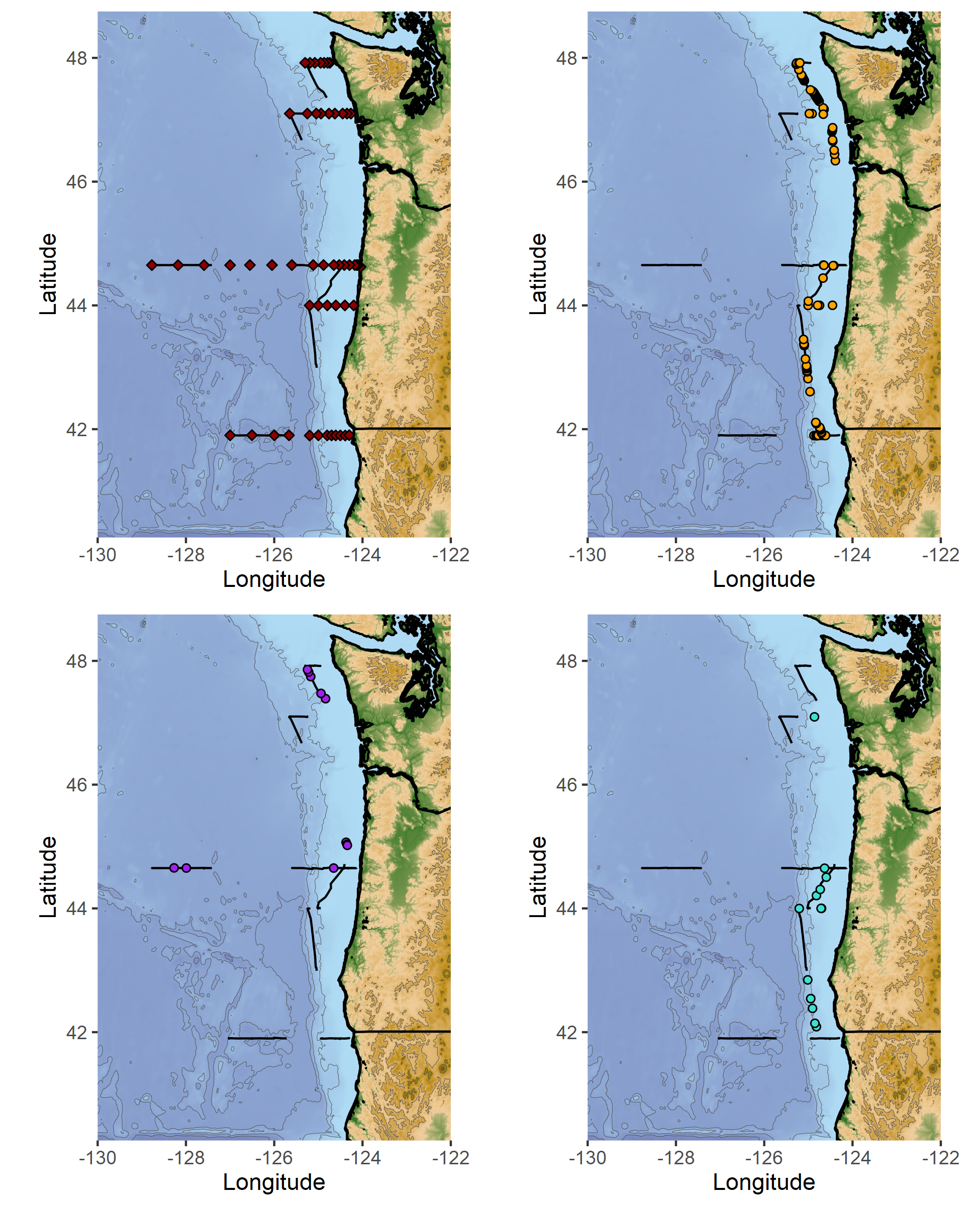
Across multiple cross-validations of the species-specific models, sea surface temperature, sea surface height and depth were recurrently selected among the most important variables influencing both humpback and blue whale distributions. Predicted densities of blue whales were relatively higher at less than 40 fathoms compared to humpback whales, although both species’ hotspots were located outside this newly implemented seasonal fishing limit (Figure 2). Higher densities were generally predicted off Newport and Port Orford, and north of North Bend.
Figure 2: Predicted densities of humpback and blue whales during the month of September 2018, 2019, and 2020 in Oregon waters (OPAL project). Core areas of use (predicted densities in the top 25%) are represented, with darker shades of blue and orange showing higher predicted densities. Dashed lines represent the tracklines followed by USCG monthly aerial surveys. The black line represents the 40 fathom isobath. Grey boxes overlayed on predictions delineate the areas of extrapolation where environmental conditions are non-analogous to the conditions in which the models were trained. Disclaimer: these model outputs are preliminary and should be interpreted with caution.
Once our rorqual models are finalized, we will work with our partners at the Oregon Department of Fisheries and Wildlife to overlay predicted whale hotspots with areas of high crab pot densities. This overlap analysis will help us understand the times and places where co-occurrence of suitable whale habitat and fishing activities put whales at risk of entanglement.
Becker, E. A., Forney, K. A., Redfern, J. V, Barlow, J., Jacox, M. G., Roberts, J. J., & Palacios, D. M. (2019). Predicting cetacean abundance and distribution in a changing climate. Diversity and Distributions, 25(4), 626–643. https://doi.org/10.1111/ddi.12867
Fleming, A. H., Clark, C. T., Calambokidis, J., & Barlow, J. (2016). Humpback whale diets respond to variance in ocean climate and ecosystem conditions in the California Current. Global Change Biology, 22, 1214–1224. https://doi.org/10.1111/gcb.13171
Fossette, S., Abrahms, B., Hazen, E. L., Bograd, S. J., Zilliacus, K. M., Calambokidis, J., … Croll, D. A. (2017). Resource partitioning facilitates coexistence in sympatric cetaceans in the California Current. Ecology and Evolution, 7, 9085–9097. https://doi.org/10.1002/ece3.3409
Jefferson, T. A., Palacios, D. M., Clambokidis, J., Baker, S. C., Hayslip, C. E., Jones, P. A., … Schulman-Janiger, A. (2021). Sightings and Satellite Tracking of a Blue / Fin Whale Hybrid in its Wintering and Summering Ranges in the Eastern North Pacific. Advances in Oceanography & Marine Biology, 2(4), 1–9. https://doi.org/10.33552/AOMB.2021.02.000545
Jefferson, T. A., Webber, M. A., & Pitman, R. L. (2008). Marine Mammals of the World. A comprehensive guide to their identification. Elsevier, London, UK.
Mizroch, S. A., Rice, D. W., Zwiefelhofer, D., Waite, J., & Perryman, W. L. (2009). Distribution and movements of fin whales in the North Pacific Ocean. Mammal Review, 39(3), 193–227. https://doi.org/10.1111/j.1365-2907.2009.00147.x
NOAA Fisheries. (2018). Fin whale stock assessment report ( Balaenoptera physalus physalus ): California / Oregon / Washington Stock.
Scales, K. L., Schorr, G. S., Hazen, E. L., Bograd, S. J., Miller, P. I., Andrews, R. D., … Falcone, E. A. (2017). Should I stay or should I go? Modelling year-round habitat suitability and drivers of residency for fin whales in the California Current. Diversity and Distributions, 23(10), 1204–1215. https://doi.org/10.1111/ddi.12611
Large whale conservation is challenged by our limited understanding of the impacts of natural and anthropogenic disturbances on the whale´s health and its population level consequences. To better mitigate human-wildlife conflicts, we need to improve our ability to predict multi-scale responses of whales to disturbances, describe and identify disease dynamics, and understand the reproductive biology of whales (Madliger, 2020; McCormick and Romero, 2017). Conservation physiology and conservation endocrinology can provide tools to illuminate the underlying physiological mechanisms whales use to cope with changing environments and different stressors, thus filling information gaps to guide management and conservation actions.
In brief, conservation physiology is a multidisciplinary field wherein a broad suite of tools and concepts are used to understand how organisms and ecosystems respond to both environmental and anthropogenic change and stressors (Madliger et al., 2020). Conservation endocrinology is a subdiscipline within conservation physiology, which relies on endocrine measurements (hormone quantifications). However, monitoring the physiology of free ranging animals in wild populations presents many technical challenges and it is particularly difficult when studying whales. Traditionally, conservation endocrinology relied on laboratory analyses of plasma samples (derived from blood). Yet implementing this techniques for monitoring the physiology of mysticetes (baleen whales) is currently impossible, as there are no feasible, non- (or minimally) invasive, methods to obtain a blood sample from living large whales (Hunt et al., 2013).
Therefore, we are interested in the development and further validation of alternative sample types from whales to obtain endocrine data. During my Ph.D. dissertation I worked to develop and ground truth the endocrine analyses of whale baleen as a novel sample type that can be used for retrospective assessments of the whale´s physiology. Baleen, the filter-feeding apparatus of the mysticete whales (Figure 1), consists of long fringed plates of stratified, keratinized tissue that grow continuously and slowly downward from the whale´s upper jaw (Hunt et al., 2014). Baleen plates are readily accessible at necropsy and routinely collected from carcasses of stranded whales.
Figure 1: Southern Right Whale skim feeding at the surface. Baleen plates hanging downwards are visible in the image. Photo: Alejandro Fernández Ajó / Instituto de Conservación de Ballenas de Argentina.
Like hair, nails, feathers, spines, or horns of other animals, baleen is a keratinized tissue that can store steroid and thyroid hormones in detectable and relevant concentrations to provide an integrated measure of hormonal plasma levels over the period that the structure was growing. Thus, baleen contains a progressive time-series that captures months and often years of an individual’s endocrine history with sufficient temporal resolution to determine seasonal endocrine patterns allowing to explore questions that have historically been difficult to address in large whales, including pregnancy and inter-calving interval, age of sexual maturation, timing and duration of seasonal reproductive cycles, adrenal physiology, and metabolic rate. Additionally, their robust and stable keratin matrix allows baleen samples to be stored for years to decades, enabling the analysis and comparison of endocrine patterns from past and modern populations. Therefore, keratinized sample matrices are valuable tools to investigate reproductive and stress physiology in whales and other vertebrates.
However, due to its novelty, the extraction and analysis of hormones from baleen and other keratinized tissues requires both biological and analytical validations to ensure the method fulfills the requirements for its intended use. Baleen hormone analyses has already passed several essential assay validations, including parallelism and accuracy of immunoassays (Hunt et al., 2017b), and numerous biological validations, such as the study of animals with known physiological status (i.e., pregnancy, and known stress events such as entanglement in fishing gear or presence of lesions) to assess the degree to which the endocrine data reflect the physiology of the individual (Fernández Ajó et al., 2020, 2018; Hunt et al., 2018, 2017a; Lysiak et al., 2018; Palme, 2019). Yet, other questions essential for technical validation remain unknown, including choice and volume of extraction solvent, the effect of solvent-to-sample ratio (solvent:sample) on extraction yield, and the amount of sample (e.g., mg) needed for analysis to obtain reliable hormonal data.
In our recent contribution,Optimizing hormone extraction protocols for whale baleen: Tackling questions of solvent:sample ratio and variation, we aimed to tackle two of these important questions: “1) what is the minimum sample mass of baleen powder required to reliably quantify hormone content of baleen samples analyzed using commercially available enzyme immune assays (EIAs); and 2) what is the optimal ratio of solvent volume to sample mass for steroids extracted from baleen, i.e., the ratio that yields the maximum amount of hormone with high accuracy and low variability between replicates.”
We performed the extraction with methanol and tested a variety of sample masses with the objective to provide methodological guidance regarding optimizing sample mass and solvent volume for steroid hormone extraction from powdered baleen. Our results suggest that the optimal sample mass for methanol extraction of steroid hormones from baleen samples is 20 mg, and that larger sample masses did not produce either better yield or less variation in the apparent hormone per g of baleen sample (Figure 2). In addition, when the extraction was performed keeping the volume of solvent proportional to the sample mass (namely, a solvent:sample ratio of 80:1), masses as small as 10 mg yielded reliable hormone measurement (Figure 2).
Figure 2. Extraction experiment results for variable (solid circles) or constant (open circles) solvent:sample ratio (mean progesterone concentration ± standard error of the mean). Extractions for 5, 10, 20, 50, and 75 mg of baleen powder were performed in triplicate for each sample mass. Letters indicate significant differences between groups after Tukey’s post hoc test. The right-side panel presents a plot for 100 mg of baleen powder extracted with 8 mL of solvent performed in triplicate; error bars are too small to show in the figure. Source: Fernández Ajó et al., 2021.
Our results indicate how baleen hormone analytic techniques can be more widely employed on small sample masses from rare specimens (i.e., less sample than is currently employed, which is typically 75 mg or 100 mg in most studies to date), such as from natural history museums and stranding archives. Thus, we demonstrate that greater use of this valuable technique to reconstruct the endocrine and physiological history of individual whales over time can be achieved with reduced sample size (so reduced damage to the sample). I hope these findings encourage researchers to apply these methods more broadly to analyze historical archives of baleen plates that can date back to the era of commercial whaling, and modern archives of baleen collected from stranded animals to help continue further developing techniques that can make headway in gaining conservation-relevant physiological knowledge of this particularly challenging taxon.
Bibliography:
Fernández Ajó, A., Hunt, K.E., Dillon, D., Uhart, M., Sironi, M., Rowntree, V., Loren Buck, C., 2021. Optimizing hormone extraction protocols for whale baleen: Tackling questions of solvent:sample ratio and variation. Gen. Comp. Endocrinol. 113828. https://doi.org/10.1016/j.ygcen.2021.113828
Fernández Ajó, A.A., Hunt, K.E., Giese, A.C., Sironi, M., Uhart, M., Rowntree, V.J., Marón, C.F., Dillon, D., DiMartino, M., Buck, C.L., 2020. Retrospective analysis of the lifetime endocrine response of southern right whale calves to gull wounding and harassment: A baleen hormone approach. Gen. Comp. Endocrinol. 296, 113536. https://doi.org/10.1016/j.ygcen.2020.113536
Fernández Ajó, A.A., Hunt, K.E., Uhart, M., Rowntree, V., Sironi, M., Marón, C.F., Di Martino, M., Buck, C.L., 2018. Lifetime glucocorticoid profiles in baleen of right whale calves: potential relationships to chronic stress of repeated wounding by Kelp Gulls. Conserv. Physiol. 6, 1–12. https://doi.org/10.1093/conphys/coy045
Hunt, K.E., Lysiak, N.S., Moore, M., Rolland, R.M., 2017a. Multi-year longitudinal profiles of cortisol and corticosterone recovered from baleen of North Atlantic right whales (Eubalaena glacialis). Gen. Comp. Endocrinol. 254, 50–59. https://doi.org/10.1016/j.ygcen.2017.09.009
Hunt, K.E., Lysiak, N.S.J., Matthews, C.J.D., Lowe, C., Fernández Ajó, A., Dillon, D., Willing, C., Heide-Jørgensen, M.P., Ferguson, S.H., Moore, M.J., Buck, C.L., 2018. Multi-year patterns in testosterone, cortisol and corticosterone in baleen from adult males of three whale species. Conserv. Physiol. 6, 1–16. https://doi.org/10.1093/conphys/coy049
Hunt, K.E., Moore, M.J., Rolland, R.M., Kellar, N.M., Hall, A.J., Kershaw, J., Raverty, S.A., Davis, C.E., Yeates, L.C., Fauquier, D.A., Rowles, T.K., Kraus, S.D., 2013. Overcoming the challenges of studying conservation physiology in large whales: a review of available methods. Conserv. Physiol. 1, cot006–cot006. https://doi.org/10.1093/conphys/cot006
Hunt, K.E., Stimmelmayr, R., George, C., Hanns, C., Suydam, R., Brower, H., Rolland, R.M., 2014. Baleen hormones: a novel tool for retrospective assessment of stress and reproduction in bowhead whales (Balaena mysticetus). Conserv. Physiol. 2, cou030–cou030. https://doi.org/10.1093/conphys/cou030
Lysiak, N.S.J., Trumble, S.J., Knowlton, A.R., Moore, M.J., 2018. Characterizing the Duration and Severity of Fishing Gear Entanglement on a North Atlantic Right Whale (Eubalaena glacialis) Using Stable Isotopes, Steroid and Thyroid Hormones in Baleen. Front. Mar. Sci. 5, 1–13. https://doi.org/10.3389/fmars.2018.00168
Madliger, C. L., Franklin, C. E., Love, O. P., & Cooke, S.J. (Ed.), 2020. Conservation Physiology: Applications for Wildlife Conservation and Management., 1st ed. Oxford University Press. https://doi.org/10.1093/oso/ 978019883610.001.0001
Palme, R., 2019. Non-invasive measurement of glucocorticoids: Advances and problems. Physiol. Behav. 199, 229–243. https://doi.org/10.1016/j.physbeh.2018.11.021
Hello from the RV Bell M. Shimada! We are currently sampling at an inshore station on the Heceta Head Line, which begins just south of Newport and heads out 45 nautical miles west into the Pacific Ocean. We’ll spend 10 days total at sea, which have so far been full of great weather, long days of observing, and lots of whales.
Dawn and Rachel in matching, many-layered outfits, 125 miles offshore on the flying bridge of the RV Bell M. Shimada.
Run by NOAA, this Northern California Current (NCC) cruise takes place three times per year. It is fabulously interdisciplinary, with teams concurrently conducting research on phytoplankton, zooplankton, seabirds, and more. The GEMM Lab will use the whale survey, krill, and oceanographic data to fuel species distribution models as part of Project OPAL. I’ll be working with this data for my PhD, and it’s great to be getting to know the region, study system, and sampling processes.
I’ve been to sea a number of times and always really enjoyed it, but this is my first time as part of a marine mammal survey. The type and timing of this work is so different from the many other types of oceanographic science that take place on a typical research cruise. While everyone else is scurrying around, deploying instruments and collecting samples at a “station” (a geographic waypoint in the ocean that is sampled repeatedly over time), we – the marine mammal team – are taking a break because we can only survey when the boat is moving. While everyone else is sleeping or relaxing during a long transit between stations, we’re hard at work up on the flying bridge of the ship, scanning the horizon for animals.
Top left: marine mammal survey effort (black lines), and oceanographic sampling stations (red diamonds). Top right: humpback whale sighting locations. Bottom left: fin whale sighting locations. Bottom right: pacific white-sided dolphin sighting locations.
During each “on effort” survey period, Dawn and I cover separate quadrants of ocean, each manning either the port or starboard side. We continuously scan the horizon for signs of whale blows or bodies, alternating between our eyes and binoculars. During long transits, we work in chunks – forty minutes on effort, and twenty minutes off effort. Staring at the sea all day is surprisingly tiring, and so our breaks often involve “going to the eye spa,” which entails pulling a neck gaiter or hat over your eyes and basking in the darkness.
Dawn has been joining these NCC cruises for the last four years, and her wealth of knowledge has been a great resource as I learn how to survey and identify marine mammals. Beyond learning the telltale signs of separate species, one of the biggest challenges has been learning how to read the sea better, to judge the difference between a frothy whitecap and a whale blow, or a distant dark wavelet and a dorsal fin. Other times, when conditions are amazing and it feels like we’re surrounded by whales, the trick is to try to predict the positions and trajectory of each whale so we don’t double-count them.
Over the last week, all our scanning has been amply rewarded. We’ve seen pods of dolphins play in our wake, and spotted Dall’s porpoises bounding alongside the ship. Here on the Heceta Line, we’ve seen a diversity of pinnipeds, including Northern fur seals, Stellar sea lions, and California sea lions. We’ve been surprised by several groups of fin whales, farther offshore than expected, and traveled alongside a pod of about 12 orcas for several minutes, which is exactly as magical as it sounds.
Killer whales traveling alongside the Bell M. Shimada, putting on a show for the NCC science team and ship crew. Photo by Dawn Barlow.
Notably, we’ve also seen dozens of humpbacks, including along what Dawn termed “the humpback highway” during our transit offshore of southern Oregon. One humpback put on a huge show just 200 meters from the ship, demonstrating fluke slapping behavior for several minutes. We wanted to be sure that everyone onboard could see the spectacle, so we radioed the news to the bridge, where the officers control the ship. They responded with my new favorite radio call ever: “Roger that, we are currently enamored.”
A group of humpbacks traveling along the humpback highway. Photo by Dawn Barlow.A humpback whale fluke slapping. Photo by Dawn Barlow.
Even with long days and tired eyes, we are still constantly enamored as well. It has been such a rewarding cruise so far, and it’s hard to think of returning back to “real life” next week. For now, we’re wishing you the same things we’re enjoying – great weather, unlimited coffee, and lots of whales!