Dynamic forecast models predict environmental conditions and blue whale distribution up to three weeks into the future, with applications for spatial management. Founded on a robust understanding of ecological links and lags, a recent study by Barlow & Torres presents new tools for proactive conservation.
The ocean is dynamic. Resources are patchy, and animals move in response to the shifting and fluid marine environment. Therefore, protected areas bounded by rigid lines may not always be the most effective way to conserve marine biodiversity. If the animals we wish to protect are not within protected area boundaries, then ocean users pay a price without the conservation benefit. Management that is adaptive to current conditions may more effectively match the dynamic nature of the species and places of concern, but this approach is only feasible if we have the relevant ecological knowledge to implement it.
The South Taranaki Bight region of New Zealand is home to a foraging ground for a unique population of blue whales that are genetically distinct and present year-round. The area also sustains New Zealand’s most industrial marine region, including active petroleum exploration and extraction, and vessel traffic between ports.
To minimize overlap between blue whale habitat and human use of the area, we develop and test forecasts of oceanographic conditions and blue whale habitat. These tools enable managers to make decisions with up to three weeks lead time in order to minimize potential overlap between blue whales and other ocean users.
Overlap between blue whale habitat and industry presence in the South Taranaki Bight region. A blue whale surfaces in front of a floating production storage and offloading (FPSO) vessel, servicing the oil rigs in the area. Photo by Dawn Barlow.
Predicting the future
Knowing where animals were yesterday may not create effective management boundaries for tomorrow. Like the weather, our expectation of when and where to find species may be based on long-term averages of previous patterns, real-time descriptions based on recent data, and forecasts that predict the future using current conditions. Forecasts allow us to plan ahead and make informed decisions needed to produce effective management strategies for dynamic systems.
Just as weather forecasts help us make decisions about whether to wear a raincoat or pack sunscreen before leaving the house, ecological forecasts can enable managers to anticipate environmental conditions and species distribution patterns in advance of industrial activity that may pose risk in certain scenarios.
In our recent study, we develop and test models that do just that: forecast where blue whales are most likely to be, allowing informed decision making with up to three weeks lead time.
Harnessing accessible data for an applicable tool
We use readily accessible data gathered by satellites and shore-based weather stations and made publicly available online. While our understanding of the ecosystem dynamics in the South Taranaki Bight is founded on years of collecting data at-sea and ecological analyses, using remotely gathered data for our forecasting tool is critical for making this approach operational, sustainable, and useful both now and into the future.
Measurements of conditions such as wind speed and ocean temperature anomaly are paired with known measurements of the lag times between wind input, upwelling, productivity, and blue whale foraging opportunities to produce forecasted environmental conditions.
Example environmental forecast maps, illustrating the predicted sea surface temperature and productivity in the South Taranaki Bight region, which can be forecasted by the models with up to three weeks lead time.
The forecasted environmental layers are then implemented in species distribution models to predict suitable blue whale habitat in the region, generating a blue whale forecast map. This map can be used to evaluate overlap between blue whale habitat and human uses, guiding management decisions regarding potential threats to the whales.
Example forecast of suitable blue whale habitat, with areas of higher probability of blue whale occurrence shown by the warmer colors and the area classified as “suitable habitat” denoted by the white boundaries. This habitat suitability map can be produced for any day in the past 10 years or for any day up to three weeks in the future.
Dynamic ecosystems, dynamic management
These forecasts of whale distribution can be effectively applied for dynamic spatial management because our models are founded on carefully measured links and lags between physical forcing (e.g., wind drives cold water upwelling) and biological responses (e.g., krill aggregations create feeding opportunities for blue whales). The models produce outputs that are dynamic and update as conditions change, matching the dynamic nature of the ecosystem.
A blue whale raises its majestic fluke on a deep foraging dive in the South Taranaki Bight. Photo by Leigh Torres.
Engagement with stakeholders—including managers, scientists, industry representatives, and environmental organizations—has been critical through the creation and implementation of this forecasting tool, which is currently in development as a user-friendly desktop application.
Our forecast tool provides managers with lead time for decision making and allows flexibility based on management objectives. Through trial, error, success, and feedback, these tools will continue to improve as new knowledge and feedback are received.
The people behind the science, from data collection to conservation application. Left: Dawn Barlow and Dr. Leigh Torres aboard a research vessel in New Zealand in 2017, collecting data on blue whale distribution patterns that contributed to the findings in this study. Right: Dr. Leigh Torres and Dawn Barlow at the Parliament buildings in Wellington, New Zealand, where they discussed research findings with politicians and managers, gathered feedback on barriers to implementation, and subsequently incorporated feedback into the development and implementation of the forecasting tools.
Reference: Barlow, D. R., & Torres, L. G. (2021). Planning ahead: Dynamic models forecast blue whale distribution with applications for spatial management. Journal of Applied Ecology, 00, 1–12. https://doi.org/10.1111/1365-2664.13992
The 2021 TOPAZ (Theodolite Overlooking Predators And Zooplankton) field season in Port Orford has come to a close. Its close also signals the end of my tenure as field project lead, after I took over from my predecessor Florence Sullivan (OSU/GEMM Lab MSc grad) in the summer of 2018. Allison Dawn, incoming GEMM Lab Master’s student, is my successor and I am excited to pass the torch to her and see what new directions she will take the project. In today’s post, I will not recap the field season as I often do at the end of August. However, I strongly encourage you to read the blog posts written by the JASPER (Journey for Aspiring Scientists Pursuing Ecological Research) interns that made up Team “Heck Yeah”, Nadia Leal, Damian Amerman-Smith, and Jasen White, as they did an excellent job summarizing what we saw and experienced over the last six weeks. Instead, I want to take this opportunity to highlight a few people in Port Orford (and their most memorable gray whale encounters) who created a home away from home for me in Port Orford and played a large part in creating rich and meaningful experiences during my time as field project lead.
Up first is Tom Calvanese, the OSU Port Orford Field Station manager. The field station can be an extremely busy place, especially during the summer when ideal weather conditions allow many marine scientists to conduct their research. There can be a lot of comings and goings at the field station, with swift turnarounds between groups and individuals from different departments and projects; some staying just one night, while others (such as the TOPAZ field teams) stay for several weeks. Leigh and I like to call Tom “the man behind the machine” because he manages to keep this busy field station running smoothly. From the get go, Tom has been a solid rock for me in Port Orford and he has never hesitated to give me the time and attention I needed, be it because I was seeking him out for advice about how to handle a personnel issue, a lesson in how to tie strong knots, or just a friendly conversation at the end of a long field day. I know that I have found a life-long friend and colleague in Tom through this project and for this I am very grateful.
One of Tom’s most iconic gray whale encounters happened when he was kayak fishing with a few friends in Tichenor Cove (coincidentally one of the two TOPAZ study sites). The individual kayakers were scattered throughout the cove, all in search of a good spot to hook some rockfish or lingcod. The group had not been out on the water for very long, which likely plays a large part in the shock and surprise that comes next, when Tom suddenly heard the blow of whale. He looked up from his fishing in the direction of the blow, only to see that a gray whale was surfacing right underneath one of his kayak fishing friends. Said friend could do nothing as he sat paralyzed in his kayak which slowly slid off the back of the gray whale as it dove once again. Neither whale nor human was harmed in this encounter, as the whale went back to foraging in the area, and the human (after several minutes of incredulity) went back to fishing. Every year, Tom has warned me of this location where this interaction happened (an uncharacteristically deep spot in Tichenor Cove compared to the rest of the area), though his warning is always accompanied with a twinkle in his eye.
An image captured by 2018’s Team “Whale Storm” aboard the kayak while sampling in Tichenor Cove, Port Orford. Source: GEMM Lab.
Dave Lacey. Source: L Hildebrand.
Dave Lacey owns South Coast Tours (SCT), a tour operating business that offers boat, kayak, and snorkeling tours, as well as surf lessons. Dave has been one of the most generous individuals to the TOPAZ/JASPER projects, never hesitating to loan us wetsuits and/or kayaks and allowing us to use his office and storage areas every day. He has also delivered excellent kayak paddle & safety instruction to the field teams over the last two years. Dave has truly become a vital partner during the Port Orford field seasons. It has been such a pleasure to be able to learn from and work with him, as well as see his business grow each year. Even though I will not be leading the project in Port Orford anymore, I am excited to continue my working relationship with Dave through obtaining important photo identification and sighting data of gray whales in the area when the GEMM Lab team is not there.
Although SCT is not even 10 years old (though it will be next year in 2022!), Dave has had so many gray whale encounters that he said it was really hard for him to pick just one. However, he ultimately picked the first time that he smelled a gray whale’s breath. It happened during a kayak tour when the group rounded the corner from Tichenor to Nellie’s Cove and a whale suddenly surfaced right in front of everyone, hitting them with the misty cloud of its blow. Up until this moment, Dave had both seen and heard hundreds of whale blows, but had never smelled one. He says, “to hear and see [the blow] is pretty normal but to get the third sense [of smell] is really phenomenal.”. Upon asking what he thinks of the smell, Dave replied that he does not think it is as gross as some people may think and during tours on his boat, the Black Pearl, he now actually tries to (safely) maneuver the boat downwind of the blow so that his clients can get a whiff as well.
The misty cloud emitted by whales when they come to the surface to breathe is referred to as the “blow”. Source: GEMM Lab.
Mike Baran. Source: L Hildebrand.
Mike Baran is a co-owner of Port Orford Sustainable Seafood (POSS) and he also occasionally guides kayak and snorkel tours for SCT. POSS is a community supported fishery that delivers wild, line-caught seafood direct from Port Orford to communities throughout western Oregon. I developed a great friendship with Mike through seeing him on the water a lot as a kayak guide for SCT in my first summer leading the TOPAZ/JASPER projects (2018), as well as seeing him at the field station on most days since POSS’ office and fish-processing facility are located there as well. If you are a keen follower of the GEMM Lab blog, you will know by now that the field season in Port Orford is short, yet very intense and taxing. Therefore, uplifting and sometimes goofy interactions with someone can really turn an upsetting day (potentially due to kayak gear loss or simply exhaustion) into a better one. Mike provided me with a lot of uplifting and goofy interactions and always helped put a smile on my face.
As a SCT kayak guide, Mike has also had many gray whale encounters, however none are as memorable as the one he had on August 2nd, 2019. Mike describes it as a typical Port Orford day: “windy with lots of whale activity all morning”, though all of the activity had been at a distance (the whale blows were far away). Yet, on the paddle back through Tichenor Cove along the backside of the port jetty, Mike and his tour glimpsed a whale that was headstanding along the jetty rocks. The paddlers slowed down and kept their distance, watching as the gray whale foraged, diving down for 3-4 minutes at a time before resurfacing in almost the same location as it had surfaced in before. Suddenly, the whale surfaced right in the middle of the kayak group, with Mike to its left, a mere meter or so away, and the rest of the group to its right. Despite the fact that the sudden appearance of the whale scared the living daylights out of Mike, he was able to take a picture of the surfacing, which features one of the tour clients in the background with her hands lifted up to her face in total shock. So, thankfully for us the moment is not just eternalized in Mike’s memory but also in photographic form.
The photo of the gray whale that surfaced right next to Mike’s kayak, which also captured the shock & surprise of one of the tour clients in the background. Source: South Coast Tours.
Tara Ramsey. Source: L Hildebrand.
Last but certainly not least is Tara Ramsey, the coordinator of the Redfish Rocks Community Team since the summer of 2020. Despite arriving to Port Orford and her job in the middle of a pandemic, Tara has developed a lot of exciting new outreach and education material for the Redfish Rocks Marine Reserve, including an excellent walking tour of Port Orford (if you are ever there, I cannot recommend it highly enough – it starts at the Visitor Center!). While I have not known Tara as long as the other individuals featured in this blog, she has become a really great friend of mine, teaching me a lot about the reserve and Port Orford in general, including the best spot on Battle Rock beach for a small nighttime bonfire.
Tara’s most memorable encounter with a gray whale is in fact her only encounter with a gray whale to date, and it happened just a few weeks ago when she was doing an Instagram livestream of the Redfish Rocks Marine Reserve aboard SCT’s Black Pearl. The purpose of the livestream was to bring the public into the reserve without having to leave the comfort and current safety of their homes. Tara describes the conditions in the reserve as “quite eerie” that day as there was a combination of smoke, fog, and no wind in the air. These conditions resulted in some pretty poor visibility, but gave the reserve an almost mystical appearance. Tara was actually mid-sentence on the livestream, talking about how special this moment was for her because it was her first time being in the reserve, when a whale surfaced a few meters from the boat. While the encounter was brief (the whale only surfaced 3 or 4 times before disappearing into the fog), Tara says the vision will be etched in her memory forever as Redfish Rocks is “a circle of islands, kind of like an amphitheater and it was amazing to see the whale just in the middle of it all.”
An aerial view of Redfish Rocks Marine Reserve. Source: FishTracker.
I will miss being the field project lead of the TOPAZ and JASPER projects. I will miss kayaking every other day and spying on gray whales from the cliff site. I will miss having the opportunity to work closely with and train a new crop of aspiring marine scientists. I will miss my daily interactions with Tom, Dave, Mike, Tara, and many more individuals, when I do not go to Port Orford for six weeks next summer. I will cherish all the memories I have amassed over my last four summers in Port Orford for a very long time. Most of all, I will always be grateful to the gray whales that brought me back every summer and who (in a way) made all those memories happen.
PI Leigh Torres and Lisa at the end of the 2021 TOPAZ field season in Port Orford after the annual community presentation with Battle Rock Beach, Humbug Mountain, and Redfish Rocks Marine Reserve in the background. Source: L Torres.
By Jasen C. White, GEMM Lab summer intern, OSU senior, Department of Fisheries, Wildlife, and Conservation Sciences
Field work is predictably unpredictable. Even with years of experience and exhaustive planning, nature always manages to throw a few curveballs, and this gray whale foraging ecology field season is no exception. We are currently in our sixth week of data collection here in Port Orford, and we have been battling the weather, our equipment, and a notable lack of whales and their zooplankton prey. Throughout all of these setbacks, Team “Heck Yeah” has lived up to its mantra as we have approached each day ready to hit the ground running. When faced with any of our myriad of problems, we have managed to work collaboratively to assess our options and develop solutions to keep the project on track.
For those of you that are unfamiliar with Port Orford, it is windy here, and when it is not, it can be foggy. Both of these weather patterns have the potential to make unsafe paddling conditions for our kayak sampling team. This summer we have frequently delayed or altered our field work routines to accommodate these weather patterns. Occasionally, we had to call off kayaking altogether as the winds and swell precluded us from maintaining our boat “on station” at the predetermined GPS coordinates during our samples, only for the winds to die down once we had returned to shore and completed the daily gear maintenance. Despite weather challenges, we have made the most of our data collection opportunities over these past six weeks, and we have only been forced to give up four total days of data collection. Flexibility to take advantage of the good weather windows when they arrive is the key!
Equipment issues can be even more unpredictable than the weather. The first major stumbling block for our equipment was a punctured membrane in the dissolved oxygen probe that we lower into the water at each of our twelve sample locations. This puncture was likely the result of a stray urchin’s spine that was in the wrong place at the wrong time. Soon after noticing the problem, we quickly rallied to refurbish the membrane, recalibrate the sensor, and design a protective housing using some plumbing parts from the local hardware store to prevent any future damage to the membrane (Figures 1a-d). Within 6 days, we were back up and running with the dissolved oxygen sensor.
Figure 1. a) Punctured dissolved oxygen sensor membrane; b) plans for constructing a protective housing for the sensor; c) the new protective housing for the dissolved oxygen sensor (yellow) is attached to the sensor array; d) intern Jasen White measuring seawater for the dissolved oxygen sensor calibration after replacing the punctured membrane. Source: A. Dawn
The next major equipment issue involved a GoPro camera whose mounting hardware snapped while being retrieved at a sample site. This event was captured on the camera itself (see below). Fortunately, thanks to our collaborators at the Oregon Institute of Marine Biology, we were soon able to recover the lost GoPro camera, and in the meantime, we relied on our spare to continue sampling.
Figure 2. The steel cable of the downrigger used to deploy and retrieve our sensor array had worn down until only two strands remained intact. Source: J. White.
The most recent equipment problem was a fraying cable (Figure 2) on our downrigger. We use the downrigger as a winch to lower and raise our sensor array and zooplankton nets into the water to obtain our samples. Fortunately, keen eyes on our team noticed the fray before it fully separated while the sensor array was in the water which could have resulted in losing our gear. We were quickly able to find the necessary repair part locally and get back on the water to finish out our sample regime within an hour of noticing the problem.
Finally, as Damian mentioned in his post last week, this season seemed to start much slower than the previous field seasons. In the early weeks, many of our zooplankton sampling nets repeatedly came up almost empty. There was often nothing but murky water to see in the GoPro videos that accompany the zooplankton samples. Likely due to the lack of prey, we have only managed to spot a couple of transitory whales that rarely entered our study area. Those few whales that we did observe were difficult to track as the relatively high winds and waves quickly dissipated the tell-tale blows and camouflaged their briefly exposed backs and flukes.
Our determination and perseverance have recently started to pay off, however, as the prey abundance in at least some of our sample sites has begun to increase. This increase in prey has also corresponded to a slight increase in whale sightings. One whale even spent nearly 30 minutes around the sampling station that consistently yields the most prey, likely indicating foraging behavior. These modest increases in zooplankton prey and whale sightings provide more evidence in support of the hypothesis Damian mentioned last week that reduced whale abundance in the area is likely the result of low prey abundance.
Figure 3. Example of a previously unidentified mysid that dominates several of our zooplankton samples. Due to the unique fat and flat telson (the “tail”) portion, we have been affectionately calling these “beavertail” mysids. Source: J. White.
As the zooplankton abundance finally started to increase, we noticed an interesting shift in the kinds of prey that we are capturing compared to previous seasons. Donovan Burns, an intern from the 2019 field season, noted in his blog post that the two most common types of zooplankton they found in their samples were the mysid species Holmesimysis sculpta and members of the genus Neomysis. While Neomysis mysid shrimp are continuing to make up a large proportion of our prey samples this year, we have noticed that many of our samples are dominated by a different type of mysid shrimp (Figure 3) which, in previous years, was a very rare capture. After searching through several mysid identification guides, this unknown mysid appears to be a member of the genus Lucifer, identified based on the presence of some distinctive characteristics that are unique to this genus (Omori 1992).
This observation is interesting because historically, Lucifer mysid shrimp are typically found in warmer tropical and subtropical waters and were rarely reported in the eastern North Pacific Ocean before the year 1992 (Omori 1992). Additionally, a key to common coastal mysid shrimp of Oregon, Washington, and British Columbia does not include members of the Lucifer genus, nor does it include any examples of mysids that resemble these new individuals showing up in our zooplankton nets (Daly and Holmquist 1986). If our initial identification of this mysid species is correct, then the sudden rise in the abundance of a typically warm water mysid species in Port Orford may indicate some fascinating shifts in oceanographic conditions that could lend some insight into why our prey and subsequent whale observations are so different this year than in years past.
Figure 4. View from the cliff site where we track gray whales using a theodolite. Source: A. Dawn.
As the 2021 field season draws to a close in Port Orford, I cannot help but reflect on what a wonderful opportunity we have been given through this summer internship program. I have loved the short time that I have spent living in this small but lively community for these past five weeks. Most days we could either be found kayaking around the nearshore to sample for the tiny creatures that our local gray whales call dinner, or we were on a cliff, gazing at the tirelessly beautiful, rugged coastline (Figure 4), hoping to glimpse the blow of a foraging whale so that we could track its course with our theodolite. Though the work can be physically exhausting during long and windy kayaking trips, mentally taxing when processing the data for each of the new samples after a full day of fieldwork, or incredibly frustrating with equipment failures, weather delays and shy whales, it is also tremendously satisfying to know that I contributed in a small but meaningful way to the mission of the GEMM Lab. I cannot imagine a better way to obtain the experience that my fellow interns and I have gained from this work, and I know that it will serve each of us well in our future ambitions.
References
Daly, K. L., and C. Holmquist. 1986. A key to the Mysidacea of the Pacific Northwest. Canadian Journal of Zoology 64:1201–1210.
Omori, M. 1992. Occurrence of Two Species of Lucifer (Dendrobranchiata: Sergestoidea: Luciferidae) off the Pacific Coast of America. Journal of Crustacean Biology 12:104–110.
By Damian Amerman-Smith, Pacific High School senior, GEMM Lab summer intern
Left to right: Damian, Nadia, Jasen. The group scans the ocean looking for whales, while Damian puts on sunscreen. Source: A. Dawn.
Growing up in Port Orford, a short ten-minute walk from the Pacific Ocean, has certainly shaped my life a lot. It has given me a great regard for the ocean, the diversity of life within it, and how life seems to bypass human derived borders in order to go wherever it can. I often marvel at all the beautiful, intricate ecosystems that are able to exist inside of our planet’s vast oceanic expanses. Along with my love of the ocean has come a great regard for marine mammals and the novelties of these animals that allow them to live entirely in the ocean despite not having gills. Every new discovery of these beautiful ocean creatures brings me such simple and pure joy, such as my very recent discovery that baleen whales have two blow holes. These blow holes look so peculiar on the top of their bodies, like a short upside-down nose.
My interest in the ocean and its inhabitants was a large part of what made me so enthused to take a part in the gray whale foraging ecology (GWFE) project in Port Orford this summer. When Elizabeth Kelly, my friend and a previous intern for the GWFE project mentioned her experiences from the previous summer, I was very happy when she put me in contact with Lisa Hildebrand and Leigh Torres so that I could apply to be an intern. Since then, I have been very ecstatically awaiting the beginning of the project and could hardly believe it when it finally began, and I was able to meet my fellow team members: Lisa Hildebrand, the PhD student who has been leading the GWFE project for the last four years; Allison Dawn, a Master’s student who is going to take over the project in Lisa’s stead; Nadia Leal, an OSU undergrad hoping to further pursue the field of marine biology; and Jasen White, an OSU undergrad whose time in the Navy has made him a very steeling presence while out on the water.
The three weeks that we have spent together learning the procedures that make up the project have been well spent, teaching all of us a lot of new things, such as what a theodolite is, how to operate a dissolved oxygen sensor, and (for me) how to use Excel. The first two weeks were largely spent just learning about how we collect data and improving our field skills, but as we have become more comfortable with our skills, we have also begun looking beyond the procedures, towards the data itself and what it can mean. Primarily, we started to notice the distinct lack of gray whales and almost complete lack of zooplankton prey for any gray whales in the area to eat.
A calm & beautiful, yet whale-less, view from the cliff site. Source: L. Hildebrand.
As we pass the halfway point in the project, we have only witnessed two whales inside our study area. While in the beginning it was not surprising that there were no whales, it has started to become concerning to me. We have a strong working hypothesis about why there have not been many whale sightings in our monitored sites of Mill Rocks and Tichenor’s Cove: there is not nearly enough zooplankton prey to attract them. Monday, August 9th is a good example to support this hypothesis. On that day, when we pulled up our sample net at Tichenor Cove station #1, we collected fifty-three individual Neomysis mysid shrimp, which are a tasty treat for gray whales. However, all the other prey samples from the remaining eleven kayak sampling stations had perhaps a maximum of five assorted zooplankton each, which is certainly not enough to attract the attention of such a large predator as Eschrichtius robustus (a gray whale). Unfortunately, we have yet to see much change in zooplankton prey availability in our sampling nets over the season so far, but we are hopeful that swarms of zooplankton in the area will resurge and the gray whales will begin using the area around the port as their August feeding grounds.
Our hopes aside, it is intriguing to think about why there has been so few zooplankton at our sampling sites. A main factor is likely the decrease of Port Orford’s kelp forests over the past few years. Kelp is very important to zooplankton, particularly mysids, as it allows them to seek shelter from predators. Declines in kelp forests have been documented all along the southern Oregon coast, and are believed to be fueled by many factors (ORKA, 2021). A combination of warming waters with decreasing amount of nutrients available to the kelp (Richardson 2008), and the increasing abundances of purple sea urchins that eat the kelp has vastly impacted the amount of kelp in the area. The decline in local kelp forests may be the reason that we are seeing fewer mysid swarms than in previous years. This reduced kelp and mysid availability could, in turn, be making Port Orford waters an unappetizing area for hungry whales to visit this year. While this trophic cascade is still just an educated hypothesis, it is important for us and others to keep watch on the situation, and to see how it changes. There are organizations such as the Oregon Kelp Alliance (ORKA) that are working hard to study why the kelp populations are hurting and how we can help. We will power through the season with the hopes that the gray whales will come. It is still very possible that the zooplankton will resurge and the whales will return with plenty to feed on.
References
Richardson, Anthony J. 2008. In hot water: zooplankton and climate change, ICES Journal of Marine Science, Volume 65, Issue 3, Pages 279–295, https://doi.org/10.1093/icesjms/fsn028
By Nadia Leal, GEMM Lab summer intern, OSU senior, Department of Fisheries, Wildlife, and Conservation Sciences
The OSU GEMM Lab gray whale foraging ecology project in Port Orford is in its seventh year of research. I have the honor to serve as a field assistant for the project as part of Team “Heck Yeah” for the summer 2021 field season. In doing so, I have been presented with the opportunity to take part in its enduring legacy. It is a legacy characterized by novel discovery, distinguished leadership, and endless adventure. These particular aspects motivated me to pursue this internship. Further, the desire to seek out gray whales (Eschrichtius robustus) — a species epitomizing the ability to exhibit resilience in the face of adversity after having experienced two unusual mortality events (UME) in the past two decades and having recovered from historically low population abundances due to whaling — sparked immeasurable excitement.
Figure 1. Nadia operating the theodolite to calculate the location of a gray whale. Source: A. Dawn.
The skills we are acquiring during this field season are essential to master so that I can pursue my aspirations of becoming a marine conservation biologist. For example, we have learned how to operate a theodolite, which is a surveying tool used regularly in marine mammal research to accurately calculate the location of cetaceans and track their movements (Figure 1). We are also learning how to operate a number of other research equipment, to navigate a tandem kayak using a GPS, to process various forms of data, and to identify gray whales! I have especially enjoyed collecting prey samples and navigating our tandem kayak, as kayaking is a summer tradition for my family and the opportunity to kayak in this context is certainly the high point of this internship. The kayak is named “Robustus” after the scientific name of the gray whale: Eschrichtius robustus! (Figure 2).
Figure 2. Nadia navigating Robustus, the research kayak.
The Port Orford project aims to determine how gray whale foraging is affected by prey quantity and quality. In fact, gray whales exhibit specificity in their selection of prey on the basis of caloric content (Hildebrand 2020). I am particularly interested in the underlying implications these findings imply: the notion of conscious reasoning and decision-making by individual whales as they seek the most suitable prey for its dietary needs among other options to maximize its survivability. Are gray whales in possession of an awareness that allows them to exhibit intentional preference? Can the behavior be attributed to instinct and/or learned behavior, or to cognition comparable to human preference? These and similar questions are my motivation for studying the realm of marine mammal biology. These questions concern intelligence and evolution, which can be effectively investigated through an analysis of cetacean brain structure, as it likely has compelling relationships to their extensive behavioral abilities (Hof and Van Der Gucht 2007).
For instance, the brain of the gray whale has expanded and developed extensively over evolutionary time in response to distinct selection pressures. Evidence affirms that the behavioral challenges associated with foraging exert strong selection pressures on the evolution of their brain size and structure (Muller and Montgomery 2019)! Selection pressures associated with social cognition are also believed to have contributed to such growth (Connor et al. 1998; Marino 2002; Shultz and Dunbar 2010 ). Further, their neural organization has increased in complexity, leading to greater function and usage of the cortical portion of the brain, which is the portion responsible for higher level activity (Oelschläger and Oelschläger 2002).
Figure 3. Structure of humpback whale brain representative of baleen species used to infer about gray whales (Hof and Van Der Gucht 2007).
Though research about baleen whale brain morphology is not as pervasive as that of toothed whales (due to increased susceptibility of toothed whales to captivity given the feasibility of their capture and subsequent analysis in lab/controlled setting), studies have indicated that the brain of baleen whales share similarities to those of humans (Wade et. al 2012). In particular, similarities exist in the frontal lobe of the brain, which is responsible for the complex activities of self-awareness, reasoning, and behavior, as well as for problem-solving and motivation (Hof and Van Der Gucht 2007) (Figure 3). These findings indicate that baleen whales, including the gray whale, have the capability to exhibit intentional preference and take part in conscious decision-making in the recognition of different prey species. The mechanisms responsible for how gray whales may discern prey likely involve a number of the sensory systems, differing in respect to spatial scale (Torres 2017). Thus, gray whales likely rely on various sensory methods, such as vision, sound perception/reception, chemoreception, or an oceanographic stimulus, at differing scales to locate and discern prey. The sensory method employed is dependent on their distance from prey.
Though we cannot yet confirm whether and/or how gray whales are capable of distinguishing between prey species, what is certain, is that the gray whale is intelligent and quite similar to us. Moreover, they are representative of strength and endurance, providing lessons we can learn from and qualities we can embody. Despite the threats to the species from fishing gear entanglement, ship collisions, climate change, oil industry developments, and being historically hunted, they have remarkably persisted. Thus, we must ensure the existence of the gray whale so they too may thrive for the rest of time, with healthy lives and habitat that is rightfully theirs.
P.S. I would like to thank the GEMM Lab, Oregon State University, Shalynn Pack, Port Orford Sustainable Seafood, Port Orford Co-op, South Coast Tours, Nicki’s Knick Knacks, Leigh Torres, Lisa Hildebrand, Allison Dawn, Clara Bird, Tom Calvanese, Maddie English, Jasen White, and Damian Amerman-Smith for making the internship as special and memorable as it is/was.
References
Connor, R. C., Mann, J., Tyack, P. L., and Whitehead, H. (1998). Social evolution in toothed whales. Trends in Ecology and Evolution, 13(6): 228– 232. doi: https://doi.org/10.1016/S0169‐5347(98)01326‐3
Hildebrand, L. (2020). Tonight’s specials include mysids, amphipods, and more: an examination of the zooplankton prey of Oregon gray whales and its impact on foraging choices and prey selection. Master’s thesis, Oregon State University.
Hof, P.R., and Van Der Gucht, E. (2007). Structure of the cerebral cortex of the humpback whale, Megaptera novaengliae(Cetacea, Mysticeti, Balaenopteridae). The Anatomical Record 290:1-31 doi: 10.1002/ar.a.20407
Marino, L. (2002). Convergence of complex cognitive abilities in cetaceans and primates. Brain, Behavior, and Evolution59: 21–32. doi: https://doi. org/10.1159/000063731
Oelschläger, H.A., and Oelschläger, J.S. (2002). Brains. In: Perrin WF, Wu¨ rsig B, Thewissen JGM, editors. Encyclopedia of marine mammals. San Diego, CA: Academic Press. p 133–158.
Shultz, S., & Dunbar, R. (2010). Encephalization is not a universal macroevolutionary phenomenon in mammals but is associated with sociality. Proceedings of the National Academy of Sciences of the United States of America 107(50): 21582–21586. doi: https://doi.org/10.1073/ pnas.1005246107
Torres, L.G. (2017). A sense of scale: foraging cetaceans’ use of scale-dependent multimodal sensory systems. Marine Mammal Science 33: 1170-1193. doi: 10.1111/mms.12426
Wade, P.R., Reeves, R.R., and Mesnick, S.L. (2012). Social and behavioral factors in cetacean responses to overexploitation: are odontocetes less “resilient” than mysticetes?. The Journal of Marine Biology 2012: 1-15. doi:10.1155/2012/567276
Predators have high energetic requirements that must be met to ensure reproductive success and population viability. For baleen whales, this task is particularly challenging since their foraging seasons are typically limited to short temporal windows during summer months when they migrate to productive high latitude environments. Foraging success is a balancing act whereby baleen whales must maximize the amount of energy they intake, while minimizing the amount of energy they expend to obtain food. Maximization of energy intake can be achieved by targeting the most beneficial prey. How beneficial a particular prey type (or prey patch) is can depend on a number of factors such as abundance, density, quality, size, and availability. Determining why baleen whales target particular prey types or patches is an important factor to enhance our understanding of their ecology and can ultimately aid in informing their management and conservation.
The GEMM Lab has several research projects in Newport and Port Orford, Oregon, on the Pacific Coast Feeding Group (PCFG), which is a sub-group of gray whales from the Eastern North Pacific (ENP) population. While ENP gray whales feed in the Bering, Chukchi, and Beaufort Seas (Arctic) in the summer months, the PCFG utilizes the range from northern California, USA to northern British Columbia, Canada. Our work to date has revealed a number of new findings about the PCFG including that they successfully gain weight during the summer on Oregon foraging grounds (Soledade Lemos et al. 2020). Furthermore, females that consistently use the PCFG range as their foraging grounds have successfully reproduced and given birth to calves (Calambokidis & Perez 2017). Yet, the abundance of the PCFG (~250 individuals; Calambokidis et al. 2017) is two orders of magnitude smaller than the ENP population (~20,000; Stewart & Weller 2021). So, why do more gray whales not use the PCFG range as their foraging grounds when it provides a shorter migration while also allowing whales to meet their high energetic requirements and ensure reproductive success? There are several hypotheses regarding this ecological mystery including that prey abundance, density, quality, and/or availability are higher in the Arctic than in the PCFG range, thus justifying the much larger number of gray whales that migrate further north for the summer feeding season.
Figure 1. Locations of prey samples collected with a light trap (open circles) or opportunistic collections of surface swarms of crab larvae (black triangles) in Newport, along the Oregon coast in the Pacific Northwest coast of the United States.
Our recent paper in Frontiers in Marine Science addressed the hypothesis that prey quality in the Arctic is higher than that of PCFG prey. To test this hypothesis, we first determined the quality (energetic value) of nearshore Oregon zooplankton species that PCFG gray whales are assumed to feed on (based on observations of fine-scale spatial and temporal overlap of foraging gray whales and sampled zooplankton). We obtained prey samples from nearshore reefs along the Oregon coast (Figure 1) as part of the GRANITE project using a light trap, which is a modified water jug with a weight and two floats attached to it, allowing the trap to sit approximately 1 meter above the seafloor. The trap contained a light which attracted zooplankton and effectively captured epibenthic prey of gray whales. Traps were left to soak overnight in locations where gray whales had been observed feeding extensively and collected the following morning. After identifying each specimen to species level and sorting them into reproductive stages, we used a bomb calorimeter to determine the caloric content of each species by month, year, and reproductive stage. We then compared these values to the literature-derived caloric value of the predominant benthic amphipod species that ENP gray whales feed on in the Arctic. These comparisons allowed us to extrapolate the caloric values gained from each prey type to estimated energetic requirements of pregnant and lactating female gray whales (Villegas-Amtmann et al. 2017).
Figure 2. Median caloric content and interquartile ranges by (A) species, (B) reproductive stage, and (C) month. Sizes of the zooplankton images are scaled at actual ratios relative to one another.
So, what did we find? Our sampling along the Oregon coast revealed six predominant zooplankton species: two mysid shrimp (Neomysis rayii, Holmesimysis sculpta), two amphipods (Atylus tridens, Polycheria osborni), and two types of crab larvae (Dungeness crab megalopae, porcelain crab larvae). These six Oregon prey species showed significant differences in their caloric values, with N. rayii and Dungeness crab megalopae having significantly higher calories per gram than the other prey species (Figure 2), though Dungeness crab megalopae stood out as the caloric gold mines for feeding gray whales in the PCFG range. Furthermore, month and reproductive stage also influenced the caloric content of some prey species, with gravid (aka pregnant) female mysid shrimp significantly increasing in calories throughout the summer (Figure 3).
Figure 3. Caloric content of different reproductive stages as a function of day of year (DOY; ranging from June to October) for the mysids Holmesimysis sculpta and Neomysis rayii, and the amphipod Atylus tridens. A. tridens is only represented on one panel due to small sample size of this species for the empty brood pouch and gravid reproductive stages. Asterisks indicate significant regressions (p<0.05).
The comparison of our Oregon prey caloric values to the predominant Arctic amphipod (Ampelisca macrocephala) proved our hypothesis wrong: Arctic amphipods do not have higher caloric value than Oregon prey, which would have help to explain why many more gray whales feed in the Arctic. We found that two Oregon prey species (N. rayii and Dungeness crab megalopae) have higher caloric values than A. macrocephala. If we translate the caloric contents of these prey to gray whale energetic needs, these differences mean that lactating and pregnant gray whales feeding in the PCFG area would need between 0.7-1.03 and 0.22-0.33 metric tons of prey less per day if they fed on Dungeness crab megalopae or N. rayii, respectively, than a whale feeding on Arctic A. macrocephala (Figure 4).
Figure 4. Daily prey requirements (A: metric tons; B: number of individuals) needed by pregnant and lactating female gray whales to meet their energetic requirements on the foraging ground. Energetic requirement estimates obtained from Villegas-Amtmann et al. (2017). Note the logarithmic scale of y-axis in panel (B).
If quality were the only prey metric that gray whales used to evaluate which food to eat, then it would make very little sense for so many gray whales to migrate to the Arctic when there are prey types of equal and greater quality available to them in the PCFG range. However, quality is not the only metric that influences gray whale foraging decisions. We therefore posit that the abundance, density, and availability of benthic amphipods in the Arctic are higher than the prey species found in the PCFG range. In fact, knowledge of the pulsed reproductive cycle of Dungeness and porcelain crabs allows us to conclude that the larvae of these two species are only available for a few weeks in the late spring and early summer on the Oregon coast. While mysid shrimp, such as N. rayii, are continuously available in the PCFG range throughout the summer, they may occur in less dense and more patchy aggregations than Arctic benthic amphipods. However, current estimates of prey density and abundance for either region are not available, and we do not have data on the energetic costs of the different foraging strategies. While there are still several unknowns, we have documented that higher prey quality in the Arctic is not the reason for the difference in gray whale foraging ground use in the eastern North Pacific.
References
Calambokidis, J., & Perez, A. 2017. Sightings and follow-up of mothers and calves in the PCFG and implications for internal recruitment. IWC Report SC/A17/GW/04 for the Workshop on the Status of North Pacific Gray Whales (La Jolla: IWC).
Calambokidis, J., Laake, J., & Perez, A. 2017. Updated analysis of abundance and population structure of seasonal gray whales in the Pacific Northwest, 1996-2015. IWC Report SC/A17/GW/05 for the Workshop on the Status of North Pacific Gray Whales (La Jolla: IWC).
Soledade Lemos, L., Burnett, J. D., Chandler, T. E., Sumich, J. L., & Torres, L. G. 2020. Intra- and inter-annual variation in gray whale body condition on a foraging ground. Ecosphere 11(4):e03094.
Stewart, J. D., & Weller, D. W. 2021. Abundance of eastern North Pacific gray whales 2019/2020. Department of Commerce, NOAA Technical Memorandum NMFS-SWFSC-639. United States: NOAA. doi:10.25923/bmam-pe91.
Villegas-Amtmann, S., Schwarz, L. K., Gailey, G., Sychenko, O., & Costa, D. P. 2017. East or west: the energetic cost of being a gray whale and the consequence of losing energy to disturbance. Endangered Species Research 34:167-183.
Since early May, much of the GEMM Lab has been consumed by the GRANITE project, which stands for Gray whale Response to Ambient Noise Informed by Technology and Ecology. Two weeks ago, PhD student Clara Bird discussed our field work preparations, and since May 20th we have conducted five successful days of field work (and one unsuccessful day due to fog). If you are now expecting a blog about the data we have collected so far and whales we encountered, I am sorry to disappoint you. Rather, I want to take a big step back and provide the context of the GRANITE project as a whole, explain why this project and data collection is so important, and discuss what it is that we hope to achieve with our ever-growing, multidisciplinary dataset and team.
We use the Pacific Coast Feeding Group (PCFG) of gray whales that forage off the Oregon coast as our study system to better understand the ecological and physiological response of baleen whales to multiple stressors. Our field methodology includes replicate physiological and ecological sampling of this accessible baleen whale population with synoptic measurement of multiple types of stressors. We collect fecal samples for hormone analysis, conduct drone overflights of whales to collect body condition and behavioral data, record the ambient soundscape through deployment of two hydrophones, and conduct whale photo-identification to link all data streams to each individual whale of known sex, estimated age, and reproductive status. We resample these data from multiple individuals within and between summer foraging seasons, while exposed to different potential stressors occurring at different intensities and temporal periods and durations. The hydrophones are strategically placed with one in a heavily boat-trafficked (and therefore noisy) area close to the Port of Newport, while the second is located in a relatively calm (and therefore quieter) spot near the Otter Rock Marine Reserve (Fig. 1). These hydrophones provide us with information about both natural (e.g. killer whales, wind, waves) and anthropogenic (e.g. boat traffic, seismic survey, marine construction associated with PacWave wave energy facility development) noise that may affect gray whales. During sightings with whales, we also drop GoPro cameras and sample for prey to better understand the habitats where whales forage and what they might be consuming.
Figure 1. Map of GRANITE study area from Seal Rock to Lincoln City with gray whale sightings (yellow circles) and and fecal samples collected (red triangles) from the 2020 field season. Green stars represent the two hydrophone locations. Source: L. Torres.
GEMM Lab PI Dr. Leigh Torres initiated this research project in 2015 and established partnerships with acoustician Dr. Joe Haxel and (then) PhD student Dr. Leila Lemos. Since then, the team working on this project has grown considerably to provide expertise in the various disciplines that the project integrates. Leigh is currently joined at the GRANITE helm by 4 co-PIs: Dr. Haxel, endocrinologist Dr. Kathleen Hunt, biological statistician Dr. Leslie New, and physiologist Dr. Loren Buck. Drs. Alejandro Fernandez Ajo, KC Bierlich and Enrico Pirotta are postdoctoral scholars who are working on the endocrinology, photogrammetry, and biostatistical modelling components, respectively. Finally, Clara and myself are partially funded through this project for our PhD research, with Clara focusing on the links between behavior, body condition, individualization, and habitat, while I am tackling questions about the recruitment and site fidelity of the PCFG (more about these topics below).
Faculty Research Assistant Todd Chandler supervises PhD student Clara Bird during her maiden drone flight over a whale. Source: L. Torres.
The ultimate goal of this project is to use the PCFG as a case study to quantify baleen whale physiological response to different stressors and model the subsequent impacts on the population by implementing our long-term, replicate dataset into a framework called Population consequences of disturbance (PCoD; Fig. 2). PCoD is built upon the underlying concept that changes in behavior and/or physiology caused by disturbance (i.e. noise) affect the fitness of individuals by impacting their health and vital rates, such as survival, reproductive success, and growth rate (Pirotta et al. 2018). These impacts at the individual level may (or may not) affect the population as a whole, depending on what proportion of individuals in the population are affected by the disturbance and the intensity of the disturbance effect on each individual. The PCoD framework requires quantification of four stages: a) the physiological and/or behavioral changes that occur as a result of exposure to a stressor (i.e. noise), b) the acute effects of these physiological and/or behavioral responses on individual vital rates, and their chronic effects via individual health, c) the way in which changes in health may affect the vital rates of individuals, and d) how changes in individual vital rates may affect population dynamics (Fig. 2; Pirotta et al. 2018). While four stages may not sound like a lot, the amount and longevity of data needed to quantify each stage is immense.
Figure 2. Conceptual framework of the population consequences of disturbance (PCoD). Letters (A-D) represent the four stages that require quantification in order for PCoD to be implemented. Each colored box represents external (ecological drivers, stressors) and internal (physiology, health, vital rates, behavior) factors that can change over time that are measured for each individual whale (dashed grey boundary line). The effects are then integrated across all individuals in the population to project their effects on the population’s dynamics. Figure and caption adapted from Pirotta et al. 2018.
The ability to detect a change in behavior or physiology often requires an understanding of what is “normal” for an individual, which we commonly refer to as a baseline. The best way to establish a baseline is to collect comprehensive data over a long time period. With our data collection efforts since 2015 of fecal samples, drone flights and photo identification, we have established useful baselines of behavioral and physiological data for PCFG gray whales. These baselines are particularly impressive since it is typically difficult to collect repeated measurements of hormones and body condition from the same individual baleen whale across multiple years. These repeated measurements are important because, like all mammals, hormones and body condition vary across life history phases (i.e., with pregnancy, injury, or age class) and across time (i.e., good or bad foraging conditions). To achieve these repeated measurements, GRANITE exploits the high degree of intra- and inter-annual site fidelity of the PCFG, their accessibility for study due to their affinity for nearshore habitat use, and the long-term sighting history of many whales that provides sex and approximate age information. Our work to-date has already established a few important baselines. We now know that the body condition of PCFG gray whales increases throughout a foraging season and can fluctuate considerably between years (Soledade Lemos et al. 2020). Furthermore, there are significant differences in body condition by reproductive state, with calves and pregnant females displaying higher body conditions (Soledade Lemos et al. 2020). Our dataset has also allowed us to validate and quantify fecal steroid and thyroid hormone metabolite concentrations, providing us with putative thresholds to identify a stressed vs. not stressed whale based on its hormone levels (Lemos et al. 2020).
PhD student Lisa Hildebrand and GRANITE co-PI Dr. Kathleen Hunt collecting a fecal sample. Source: L. Torres.
We continue to collect data to improve our understanding of baseline PCFG physiology and behavior, and to detect changes in their behavior and physiology due to disturbance events. All these data will be incorporated into a PCoD framework to scale from individual to population level understanding of impacts. However, more data is not the only thing we need to quantify each of the PCoD stages. The implementation of the PCoD framework also depends on understanding several aspects of the PCFG’s population dynamics. Specifically, we need to know whether recruitment to the PCFG population occurs internally (calves born from “PCFG mothers” return to the PCFG) or externally (immigrants from the larger Eastern North Pacific gray whale population joining the PCFG as adults). The degree of internal or external recruitment to the PCFG population should be included in the PCoD model as a parameter, as it will influence how much individual level disturbance effects impact the overall health and viability of the population. Furthermore, knowing residency times and home ranges of whales within the PCFG is essential to understand exposure durations to disturbance events.
To assess both recruitment and residency patterns of the PCFG, I am undertaking a large photo-identification effort, which includes compiling sightings and photo data across many years, regions, and collaborators. Through this effort we aim to identify calves and their return rate to the population, the rate of new adult recruits to the population, and the spatial residency of individuals in our study system. Although photo-id is a basic, commonplace method in marine mammal science, its role is critical to tracking individuals over time to understand population dynamics (in a non-invasive manner, no less). A large portion of my PhD research will focus on the tedious yet rewarding task of photo-id data management and matching in order to address these pressing knowledge gaps on PCFG population dynamics needed to implement the PCoD model that is an ultimate goal of GRANITE. I am just beginning this journey and have already pinpointed many analytical and logistical hurdles that I need to overcome. I do not anticipate an easy path to addressing these questions, but I am extremely eager to dig into the data, reveal the patterns, and integrate the findings into our rock-solid GRANITE project.
Funding for the GRANITE project comes from the Office of Naval Research, the Department of Energy, Oregon Sea Grant, the NOAA/NMFS Ocean Acoustics Program, and the OSU Marine Mammal Institute.
References
Lemos, L.S., Olsen, A., Smith, A., Chandler, T.E., Larson, S., Hunt, K., and L.G. Torres. 2020. Assessment of fecal steroid and thyroid hormone metabolites in eastern North Pacific gray whales. Conservation Physiology 8:coaa110.
Pirotta, E., Booth, C.G., Costa, D.P., Fleishman, E., Kraus, S.D., Lusseau, D., Moretti, D., New, L.F., Schick, R.S., Schwarz, L.K., Simmons, S.E., Thomas, L., Tyack, P.L., Weise, M.J., Wells, R.S., and J. Harwood. 2018. Understanding the population consequences of disturbance. Ecology and Evolution 8(19):9934-9946.
Soledade Lemos, L., Burnett, J.D., Chandler, T.E., Sumich, J.L., and L.G. Torres. 2020. Intra- and inter-annual variation in gray whale body condition on a foraging ground. Ecosphere 11(4):e03094.
Clara Bird, PhD Student, OSU Department of Fisheries, Wildlife, and Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
When I thought about what doing fieldwork would be like, before having done it myself, I imagined that it would be a challenging, but rewarding and fun experience (which it is). However, I underestimated both ends of the spectrum. I simultaneously did not expect just how hard it would be and could not imagine the thrill of working so close to whales in a beautiful place. One part that I really did not consider was the pre-season phase. Before we actually get out on the boats, we spend months preparing for the work. This prep work involves buying gear, revising and developing protocols, hiring new people, equipment maintenance and testing, and training new skills. Regardless of how many successful seasons came before a project, there are always new tasks and challenges in the preparation phase.
For example, as the GEMM Lab GRANITE project team geared up for its seventh field season, we had a few new components to prepare for. Just to remind you, the GRANITE (Gray whale Response to Ambient Noise Informed by Technology and Ecology) project’s field season typically takes place from June to mid-October of each year. Throughout this time period the field team goes out on a small RHIB (rigid hull inflatable boat), whenever the weather is good enough, to collect photo-ID data, fecal samples, and drone imagery of the Pacific Coast Feeding Group (PCFG) gray whales foraging near Newport, OR, USA. We use the data to assess the health, ecology and population dynamics of these whales, with our ultimate goal being to understand the effect of ambient noise on the population. As previous blogs have described, a typical field day involves long hours on the water looking for whales and collecting data. This year, one of our exciting new updates is that we are going out on two boats for the first part of the field season and starting our season 10 days early (our first day was May 20th). These updates are happening because a National Science Foundation funded seismic survey is being conducted within our study area starting in June. The aim of this survey is to assess geophysical structures but provides us with an opportunity to assess the effect of seismic noise on our study group by collecting data before, during, and after the survey. So, we started our season early in order to capture the “before seismic survey” data and we are using a two-boat approach to maximize our data collection ability.
While this is a cool opportunistic project, implementing the two-boat approach came with a new set of challenges. We had to find a second boat to use, buy a new set of gear for the second boat, figure out the best way to set up our gear on a boat we had not used before, and update our data processing protocols to include data collected from two boats on the same day. Using two boats also means that everyone on the core field team works every day. This core team includes Leigh (lab director/fearless leader), Todd (research assistant), Lisa (PhD student), Ale (new post-doc), and me (Clara, PhD student). Leigh and Todd are our experts in boat driving and working with whales, Todd is our experienced drone pilot, I am our newly certified drone pilot, and Lisa, Ale, and myself are boat drivers. Something I am particularly excited about this season is that Lisa, Ale, and I all have at least one field season under our belts, which means that we get to become more involved in the process. We are learning how to trailer and drive the boats, fly the drones, and handling more of the post-field work data processing. We are becoming more involved in every step of a field day from start to finish, and while it means taking on more responsibility, it feels really exciting. Throughout most of graduate school, we grow as researchers as we develop our analytical and writing skills. But it’s just as valuable to build our skillset for field work. The ocean conditions were not ideal on the first day of the field season, so we spent our first day practicing our field skills.
For our “dry run” of a field day, we went through the process of a typical day, which mostly involved a lot of learning from Leigh and Todd. Lisa practiced her trailering and launching of the boat (figure 1), Ale and Lisa practiced driving the boat, and I practiced flying the drone (figure 2). Even though we never left the bay or saw any whales, I thoroughly enjoyed our dry run. It was useful to run through our routine, without rushing, to get all the kinks out, and it also felt wonderful to be learning in a supportive environment. Practicing new skills is stressful to say the least, especially when there is expensive equipment involved, and no one wants to mess up when they’re being watched. But our group was full of support and appreciation for the challenges of learning. We cheered for successful boat launchings and dockings, and drone landings. I left that day feeling good about practicing and improving my drone piloting skills, full of gratitude for our team and excited for the season ahead.
Figure 1. Lisa (driving the truck) launching the boat.
Figure 2. Clara (seated, wearing a black jacket) landing the drone in Ale’s hands.
All the diligent prep work paid off on Saturday with a great first day (figure 3). We conducted five GoPro drops (figure 4), collected seven fecal samples from four different whales (figure 5), and flew four drone flights over three individuals including our star from last season, Sole. Combined, we collected two trifectas (photo-ID images, fecal samples, and drone footage)! Our goal is to get as many trifectas as possible because we use them to study the relationship between the drone data (body condition and behavior) and the fecal sample data (hormones). We were all exhausted after 10 hours on the water, but we were all very excited to kick-start our field season with a great day.
Figure 3. Lisa on the bow pulpit during our first sighting of the day.
Figure 4. Lisa doing a GoPro drop, she’s lowering the GoPro into the water using the line in her hands.
Figure 5. Clara and Ale collecting a fecal sample.
On Sunday, just one boat went out to collect more data from Sole after a rainy morning and I successfully flew over her from launching to landing! We have a long season ahead, but I am excited to learn and see what data we collect. Stay tuned for more updates from team GRANITE as our season progresses!
Last week marked the one year anniversary of the pandemic reality we have all been living. It has been an extremely challenging year, with everyone experiencing different kinds of difficulties and hurdles. One challenge that likely unites the majority of us is having to forego seeing our loved ones. For me personally, this is the longest time I have not seen my family (445 days and counting) and I know I am not alone in this situation. My homesickness started a train of thought about cetacean parental care and inspired me to write a blog about this topic. As you can see from the title, this post focuses on maternal care, rather than parental care. This bias isn’t due to my lack of research on this topic or active exclusion, but rather because there are currently no known cetacean species where paternal participation in offspring production and development extends beyond copulation (Rendell et al. 2019). Thus, this blog is all about the role of mothers in the lives of cetacean calves.
Like humans, cetacean mothers invest a lot of energy into their offspring. Most species have a gestation period of 10 or more months (Oftedal 1997). For baleen whale females in particular, pregnancy is not an easy feat given that they only feed during summer feeding seasons. They must therefore acquire all of the energy they will need for two migrations, birth, and (almost) complete lactation, before they will have access to food on feeding grounds again. For pregnant gray whales, a mere 4% loss in average energy intake on the foraging grounds will prevent her from successfully producing and/or weaning a calf (Villegas-Amtmann et al. 2015), demonstrating how crucial the foraging season is for a pregnant baleen whale. Once a calf is born, lactation ensues, ranging in length between approximately 6-8 months for most baleen whale species to upwards of one or two years in odontocetes (Oftedal 1997). The very short lactation period in baleen whales is offset by the large volume (for blue whales, up to 220 kg per day) and high fat percentage (30-50%) of milk that mothers provide for their calves (Oftedal 1997). In contrast, odontocetes (or toothed whales) have a more prolonged period of lactation with less fatty milk (10-30%). This discrepancy in lactation period lengths is in part because odontocete species do not undertake long migrations, which allows females to feed year-round and therefore allocate energy to nursing young for a longer time.
Blue whale calf nursing in New Zealand in 2016. Footage captured via unmanned aerial system (UAS; drone) piloted by Todd Chandler for GEMM Lab’s OBSIDIAN project. Source: GEMM Lab.
Aside from the energetically costly task of lactation, cetacean mothers must also assist their calves as they learn to swim. Echelon swimming is a common position of mother-calf pairs whereby the calf is in very close proximity to its mother’s mid-lateral flank and provides calves with hydrodynamic benefits. Studies in bottlenose dolphins have shown that swimming in echelon results in a 24% reduction in mean maximum swim speeds and a 13% decrease in distance per stroke (Noren 2008) for mothers, while concurrently increasing average swim speeds and distance per stroke of calves by 28% and 19%, respectively (Noren et al. 2007). While these studies have only been conducted in odontocete species, echelon swimming is also observed in baleen whales (Smultea et al. 2017), indicating that baleen whale females may experience the same reductions in swimming efficiency. Furthermore, mothers will forgo sleep in the first days after birth (killer whales & bottlenose dolphins; Lyamin et al. 2005) and/or shorten their dive foraging times to accommodate calf diving ability (bottlenose dolphins [Miketa et al. 2018] & belugas [Heide-Jørgensen et al. 2001]). Females must endure these losses in foraging opportunities and decreased swimming efficiency when they are at their most nutritionally stressed to ensure the well-being and success of their offspring.
It is at the time of weaning (when a calf becomes independent), that we start to see differences in the maternal role between baleen and toothed whale mothers. Odontocetes have much stronger sociality than baleen whales causing offspring to stay with their mothers for much longer periods. Among the largest toothed whales, such as killer and sperm whales, offspring stay with their mothers in stable matrilineal units for often a lifetime. Among the smaller toothed whales, such as bottlenose dolphins, maternal kin maintain strong bonds in dynamic fission-fusion societies. In contrast, post-weaning maternal care in baleen whales is limited, with the mother-calf pair typically separating soon after the calf is weaned (Rendell et al. 2019).
Conceptual diagram depicting where baleen (Mysticeti) and toothed (Odontoceti) whales fall on the continuum of low to high social structure and matrilineal kinship structure. The networks at the top depict long-term datasets of photo-identified individuals (red nodes = females, blue nodes = males, yellow nodes = calves) with thickness of connecting lines representing strength of association between individuals. Figure and caption [adapted] from Rendell et al. 2019.
The long-term impact of social bonds in odontocetes is evident through examples of vertically transmitted behaviors (from mother to calf) in a number of species. For example, the use of three unique foraging tactics (sponge carrying, rooster-tail foraging, and mill foraging) by bottlenose dolphin calves in Shark Bay, Australia, was only significantly explained by maternal use of these tactics (Sargeant & Mann 2009). In Brazil, individuals of four bottlenose dolphin populations along the coast cooperatively forage with artisanal fishermen, which involves specialized and coordinated behaviors from both species. This cooperative foraging tactic among dolphins is primarily maintained across generations via social learning from mothers to calves (Simões-Lopeset al. 2016). The risky tactic of intentional stranding by killer whales on beaches to capture elephant seal pups requires a high degree of skill and high parental investment to reduce the associated risk of stranding (Guinet & Bouvier 1995).
Evidence for vertical transmission of specialized foraging tactics in baleen whales currently does not exist. Bubble-net feeding is a specialized tactic employed by humpback whales in three oceanic regions where multiple individuals work together to herd and trap prey (Wiley et al. 2011). However, it remains unknown whether this behavior is vertically transmitted. Simultaneous video tags from a mother-calf humpback whale pair in the Western Antarctic Peninsula documented synchrony in dives, with the calf’s track lagging behind the mother’s by 4.5 seconds, suggesting that the calf was following its mother (Tyson et al. 2012). Synchronous diving likely allows calves to observe their mothers and practice their diving, and could offer a pathway for them to mimic foraging behaviors and tactics displayed by mothers.
While there currently may not be evidence for vertical transmission of specialized foraging tactics among the baleen whales, there is documentation of matrilineal fidelity to both foraging (Weinrich 1998, Barendse et al. 2013, Burnham & Duffus 2020) and breeding grounds (Carroll et al. 2015). Matrilineal site fidelity to foraging grounds is not exclusive to baleen whales and has also been documented in a number of odontocete species (Palsbøll et al. 1997, Turgeon et al. 2012).
A gallery of some GEMM Lab documented mother-calf gray whale pairs. All images captured under NOAA/NMFS permit #21678. Exclamation (mom; left) and Angie (calf; right).
Scarlett (mom; left) and Brown (calf; right).
Clouds (mom; back) and Cheetah (calf; front).
Xena (mom; back) and Evie (calf; front).
Knife (mom; left) and Daffodil (calf; right).
Pristine (mom; front) and Three Stars (calf; back).
Triplet (mom; right) and White Knuckle (calf; left).
In the GEMM Lab, we are interested in exploring the potential long-term bonds, role and impact of Pacific Coast Feeding Group (PCFG) gray whale mothers on their calves. GEMM Lab PhD student Clara Bird is digging into whether specialized foraging tactics, such as bubble blasts and headstands, are passed down from mothers to calves. I hope to assess whether using the PCFG range as a foraging ground (rather than the Arctic region) is a vertically transmitted behavior or whether environmental factors may play a larger role in the recruitment and dynamics of the PCFG. It will take us a while to get to the bottom of these questions, so in the meantime hug your loved ones if it’s safe to do so or, if you’re in my boat, continue to talk to them virtually until it is safe to be reunited.
References
Barendse, J., Best, P. B., Carvalho, I., and C. Pomilla. 2013. Mother knows best: occurrence and associations of resighted humpback whales suggest maternally derived fidelity to a southern hemisphere coastal feeding ground. PloS ONE 8:e81238.
Burnham, R. E., and D. A. Duffus. 2020. Maternal behaviors of gray whales (Eschrichtius robustus) on a summer foraging site. Marine Mammal Science 36:1212-1230.
Carroll, E. L., Baker, C. S., Watson, M., Alderman, R., Bannister, J., Gaggiotti, O. E., Gröcke, D. R., Patenaude, N., and R. Harcourt. 2015. Cultural traditions across a migratory network shape the genetic structure of southern right whales around Australia and New Zealand. Scientific Reports 5:16182.
Guinet, C., and J. Bouvier. 1995. Development of intentional stranding hunting techniques in killer whale (Orcinus orca) calves at Crozet Archipelago. Canadian Journal of Zoology 73:27-33.
Heide-Jørgensen, M. P., Hammeken, N., Dietz, R., Orr, J., and P. R. Richard. 2001. Surfacing times and dive rates for narwhals and belugas. Arctic 54:207-355.
Lyamin, O., Pryaslova, J., Lance, V., and J. Siegel. 2005. Continuous activity in cetaceans after birth. Nature 435:1177.
Miketa, M. L., Patterson, E. M., Krzyszczyk, E., Foroughirad, V., and J. Mann. 2018. Calf age and sex affect maternal diving behavior in Shark Bay bottlenose dolphins. Animal Behavior 137:107-117.
Noren, S. R. 2008. Infant carrying behavior in dolphins: costly parental care in an aquatic environment. Functional Ecology 22:284-288.
Noren, S. R., Biedenbach, F., Redfern, J. V., and E. F. Edwards. 2007. Hitching a ride: the formation locomotion strategy of dolphin calves. Functional Ecology 22:278-283.
Oftedal, O. T. Lactation in whales and dolphins: evidence of divergence between baleen- and toothed-species. Journal of Mammary Gland Biology and Neoplasia 2:205-230.
Palsbøll, P. J., Heide-Jørgensen, M. P., and R. Dietz. 1996. Population structure and seasonal movements of narwhals, Monodon monoceros, determined from mtDNA analysis. Heredity 78:284-292.
Rendell, L., Cantor, M., Gero, S., Whitehead, H., and J. Mann. 2019. Causes and consequences of female centrality in cetacean societies. Philosophical Transactions of the Royal Society B 374:20180066.
Sargeant, B. L., and J. Mann. 2009. Developmental evidence for foraging traditions in wild bottlenose dolphins. Animal Behavior 78:715-721.
Simões-Lopes, P. C., Daura-Jorge, F. G., and M. Cantor. 2016. Clues of cultural transmission in cooperative foraging between artisanal fishermen and bottlenose dolphins, Tursiops truncatus (Cetacea: Delphinidae). Zoologia (Curitiba) 33:e20160107.
Smultea, M. A., Fertl, D., Bacon, C. E., Moore, M. R., James, V. R., and B. Würsig. 2017. Cetacean mother-calf behavior observed from a small aircraft off Southern California. Animal Behavior and Cognition 4:1-23.
Turgeon, J., Duchesne, P., Colbeck, G. J., Postma, L. D., and M. O. Hammill. 2011. Spatiotemporal segregation among summer stocks of beluga (Delphinapterus leucas) despite nuclear gene flow: implication for the endangered belugas in eastern Hudson Bay (Canada). Conservation Genetics 13:419-433.
Tyson, R. B., Friedlaender, A. S., Ware, C., Stimpert, A. K., and D. P. Nowacek. 2012. Synchronous mother and calf foraging behaviour in humpback whales Megaptera novaeangliae: insights from multi-sensor suction cup tags. Marine Ecology Progress Series 457:209-220.
Villegas-Amtmann, S., Schwarz, L. K., Sumich, J. L., and D. P. Costa. 2015. A bioenergetics model to evaluate demographic consequences of disturbance in marine mammals applied to gray whales. Ecosphere 6:1-19.
Weinrich, M. 1998. Early experience in habitat choice by humpback whales (Megaptera novaeaengliae). Journal of Mammalogy 79:163-170.
Wiley, D., Ware, C., Bocconcelli, A., Cholewiak, D., Friedlaender, A., Thompson, M., and M. Weinrich. 2011. Underwater components of humpback whale bubble-net feeding behavior. Behavior 148:575-602.
By Alejandro Fernandez Ajo, PhD student at the Department of Biology, Northern Arizona University, Visiting scientist in the GEMM Lab working on the gray whale physiology and ecology project
About four years ago, I was in Patagonia, Argentina deciding where to focus my research and contribute to whale conservation efforts. At the same time, I was doing fieldwork with the Whale Conservation Institute of Argentina at the “Whale Camp” in Península Valdés. I read tons of papers and talked with my colleagues about different opportunities and gaps in knowledge that I could tackle during my Ph.D. program. One of the questions that caught my attention was about the unknown cause (or causes) for the recurrent high calf mortalities that the Southern Right Whale (SRW) population that breeds at Peninsula Valdés experienced during the 2000s (Rowntree et al. 2013). Still, at that time, I was unsure how to tackle this research question.
Golfo San José, Península Valdés – Argentina. Collecting SRW behavioral data from the cliff’s vantage point. Source: A. Fernandez Ajo.
Between 2003 and 2013, at least 672 SRWs died, of which 91% were calves (Sironi et al. 2014). These mortalities represented an average total whale death per year of 80 individuals in the 2007-2013 period, which vastly exceeded the 8.2 average deaths per year of previous years by a ten-fold increase (i.e., 1993-2002) (Rowntree et al. 2013). In fact, this calf mortality rate was the highest ever documented for any population of large whales. During this period, from 2006 to 2009, I was the Coordinator of the Fauna Area in the Patagonian Coastal Zone Management Plan, and I collaborated with the Southern Right Whale Health Monitoring Program (AKA: The Stranding Program) that conducted field necropsies on stranded whales along the coasts of the Península and collected many different samples including whale baleen.
Southern Right Whale, found stranded in Patagonia Argentina. Source: Instituto de Conservación de Ballenas.
In this process, I learned about the emerging field of Conservation Physiology and the challenges of utilizing traditional approaches to studying physiology in large whales. Basically, the problem is that there is no possible way to obtain blood samples (the gold standard sample type for physiology) from free-swimming whales; whales are just too large! Fortunately, there are currently several alternative approaches for gathering physiological information on large whales using a variety of non-lethal and minimally invasive (or non-invasive) sample matrices, along with utilizing valuable samples recovered at necropsy (Hunt et al. 2013). That is how I learned about Dr. Kathleen Hunt’s novel research studying hormones from whale baleen (Hunt et al., 2018, 2017, 2014). Thus, I contacted Dr. Hunt and started a collaboration to apply these novel methods to understand the case of calf mortalities of the SRW calves in Patagonia utilizing the baleen samples that we recovered with the Stranding program at Península Valdés (see my previous blog post).
What is conservation physiology?
Conservation physiology is a multidisciplinary field of science that utilizes physiological concepts and tools to understand underlying mechanisms of disturbances to solve conservation problems. Conservation physiology approaches can provide sensitive biomarkers of environmental change and allow for targeted conservation strategies. The most common Conservation Physiology applications are monitoring environmental stressors, understanding disease dynamics and reproductive biology, and ultimately reducing human-wildlife conflict, among other applications.
I am now completing the last semester of my Ph.D. program. I have learned much about the amazing field of Conservation Physiology and how much more we need to know to achieve our conservation goals. I am still learning, yet I feel that through my research I have contributed to understanding how different stressors impact the health and wellbeing of whales, and about aspects of their biology that have long been obscured or unknown for these giants. One contribution I am proud of is our recent publication of, “A tale of two whales: putting physiological tools to work for North Atlantic and southern right whales,” which was published in January 2021 as a book chapter in “Conservation Physiology: Applications for Wildlife Conservation and Management” published by Oxford University Press: Oxford, UK.
This book outlines the significant avenues and advances that conservation physiology contributes to the monitoring, management, and restoration of wild animal populations. The book also defines opportunities for further growth in the field and identifies critical areas for future investigation. The text and the contributed chapters illustrate several examples of the different approaches that the conservation physiology toolbox can tackle. In our chapter, “A tale of two whales,” we discuss developments in conservation physiology research of large whales, with the focus on the North Atlantic right whale (Eubalaena glacialis) and southern right whale (Eubalaena australis), two closely related species that differ vastly in population status and conservation pressures. We review the advances in Conservation Physiology that help overcome the challenges of studying large whales via a suite of creative approaches, including photo-identification, visual health assessment, remote methods of assessing body condition, and endocrine research using non-plasma sample types such as feces, respiratory vapor, and baleen. These efforts have illuminated conservation-relevant physiological questions for both species, such as discrimination of acute from chronic stress, identification of likely causes of mortality, and monitoring causes and consequences of body condition and reproduction changes.
This book provides an overview of the different applications of Conservation Physiology, outlining the significant avenues and advances by which conservation physiology contributes to the monitoring, management, and restoration of wild animal populations. By using a series of global case studies, contributors illustrate how approaches from the conservation physiology toolbox can tackle a diverse range of conservation issues, including monitoring environmental stress, predicting the impact of climate change, understanding disease dynamics, and improving captive breeding, and reducing human-wildlife conflict. The variety of taxa, biological scales, and ecosystems is highlighted to illustrate the far-reaching nature of the discipline and allow readers to appreciate the purpose, value, applicability, and status of the field of conservation physiology. This book is an accessible supplementary textbook suitable for graduate students, researchers, and practitioners in conservation science, ecophysiology, evolutionary and comparative physiology, natural resources management, ecosystem health, veterinary medicine, animal physiology, and ecology.
References
Hunt KE, Fernández Ajó A, Lowe C, Burgess EA, Buck CL. 2021. A tale of two whales: putting physiological tools to work for North Atlantic and southern right whales. In: “Conservation Physiology: Integrating Physiology Into Animal Conservation And Management”, ch. 12. Eds. Madliger CL, Franklin CE, Love OP, Cooke SJ. Oxford University press: Oxford, UK.
Sironi, M., Rowntree, V., Di Martino, M. D., Beltramino, L., Rago, V., Franco, M., and Uhart, M. (2014). Updated information for 2012-2013 on southern right whale mortalities at Península Valdés, Argentina. SC/65b/BRG/06 report presented to the International Whaling Commission Scientific Committee, Portugal. <https://iwc.int/home>.
Rowntree, V.J., Uhart, M.M., Sironi, M., Chirife, A., Di Martino, M., La Sala, L., Musmeci, L., Mohamed, N., Andrejuk, J., McAloose, D., Sala, J., Carribero, A., Rally, H., Franco, M., Adler, F., Brownell, R. Jr, Seger, J., Rowles, T., 2013. Unexplained recurring high mortality of southern right whale Eubalaena australis calves at Península Valdés, Argentina. Marine Ecology Progress Series, 493, 275-289. DOI: 10.3354/meps10506
Hunt KE, Moore MJ, Rolland RM, Kellar NM, Hall AJ, Kershaw J, Raverty SA, Davis CE, Yeates LC, Fauquier DA, et al., 2013. Overcoming the challenges of studying conservation physiology in large whales: a review of available methods. Conserv Physiol 1: cot006–cot006.
Hunt, K.E., Stimmelmayr, R., George, C., Hanns, C., Suydam, R., Brower, H., Rolland, R.M., 2014. Baleen hormones: a novel tool for retrospective assessment of stress and reproduction in bowhead whales (Balaena mysticetus). Conserv. Physiol. 2, cou030. https://doi.org/10.1093/conphys/cou030
Hunt, K.E., Lysiak, N.S., Moore, M.J., Rolland, R.M., 2016. Longitudinal progesterone profiles in baleen from female North Atlantic right whales (Eubalaena glacialis) match known calving history. Conserv. Physiol. 4, cow014. https://doi.org/10.1093/conphys/cow014


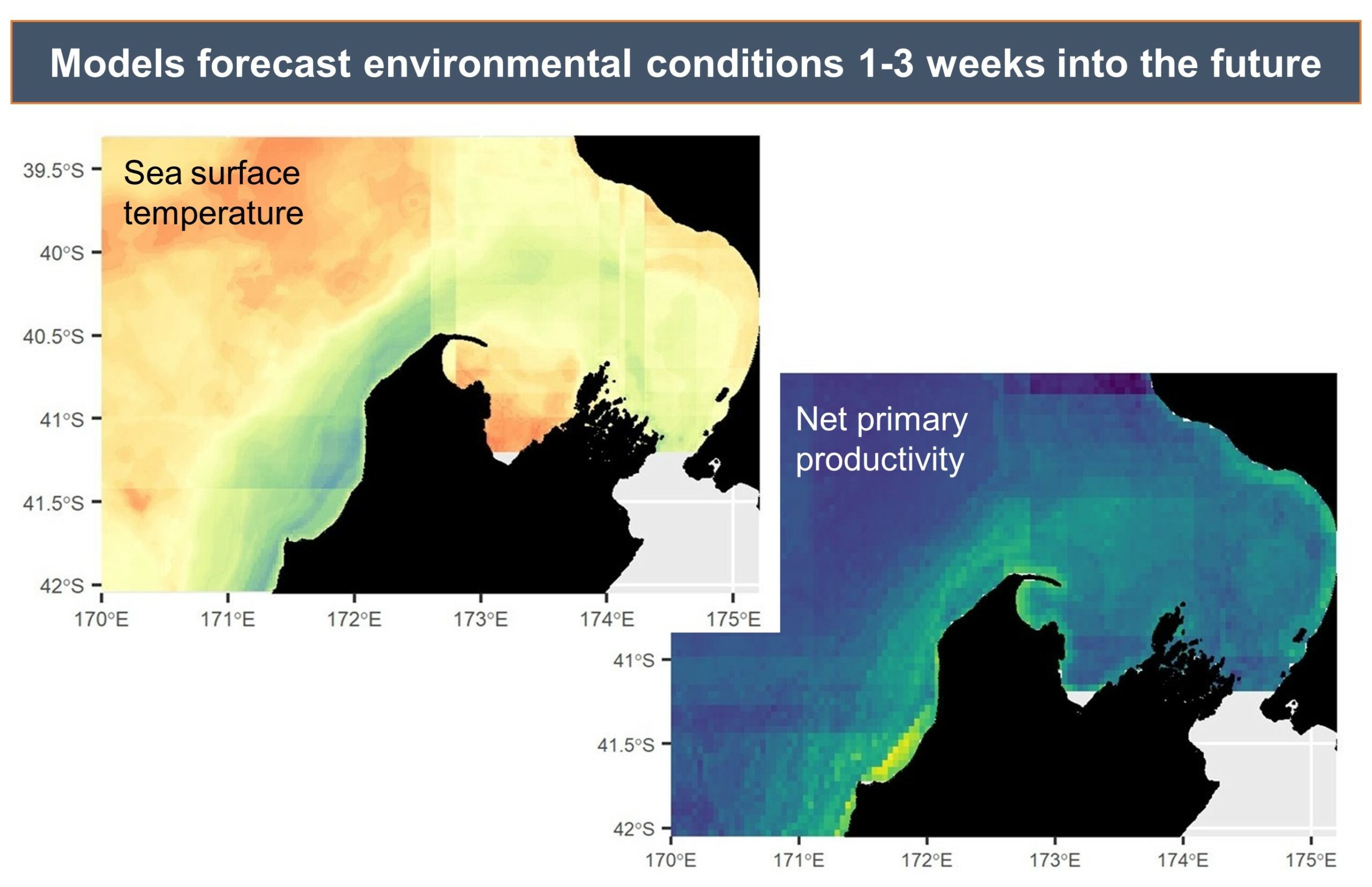





















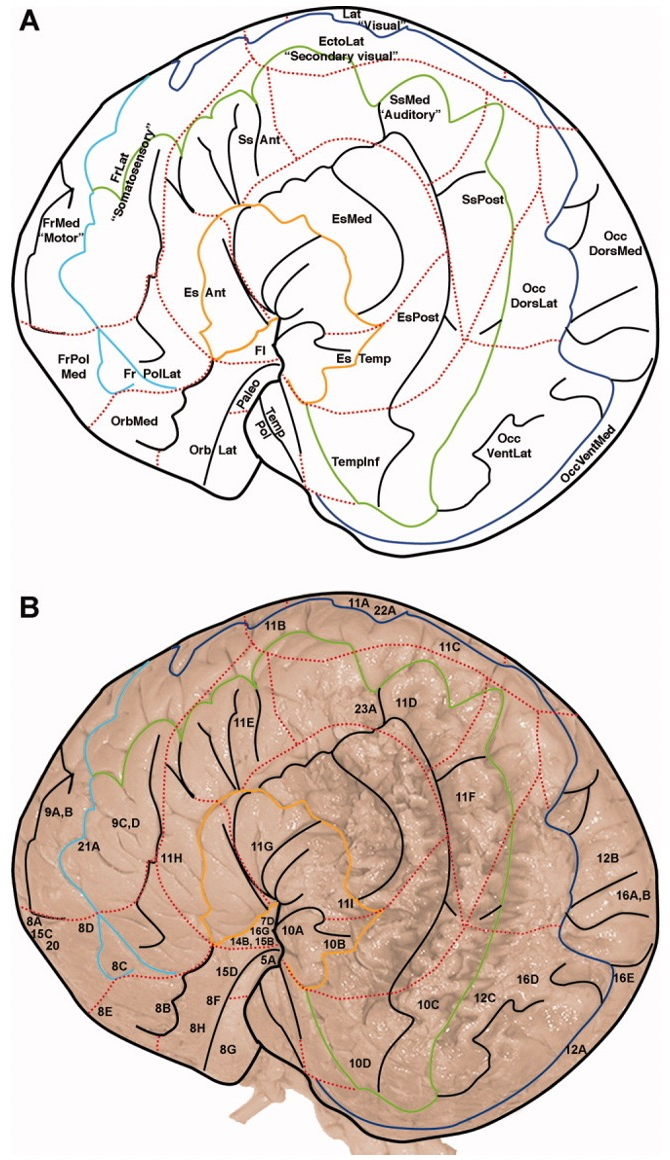
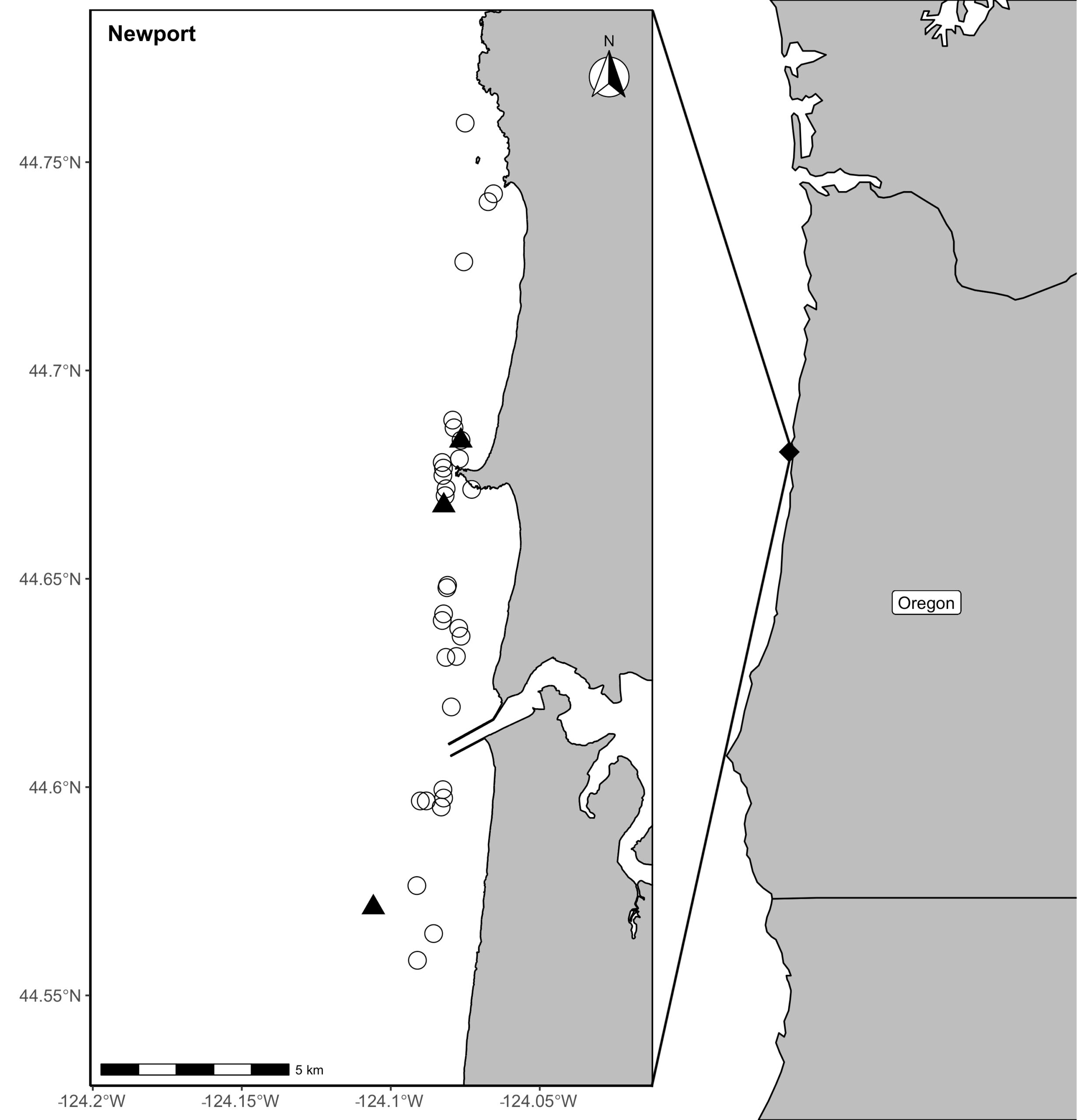
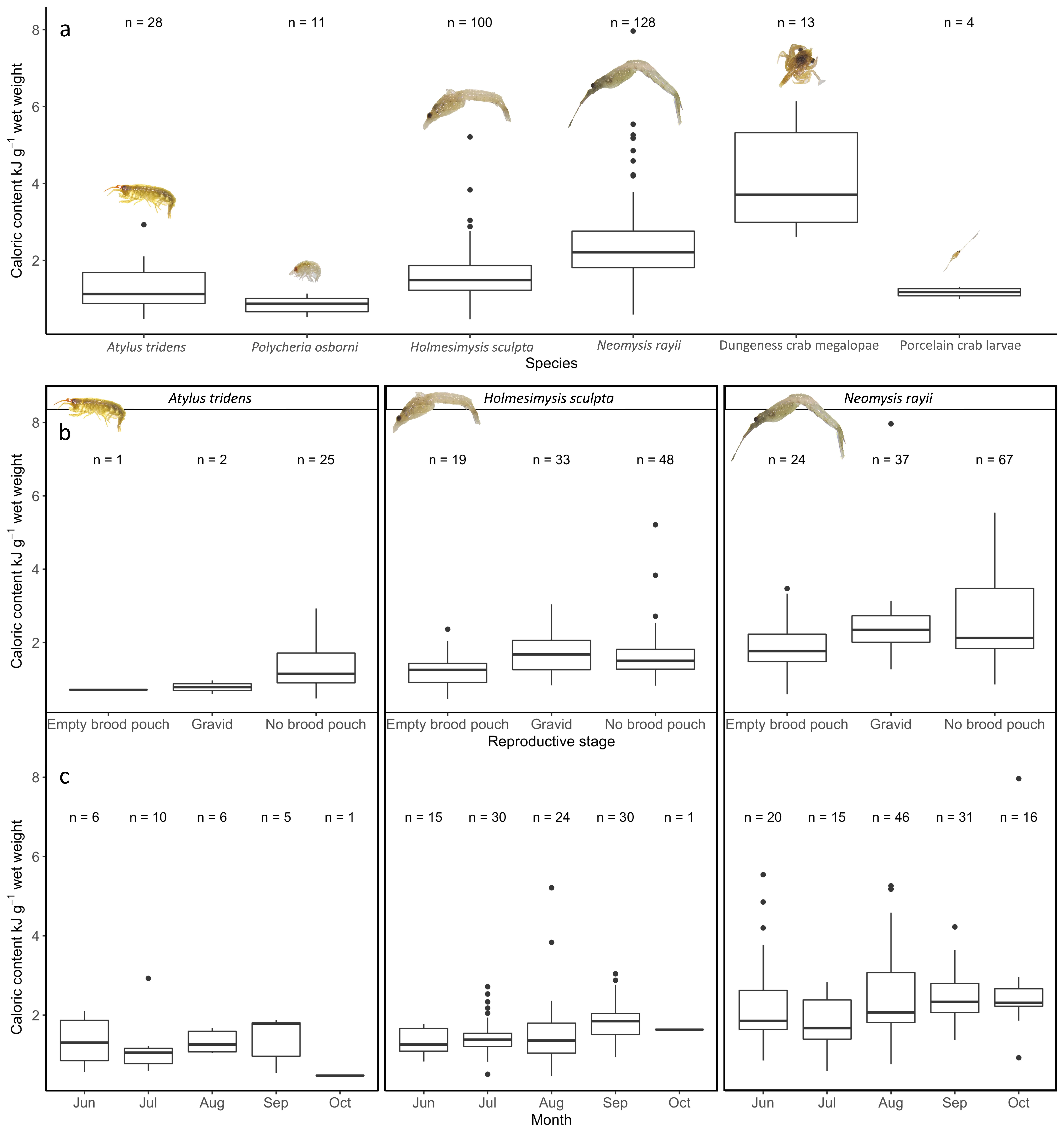


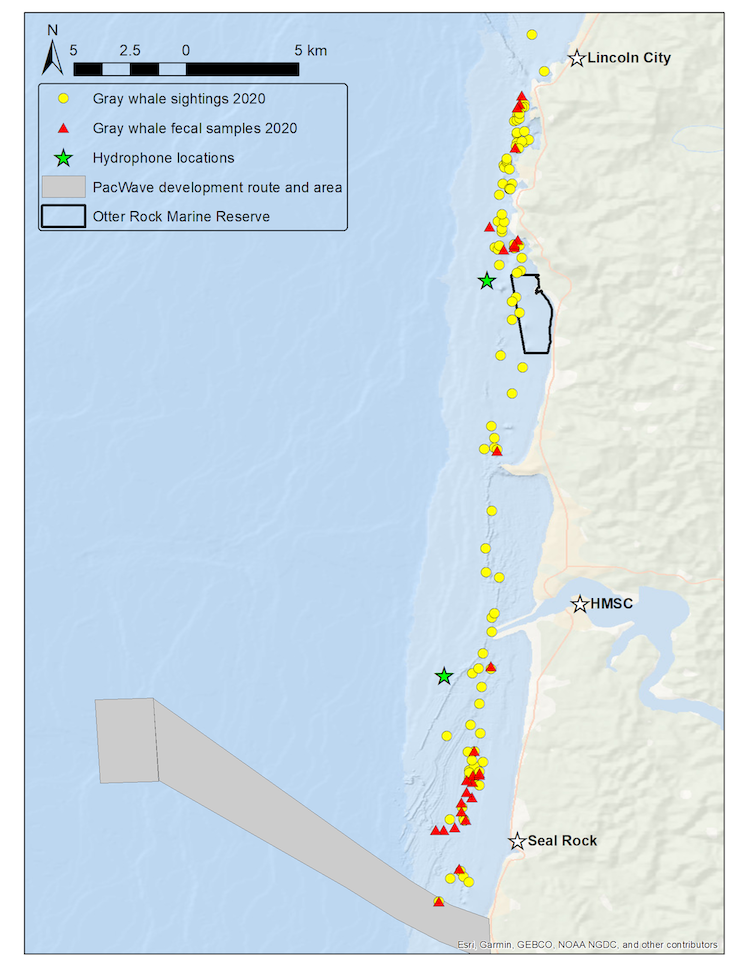

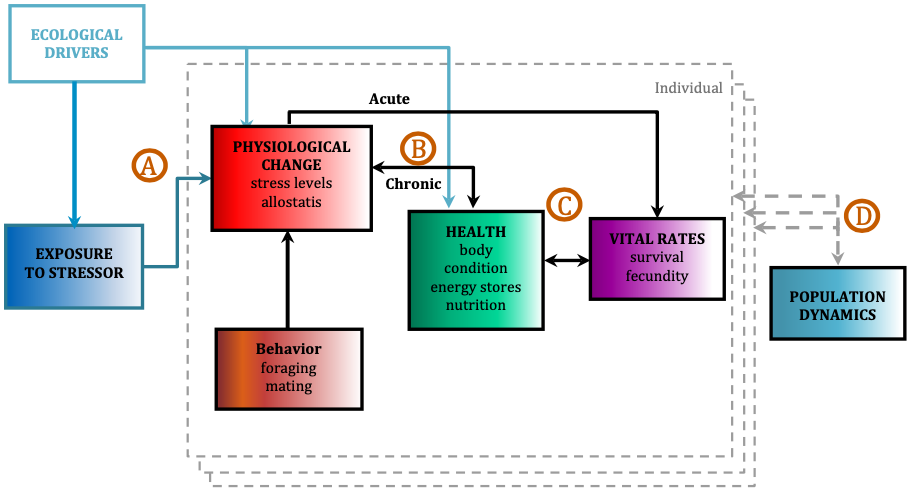


















{kind=link}