By Damian Amerman-Smith, Pacific High School senior, GEMM Lab summer intern
Left to right: Damian, Nadia, Jasen. The group scans the ocean looking for whales, while Damian puts on sunscreen. Source: A. Dawn.
Growing up in Port Orford, a short ten-minute walk from the Pacific Ocean, has certainly shaped my life a lot. It has given me a great regard for the ocean, the diversity of life within it, and how life seems to bypass human derived borders in order to go wherever it can. I often marvel at all the beautiful, intricate ecosystems that are able to exist inside of our planet’s vast oceanic expanses. Along with my love of the ocean has come a great regard for marine mammals and the novelties of these animals that allow them to live entirely in the ocean despite not having gills. Every new discovery of these beautiful ocean creatures brings me such simple and pure joy, such as my very recent discovery that baleen whales have two blow holes. These blow holes look so peculiar on the top of their bodies, like a short upside-down nose.
My interest in the ocean and its inhabitants was a large part of what made me so enthused to take a part in the gray whale foraging ecology (GWFE) project in Port Orford this summer. When Elizabeth Kelly, my friend and a previous intern for the GWFE project mentioned her experiences from the previous summer, I was very happy when she put me in contact with Lisa Hildebrand and Leigh Torres so that I could apply to be an intern. Since then, I have been very ecstatically awaiting the beginning of the project and could hardly believe it when it finally began, and I was able to meet my fellow team members: Lisa Hildebrand, the PhD student who has been leading the GWFE project for the last four years; Allison Dawn, a Master’s student who is going to take over the project in Lisa’s stead; Nadia Leal, an OSU undergrad hoping to further pursue the field of marine biology; and Jasen White, an OSU undergrad whose time in the Navy has made him a very steeling presence while out on the water.
The three weeks that we have spent together learning the procedures that make up the project have been well spent, teaching all of us a lot of new things, such as what a theodolite is, how to operate a dissolved oxygen sensor, and (for me) how to use Excel. The first two weeks were largely spent just learning about how we collect data and improving our field skills, but as we have become more comfortable with our skills, we have also begun looking beyond the procedures, towards the data itself and what it can mean. Primarily, we started to notice the distinct lack of gray whales and almost complete lack of zooplankton prey for any gray whales in the area to eat.
A calm & beautiful, yet whale-less, view from the cliff site. Source: L. Hildebrand.
As we pass the halfway point in the project, we have only witnessed two whales inside our study area. While in the beginning it was not surprising that there were no whales, it has started to become concerning to me. We have a strong working hypothesis about why there have not been many whale sightings in our monitored sites of Mill Rocks and Tichenor’s Cove: there is not nearly enough zooplankton prey to attract them. Monday, August 9th is a good example to support this hypothesis. On that day, when we pulled up our sample net at Tichenor Cove station #1, we collected fifty-three individual Neomysis mysid shrimp, which are a tasty treat for gray whales. However, all the other prey samples from the remaining eleven kayak sampling stations had perhaps a maximum of five assorted zooplankton each, which is certainly not enough to attract the attention of such a large predator as Eschrichtius robustus (a gray whale). Unfortunately, we have yet to see much change in zooplankton prey availability in our sampling nets over the season so far, but we are hopeful that swarms of zooplankton in the area will resurge and the gray whales will begin using the area around the port as their August feeding grounds.
Our hopes aside, it is intriguing to think about why there has been so few zooplankton at our sampling sites. A main factor is likely the decrease of Port Orford’s kelp forests over the past few years. Kelp is very important to zooplankton, particularly mysids, as it allows them to seek shelter from predators. Declines in kelp forests have been documented all along the southern Oregon coast, and are believed to be fueled by many factors (ORKA, 2021). A combination of warming waters with decreasing amount of nutrients available to the kelp (Richardson 2008), and the increasing abundances of purple sea urchins that eat the kelp has vastly impacted the amount of kelp in the area. The decline in local kelp forests may be the reason that we are seeing fewer mysid swarms than in previous years. This reduced kelp and mysid availability could, in turn, be making Port Orford waters an unappetizing area for hungry whales to visit this year. While this trophic cascade is still just an educated hypothesis, it is important for us and others to keep watch on the situation, and to see how it changes. There are organizations such as the Oregon Kelp Alliance (ORKA) that are working hard to study why the kelp populations are hurting and how we can help. We will power through the season with the hopes that the gray whales will come. It is still very possible that the zooplankton will resurge and the whales will return with plenty to feed on.
References
Richardson, Anthony J. 2008. In hot water: zooplankton and climate change, ICES Journal of Marine Science, Volume 65, Issue 3, Pages 279–295, https://doi.org/10.1093/icesjms/fsn028
By Nadia Leal, GEMM Lab summer intern, OSU senior, Department of Fisheries, Wildlife, and Conservation Sciences
The OSU GEMM Lab gray whale foraging ecology project in Port Orford is in its seventh year of research. I have the honor to serve as a field assistant for the project as part of Team “Heck Yeah” for the summer 2021 field season. In doing so, I have been presented with the opportunity to take part in its enduring legacy. It is a legacy characterized by novel discovery, distinguished leadership, and endless adventure. These particular aspects motivated me to pursue this internship. Further, the desire to seek out gray whales (Eschrichtius robustus) — a species epitomizing the ability to exhibit resilience in the face of adversity after having experienced two unusual mortality events (UME) in the past two decades and having recovered from historically low population abundances due to whaling — sparked immeasurable excitement.
Figure 1. Nadia operating the theodolite to calculate the location of a gray whale. Source: A. Dawn.
The skills we are acquiring during this field season are essential to master so that I can pursue my aspirations of becoming a marine conservation biologist. For example, we have learned how to operate a theodolite, which is a surveying tool used regularly in marine mammal research to accurately calculate the location of cetaceans and track their movements (Figure 1). We are also learning how to operate a number of other research equipment, to navigate a tandem kayak using a GPS, to process various forms of data, and to identify gray whales! I have especially enjoyed collecting prey samples and navigating our tandem kayak, as kayaking is a summer tradition for my family and the opportunity to kayak in this context is certainly the high point of this internship. The kayak is named “Robustus” after the scientific name of the gray whale: Eschrichtius robustus! (Figure 2).
Figure 2. Nadia navigating Robustus, the research kayak.
The Port Orford project aims to determine how gray whale foraging is affected by prey quantity and quality. In fact, gray whales exhibit specificity in their selection of prey on the basis of caloric content (Hildebrand 2020). I am particularly interested in the underlying implications these findings imply: the notion of conscious reasoning and decision-making by individual whales as they seek the most suitable prey for its dietary needs among other options to maximize its survivability. Are gray whales in possession of an awareness that allows them to exhibit intentional preference? Can the behavior be attributed to instinct and/or learned behavior, or to cognition comparable to human preference? These and similar questions are my motivation for studying the realm of marine mammal biology. These questions concern intelligence and evolution, which can be effectively investigated through an analysis of cetacean brain structure, as it likely has compelling relationships to their extensive behavioral abilities (Hof and Van Der Gucht 2007).
For instance, the brain of the gray whale has expanded and developed extensively over evolutionary time in response to distinct selection pressures. Evidence affirms that the behavioral challenges associated with foraging exert strong selection pressures on the evolution of their brain size and structure (Muller and Montgomery 2019)! Selection pressures associated with social cognition are also believed to have contributed to such growth (Connor et al. 1998; Marino 2002; Shultz and Dunbar 2010 ). Further, their neural organization has increased in complexity, leading to greater function and usage of the cortical portion of the brain, which is the portion responsible for higher level activity (Oelschläger and Oelschläger 2002).
Figure 3. Structure of humpback whale brain representative of baleen species used to infer about gray whales (Hof and Van Der Gucht 2007).
Though research about baleen whale brain morphology is not as pervasive as that of toothed whales (due to increased susceptibility of toothed whales to captivity given the feasibility of their capture and subsequent analysis in lab/controlled setting), studies have indicated that the brain of baleen whales share similarities to those of humans (Wade et. al 2012). In particular, similarities exist in the frontal lobe of the brain, which is responsible for the complex activities of self-awareness, reasoning, and behavior, as well as for problem-solving and motivation (Hof and Van Der Gucht 2007) (Figure 3). These findings indicate that baleen whales, including the gray whale, have the capability to exhibit intentional preference and take part in conscious decision-making in the recognition of different prey species. The mechanisms responsible for how gray whales may discern prey likely involve a number of the sensory systems, differing in respect to spatial scale (Torres 2017). Thus, gray whales likely rely on various sensory methods, such as vision, sound perception/reception, chemoreception, or an oceanographic stimulus, at differing scales to locate and discern prey. The sensory method employed is dependent on their distance from prey.
Though we cannot yet confirm whether and/or how gray whales are capable of distinguishing between prey species, what is certain, is that the gray whale is intelligent and quite similar to us. Moreover, they are representative of strength and endurance, providing lessons we can learn from and qualities we can embody. Despite the threats to the species from fishing gear entanglement, ship collisions, climate change, oil industry developments, and being historically hunted, they have remarkably persisted. Thus, we must ensure the existence of the gray whale so they too may thrive for the rest of time, with healthy lives and habitat that is rightfully theirs.
P.S. I would like to thank the GEMM Lab, Oregon State University, Shalynn Pack, Port Orford Sustainable Seafood, Port Orford Co-op, South Coast Tours, Nicki’s Knick Knacks, Leigh Torres, Lisa Hildebrand, Allison Dawn, Clara Bird, Tom Calvanese, Maddie English, Jasen White, and Damian Amerman-Smith for making the internship as special and memorable as it is/was.
References
Connor, R. C., Mann, J., Tyack, P. L., and Whitehead, H. (1998). Social evolution in toothed whales. Trends in Ecology and Evolution, 13(6): 228– 232. doi: https://doi.org/10.1016/S0169‐5347(98)01326‐3
Hildebrand, L. (2020). Tonight’s specials include mysids, amphipods, and more: an examination of the zooplankton prey of Oregon gray whales and its impact on foraging choices and prey selection. Master’s thesis, Oregon State University.
Hof, P.R., and Van Der Gucht, E. (2007). Structure of the cerebral cortex of the humpback whale, Megaptera novaengliae(Cetacea, Mysticeti, Balaenopteridae). The Anatomical Record 290:1-31 doi: 10.1002/ar.a.20407
Marino, L. (2002). Convergence of complex cognitive abilities in cetaceans and primates. Brain, Behavior, and Evolution59: 21–32. doi: https://doi. org/10.1159/000063731
Oelschläger, H.A., and Oelschläger, J.S. (2002). Brains. In: Perrin WF, Wu¨ rsig B, Thewissen JGM, editors. Encyclopedia of marine mammals. San Diego, CA: Academic Press. p 133–158.
Shultz, S., & Dunbar, R. (2010). Encephalization is not a universal macroevolutionary phenomenon in mammals but is associated with sociality. Proceedings of the National Academy of Sciences of the United States of America 107(50): 21582–21586. doi: https://doi.org/10.1073/ pnas.1005246107
Torres, L.G. (2017). A sense of scale: foraging cetaceans’ use of scale-dependent multimodal sensory systems. Marine Mammal Science 33: 1170-1193. doi: 10.1111/mms.12426
Wade, P.R., Reeves, R.R., and Mesnick, S.L. (2012). Social and behavioral factors in cetacean responses to overexploitation: are odontocetes less “resilient” than mysticetes?. The Journal of Marine Biology 2012: 1-15. doi:10.1155/2012/567276
Predators have high energetic requirements that must be met to ensure reproductive success and population viability. For baleen whales, this task is particularly challenging since their foraging seasons are typically limited to short temporal windows during summer months when they migrate to productive high latitude environments. Foraging success is a balancing act whereby baleen whales must maximize the amount of energy they intake, while minimizing the amount of energy they expend to obtain food. Maximization of energy intake can be achieved by targeting the most beneficial prey. How beneficial a particular prey type (or prey patch) is can depend on a number of factors such as abundance, density, quality, size, and availability. Determining why baleen whales target particular prey types or patches is an important factor to enhance our understanding of their ecology and can ultimately aid in informing their management and conservation.
The GEMM Lab has several research projects in Newport and Port Orford, Oregon, on the Pacific Coast Feeding Group (PCFG), which is a sub-group of gray whales from the Eastern North Pacific (ENP) population. While ENP gray whales feed in the Bering, Chukchi, and Beaufort Seas (Arctic) in the summer months, the PCFG utilizes the range from northern California, USA to northern British Columbia, Canada. Our work to date has revealed a number of new findings about the PCFG including that they successfully gain weight during the summer on Oregon foraging grounds (Soledade Lemos et al. 2020). Furthermore, females that consistently use the PCFG range as their foraging grounds have successfully reproduced and given birth to calves (Calambokidis & Perez 2017). Yet, the abundance of the PCFG (~250 individuals; Calambokidis et al. 2017) is two orders of magnitude smaller than the ENP population (~20,000; Stewart & Weller 2021). So, why do more gray whales not use the PCFG range as their foraging grounds when it provides a shorter migration while also allowing whales to meet their high energetic requirements and ensure reproductive success? There are several hypotheses regarding this ecological mystery including that prey abundance, density, quality, and/or availability are higher in the Arctic than in the PCFG range, thus justifying the much larger number of gray whales that migrate further north for the summer feeding season.
Figure 1. Locations of prey samples collected with a light trap (open circles) or opportunistic collections of surface swarms of crab larvae (black triangles) in Newport, along the Oregon coast in the Pacific Northwest coast of the United States.
Our recent paper in Frontiers in Marine Science addressed the hypothesis that prey quality in the Arctic is higher than that of PCFG prey. To test this hypothesis, we first determined the quality (energetic value) of nearshore Oregon zooplankton species that PCFG gray whales are assumed to feed on (based on observations of fine-scale spatial and temporal overlap of foraging gray whales and sampled zooplankton). We obtained prey samples from nearshore reefs along the Oregon coast (Figure 1) as part of the GRANITE project using a light trap, which is a modified water jug with a weight and two floats attached to it, allowing the trap to sit approximately 1 meter above the seafloor. The trap contained a light which attracted zooplankton and effectively captured epibenthic prey of gray whales. Traps were left to soak overnight in locations where gray whales had been observed feeding extensively and collected the following morning. After identifying each specimen to species level and sorting them into reproductive stages, we used a bomb calorimeter to determine the caloric content of each species by month, year, and reproductive stage. We then compared these values to the literature-derived caloric value of the predominant benthic amphipod species that ENP gray whales feed on in the Arctic. These comparisons allowed us to extrapolate the caloric values gained from each prey type to estimated energetic requirements of pregnant and lactating female gray whales (Villegas-Amtmann et al. 2017).
Figure 2. Median caloric content and interquartile ranges by (A) species, (B) reproductive stage, and (C) month. Sizes of the zooplankton images are scaled at actual ratios relative to one another.
So, what did we find? Our sampling along the Oregon coast revealed six predominant zooplankton species: two mysid shrimp (Neomysis rayii, Holmesimysis sculpta), two amphipods (Atylus tridens, Polycheria osborni), and two types of crab larvae (Dungeness crab megalopae, porcelain crab larvae). These six Oregon prey species showed significant differences in their caloric values, with N. rayii and Dungeness crab megalopae having significantly higher calories per gram than the other prey species (Figure 2), though Dungeness crab megalopae stood out as the caloric gold mines for feeding gray whales in the PCFG range. Furthermore, month and reproductive stage also influenced the caloric content of some prey species, with gravid (aka pregnant) female mysid shrimp significantly increasing in calories throughout the summer (Figure 3).
Figure 3. Caloric content of different reproductive stages as a function of day of year (DOY; ranging from June to October) for the mysids Holmesimysis sculpta and Neomysis rayii, and the amphipod Atylus tridens. A. tridens is only represented on one panel due to small sample size of this species for the empty brood pouch and gravid reproductive stages. Asterisks indicate significant regressions (p<0.05).
The comparison of our Oregon prey caloric values to the predominant Arctic amphipod (Ampelisca macrocephala) proved our hypothesis wrong: Arctic amphipods do not have higher caloric value than Oregon prey, which would have help to explain why many more gray whales feed in the Arctic. We found that two Oregon prey species (N. rayii and Dungeness crab megalopae) have higher caloric values than A. macrocephala. If we translate the caloric contents of these prey to gray whale energetic needs, these differences mean that lactating and pregnant gray whales feeding in the PCFG area would need between 0.7-1.03 and 0.22-0.33 metric tons of prey less per day if they fed on Dungeness crab megalopae or N. rayii, respectively, than a whale feeding on Arctic A. macrocephala (Figure 4).
Figure 4. Daily prey requirements (A: metric tons; B: number of individuals) needed by pregnant and lactating female gray whales to meet their energetic requirements on the foraging ground. Energetic requirement estimates obtained from Villegas-Amtmann et al. (2017). Note the logarithmic scale of y-axis in panel (B).
If quality were the only prey metric that gray whales used to evaluate which food to eat, then it would make very little sense for so many gray whales to migrate to the Arctic when there are prey types of equal and greater quality available to them in the PCFG range. However, quality is not the only metric that influences gray whale foraging decisions. We therefore posit that the abundance, density, and availability of benthic amphipods in the Arctic are higher than the prey species found in the PCFG range. In fact, knowledge of the pulsed reproductive cycle of Dungeness and porcelain crabs allows us to conclude that the larvae of these two species are only available for a few weeks in the late spring and early summer on the Oregon coast. While mysid shrimp, such as N. rayii, are continuously available in the PCFG range throughout the summer, they may occur in less dense and more patchy aggregations than Arctic benthic amphipods. However, current estimates of prey density and abundance for either region are not available, and we do not have data on the energetic costs of the different foraging strategies. While there are still several unknowns, we have documented that higher prey quality in the Arctic is not the reason for the difference in gray whale foraging ground use in the eastern North Pacific.
References
Calambokidis, J., & Perez, A. 2017. Sightings and follow-up of mothers and calves in the PCFG and implications for internal recruitment. IWC Report SC/A17/GW/04 for the Workshop on the Status of North Pacific Gray Whales (La Jolla: IWC).
Calambokidis, J., Laake, J., & Perez, A. 2017. Updated analysis of abundance and population structure of seasonal gray whales in the Pacific Northwest, 1996-2015. IWC Report SC/A17/GW/05 for the Workshop on the Status of North Pacific Gray Whales (La Jolla: IWC).
Soledade Lemos, L., Burnett, J. D., Chandler, T. E., Sumich, J. L., & Torres, L. G. 2020. Intra- and inter-annual variation in gray whale body condition on a foraging ground. Ecosphere 11(4):e03094.
Stewart, J. D., & Weller, D. W. 2021. Abundance of eastern North Pacific gray whales 2019/2020. Department of Commerce, NOAA Technical Memorandum NMFS-SWFSC-639. United States: NOAA. doi:10.25923/bmam-pe91.
Villegas-Amtmann, S., Schwarz, L. K., Gailey, G., Sychenko, O., & Costa, D. P. 2017. East or west: the energetic cost of being a gray whale and the consequence of losing energy to disturbance. Endangered Species Research 34:167-183.
Since early May, much of the GEMM Lab has been consumed by the GRANITE project, which stands for Gray whale Response to Ambient Noise Informed by Technology and Ecology. Two weeks ago, PhD student Clara Bird discussed our field work preparations, and since May 20th we have conducted five successful days of field work (and one unsuccessful day due to fog). If you are now expecting a blog about the data we have collected so far and whales we encountered, I am sorry to disappoint you. Rather, I want to take a big step back and provide the context of the GRANITE project as a whole, explain why this project and data collection is so important, and discuss what it is that we hope to achieve with our ever-growing, multidisciplinary dataset and team.
We use the Pacific Coast Feeding Group (PCFG) of gray whales that forage off the Oregon coast as our study system to better understand the ecological and physiological response of baleen whales to multiple stressors. Our field methodology includes replicate physiological and ecological sampling of this accessible baleen whale population with synoptic measurement of multiple types of stressors. We collect fecal samples for hormone analysis, conduct drone overflights of whales to collect body condition and behavioral data, record the ambient soundscape through deployment of two hydrophones, and conduct whale photo-identification to link all data streams to each individual whale of known sex, estimated age, and reproductive status. We resample these data from multiple individuals within and between summer foraging seasons, while exposed to different potential stressors occurring at different intensities and temporal periods and durations. The hydrophones are strategically placed with one in a heavily boat-trafficked (and therefore noisy) area close to the Port of Newport, while the second is located in a relatively calm (and therefore quieter) spot near the Otter Rock Marine Reserve (Fig. 1). These hydrophones provide us with information about both natural (e.g. killer whales, wind, waves) and anthropogenic (e.g. boat traffic, seismic survey, marine construction associated with PacWave wave energy facility development) noise that may affect gray whales. During sightings with whales, we also drop GoPro cameras and sample for prey to better understand the habitats where whales forage and what they might be consuming.
Figure 1. Map of GRANITE study area from Seal Rock to Lincoln City with gray whale sightings (yellow circles) and and fecal samples collected (red triangles) from the 2020 field season. Green stars represent the two hydrophone locations. Source: L. Torres.
GEMM Lab PI Dr. Leigh Torres initiated this research project in 2015 and established partnerships with acoustician Dr. Joe Haxel and (then) PhD student Dr. Leila Lemos. Since then, the team working on this project has grown considerably to provide expertise in the various disciplines that the project integrates. Leigh is currently joined at the GRANITE helm by 4 co-PIs: Dr. Haxel, endocrinologist Dr. Kathleen Hunt, biological statistician Dr. Leslie New, and physiologist Dr. Loren Buck. Drs. Alejandro Fernandez Ajo, KC Bierlich and Enrico Pirotta are postdoctoral scholars who are working on the endocrinology, photogrammetry, and biostatistical modelling components, respectively. Finally, Clara and myself are partially funded through this project for our PhD research, with Clara focusing on the links between behavior, body condition, individualization, and habitat, while I am tackling questions about the recruitment and site fidelity of the PCFG (more about these topics below).
Faculty Research Assistant Todd Chandler supervises PhD student Clara Bird during her maiden drone flight over a whale. Source: L. Torres.
The ultimate goal of this project is to use the PCFG as a case study to quantify baleen whale physiological response to different stressors and model the subsequent impacts on the population by implementing our long-term, replicate dataset into a framework called Population consequences of disturbance (PCoD; Fig. 2). PCoD is built upon the underlying concept that changes in behavior and/or physiology caused by disturbance (i.e. noise) affect the fitness of individuals by impacting their health and vital rates, such as survival, reproductive success, and growth rate (Pirotta et al. 2018). These impacts at the individual level may (or may not) affect the population as a whole, depending on what proportion of individuals in the population are affected by the disturbance and the intensity of the disturbance effect on each individual. The PCoD framework requires quantification of four stages: a) the physiological and/or behavioral changes that occur as a result of exposure to a stressor (i.e. noise), b) the acute effects of these physiological and/or behavioral responses on individual vital rates, and their chronic effects via individual health, c) the way in which changes in health may affect the vital rates of individuals, and d) how changes in individual vital rates may affect population dynamics (Fig. 2; Pirotta et al. 2018). While four stages may not sound like a lot, the amount and longevity of data needed to quantify each stage is immense.
Figure 2. Conceptual framework of the population consequences of disturbance (PCoD). Letters (A-D) represent the four stages that require quantification in order for PCoD to be implemented. Each colored box represents external (ecological drivers, stressors) and internal (physiology, health, vital rates, behavior) factors that can change over time that are measured for each individual whale (dashed grey boundary line). The effects are then integrated across all individuals in the population to project their effects on the population’s dynamics. Figure and caption adapted from Pirotta et al. 2018.
The ability to detect a change in behavior or physiology often requires an understanding of what is “normal” for an individual, which we commonly refer to as a baseline. The best way to establish a baseline is to collect comprehensive data over a long time period. With our data collection efforts since 2015 of fecal samples, drone flights and photo identification, we have established useful baselines of behavioral and physiological data for PCFG gray whales. These baselines are particularly impressive since it is typically difficult to collect repeated measurements of hormones and body condition from the same individual baleen whale across multiple years. These repeated measurements are important because, like all mammals, hormones and body condition vary across life history phases (i.e., with pregnancy, injury, or age class) and across time (i.e., good or bad foraging conditions). To achieve these repeated measurements, GRANITE exploits the high degree of intra- and inter-annual site fidelity of the PCFG, their accessibility for study due to their affinity for nearshore habitat use, and the long-term sighting history of many whales that provides sex and approximate age information. Our work to-date has already established a few important baselines. We now know that the body condition of PCFG gray whales increases throughout a foraging season and can fluctuate considerably between years (Soledade Lemos et al. 2020). Furthermore, there are significant differences in body condition by reproductive state, with calves and pregnant females displaying higher body conditions (Soledade Lemos et al. 2020). Our dataset has also allowed us to validate and quantify fecal steroid and thyroid hormone metabolite concentrations, providing us with putative thresholds to identify a stressed vs. not stressed whale based on its hormone levels (Lemos et al. 2020).
PhD student Lisa Hildebrand and GRANITE co-PI Dr. Kathleen Hunt collecting a fecal sample. Source: L. Torres.
We continue to collect data to improve our understanding of baseline PCFG physiology and behavior, and to detect changes in their behavior and physiology due to disturbance events. All these data will be incorporated into a PCoD framework to scale from individual to population level understanding of impacts. However, more data is not the only thing we need to quantify each of the PCoD stages. The implementation of the PCoD framework also depends on understanding several aspects of the PCFG’s population dynamics. Specifically, we need to know whether recruitment to the PCFG population occurs internally (calves born from “PCFG mothers” return to the PCFG) or externally (immigrants from the larger Eastern North Pacific gray whale population joining the PCFG as adults). The degree of internal or external recruitment to the PCFG population should be included in the PCoD model as a parameter, as it will influence how much individual level disturbance effects impact the overall health and viability of the population. Furthermore, knowing residency times and home ranges of whales within the PCFG is essential to understand exposure durations to disturbance events.
To assess both recruitment and residency patterns of the PCFG, I am undertaking a large photo-identification effort, which includes compiling sightings and photo data across many years, regions, and collaborators. Through this effort we aim to identify calves and their return rate to the population, the rate of new adult recruits to the population, and the spatial residency of individuals in our study system. Although photo-id is a basic, commonplace method in marine mammal science, its role is critical to tracking individuals over time to understand population dynamics (in a non-invasive manner, no less). A large portion of my PhD research will focus on the tedious yet rewarding task of photo-id data management and matching in order to address these pressing knowledge gaps on PCFG population dynamics needed to implement the PCoD model that is an ultimate goal of GRANITE. I am just beginning this journey and have already pinpointed many analytical and logistical hurdles that I need to overcome. I do not anticipate an easy path to addressing these questions, but I am extremely eager to dig into the data, reveal the patterns, and integrate the findings into our rock-solid GRANITE project.
Funding for the GRANITE project comes from the Office of Naval Research, the Department of Energy, Oregon Sea Grant, the NOAA/NMFS Ocean Acoustics Program, and the OSU Marine Mammal Institute.
References
Lemos, L.S., Olsen, A., Smith, A., Chandler, T.E., Larson, S., Hunt, K., and L.G. Torres. 2020. Assessment of fecal steroid and thyroid hormone metabolites in eastern North Pacific gray whales. Conservation Physiology 8:coaa110.
Pirotta, E., Booth, C.G., Costa, D.P., Fleishman, E., Kraus, S.D., Lusseau, D., Moretti, D., New, L.F., Schick, R.S., Schwarz, L.K., Simmons, S.E., Thomas, L., Tyack, P.L., Weise, M.J., Wells, R.S., and J. Harwood. 2018. Understanding the population consequences of disturbance. Ecology and Evolution 8(19):9934-9946.
Soledade Lemos, L., Burnett, J.D., Chandler, T.E., Sumich, J.L., and L.G. Torres. 2020. Intra- and inter-annual variation in gray whale body condition on a foraging ground. Ecosphere 11(4):e03094.
Clara Bird, PhD Student, OSU Department of Fisheries, Wildlife, and Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
When I thought about what doing fieldwork would be like, before having done it myself, I imagined that it would be a challenging, but rewarding and fun experience (which it is). However, I underestimated both ends of the spectrum. I simultaneously did not expect just how hard it would be and could not imagine the thrill of working so close to whales in a beautiful place. One part that I really did not consider was the pre-season phase. Before we actually get out on the boats, we spend months preparing for the work. This prep work involves buying gear, revising and developing protocols, hiring new people, equipment maintenance and testing, and training new skills. Regardless of how many successful seasons came before a project, there are always new tasks and challenges in the preparation phase.
For example, as the GEMM Lab GRANITE project team geared up for its seventh field season, we had a few new components to prepare for. Just to remind you, the GRANITE (Gray whale Response to Ambient Noise Informed by Technology and Ecology) project’s field season typically takes place from June to mid-October of each year. Throughout this time period the field team goes out on a small RHIB (rigid hull inflatable boat), whenever the weather is good enough, to collect photo-ID data, fecal samples, and drone imagery of the Pacific Coast Feeding Group (PCFG) gray whales foraging near Newport, OR, USA. We use the data to assess the health, ecology and population dynamics of these whales, with our ultimate goal being to understand the effect of ambient noise on the population. As previous blogs have described, a typical field day involves long hours on the water looking for whales and collecting data. This year, one of our exciting new updates is that we are going out on two boats for the first part of the field season and starting our season 10 days early (our first day was May 20th). These updates are happening because a National Science Foundation funded seismic survey is being conducted within our study area starting in June. The aim of this survey is to assess geophysical structures but provides us with an opportunity to assess the effect of seismic noise on our study group by collecting data before, during, and after the survey. So, we started our season early in order to capture the “before seismic survey” data and we are using a two-boat approach to maximize our data collection ability.
While this is a cool opportunistic project, implementing the two-boat approach came with a new set of challenges. We had to find a second boat to use, buy a new set of gear for the second boat, figure out the best way to set up our gear on a boat we had not used before, and update our data processing protocols to include data collected from two boats on the same day. Using two boats also means that everyone on the core field team works every day. This core team includes Leigh (lab director/fearless leader), Todd (research assistant), Lisa (PhD student), Ale (new post-doc), and me (Clara, PhD student). Leigh and Todd are our experts in boat driving and working with whales, Todd is our experienced drone pilot, I am our newly certified drone pilot, and Lisa, Ale, and myself are boat drivers. Something I am particularly excited about this season is that Lisa, Ale, and I all have at least one field season under our belts, which means that we get to become more involved in the process. We are learning how to trailer and drive the boats, fly the drones, and handling more of the post-field work data processing. We are becoming more involved in every step of a field day from start to finish, and while it means taking on more responsibility, it feels really exciting. Throughout most of graduate school, we grow as researchers as we develop our analytical and writing skills. But it’s just as valuable to build our skillset for field work. The ocean conditions were not ideal on the first day of the field season, so we spent our first day practicing our field skills.
For our “dry run” of a field day, we went through the process of a typical day, which mostly involved a lot of learning from Leigh and Todd. Lisa practiced her trailering and launching of the boat (figure 1), Ale and Lisa practiced driving the boat, and I practiced flying the drone (figure 2). Even though we never left the bay or saw any whales, I thoroughly enjoyed our dry run. It was useful to run through our routine, without rushing, to get all the kinks out, and it also felt wonderful to be learning in a supportive environment. Practicing new skills is stressful to say the least, especially when there is expensive equipment involved, and no one wants to mess up when they’re being watched. But our group was full of support and appreciation for the challenges of learning. We cheered for successful boat launchings and dockings, and drone landings. I left that day feeling good about practicing and improving my drone piloting skills, full of gratitude for our team and excited for the season ahead.
Figure 1. Lisa (driving the truck) launching the boat.
Figure 2. Clara (seated, wearing a black jacket) landing the drone in Ale’s hands.
All the diligent prep work paid off on Saturday with a great first day (figure 3). We conducted five GoPro drops (figure 4), collected seven fecal samples from four different whales (figure 5), and flew four drone flights over three individuals including our star from last season, Sole. Combined, we collected two trifectas (photo-ID images, fecal samples, and drone footage)! Our goal is to get as many trifectas as possible because we use them to study the relationship between the drone data (body condition and behavior) and the fecal sample data (hormones). We were all exhausted after 10 hours on the water, but we were all very excited to kick-start our field season with a great day.
Figure 3. Lisa on the bow pulpit during our first sighting of the day.
Figure 4. Lisa doing a GoPro drop, she’s lowering the GoPro into the water using the line in her hands.
Figure 5. Clara and Ale collecting a fecal sample.
On Sunday, just one boat went out to collect more data from Sole after a rainy morning and I successfully flew over her from launching to landing! We have a long season ahead, but I am excited to learn and see what data we collect. Stay tuned for more updates from team GRANITE as our season progresses!
¹PhD Student, OSU Department of Fisheries, Wildlife, and Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
²Postdoctoral Scholar, OSU Department of Fisheries, Wildlife, and Conservation Sciences, Cooperative Institute for Marine Resources Studies, Hatfield Marine Science Center
Many aspects of studying cetacean ecology, behavior, population dynamics, health, and sociality depend on being able to see and/or sample cetaceans when they come to the surface. While this research is not necessarily easy given that cetaceans spend the majority of their time underwater out of human sight, it is definitely feasible, as evidenced by decades of cetacean research. However, in order for researchers to observe cetaceans at the surface they need to get out to sea, and this boat-based effort can realistically only be done in good ocean conditions. Any sea-going individual likely uses the Beaufort sea state (BSS) scale as a measure of ocean conditions. For a full breakdown and excellent explanation of what the BSS is, check out our beloved Alexa’s blog; but for the purposes of this blog all you really need to know is that the smaller the BSS (which starts at 0), the calmer the ocean, and the higher the BSS, the rougher & stormier the ocean. There are two main reasons for conducting cetacean research in low BSS: 1) above a certain threshold (usually BSS 4) it becomes difficult to reliably spot and recognize cetaceans at the surface, thus compromising good data collection, and/or 2) to ensure safety and comfort of the research team.
So, when the BSS gets too high, us humans usually do not go out to sea to study cetaceans, which means that the cetaceans, for the most part, go unobserved. So, many questions arise about what cetaceans are doing during these rough ocean conditions. What does an increased BSS mean for them? Are they unfazed by big waves and strong winds, or are they affected by the weather and take longer dives or seek out fairer seas? A conversation among friends sparked our curiosity of what cetaceans do in stormy conditions and inspired us to collaborate on this blog. Here, we report on what is and is not known about cetaceans in storms, and discuss some ideas about how best to quantify the effects of rough sea conditions on cetaceans.
Slide the arrows to compare sea conditions (BSS 1 [left] vs BSS 6 [right]) experienced by Alexa, the GEMM Lab marine mammal observer on the May 2019 Northern California Current cruise onboard NOAA ship Bell M. Shimada. Source: A. Kownacki/GEMM Lab.
A literature search of cetaceans during storms did not generate many results, which was not surprising to us given the above reasons about researchers not being able to survey in rough sea conditions. However, we did find a couple of interesting studies about cetacean behavior and distribution after storms.
Changes in foraging behavior
Autumnal storms in Maryland, USA resulted in less frequent and shorter encounters of bottlenose dolphins in the US Mid-Atlantic Bight. However, dolphins spent a significantly higher percentage of their encounters feeding after storms than they did before or during them (Fandel et al. 2020). Similarly, bottlenose dolphins in Mississippi Sound displayed an approximately 15% increase in foraging activity for up to 2 years following Hurricane Katrina (Smith et al. 2013). These changes in foraging behavior are attributed to shifts in distributions and behavior of dolphin prey species as a result of altered environmental conditions (primarily sea surface temperature and salinity) following the hurricanes.
Out-of-habitat events and strandings
An out-of-habitat event occurs when an animal is displaced out of its typical habitat. Seven of these events were reported following Hurricane Rita, which hit the southwest Louisiana coast in 2005, with bottlenose dolphins found in flooded roadside ditches, canals, shallow flooded fields, and a natural creek area (Rosel & Watts 2008). These locations ranged from 2.5 to 11 km inland from the coast of the Gulf of Mexico, where these dolphins were displaced from. It is believed that the animals were carried inland on the storm surge that accompanied Hurricane Rita and were left stranded in areas that held water the longest once it started receding (Rosel & Watts 2008).
One of the roadside ditches where a bottlenose dolphin was trapped in Louisiana following Hurricane Rita. Taken from Rosel & Watts (2008).
There have been two mass strandings of pygmy killer whales that are believed to have been a result of hurricanes. In 1995, five pygmy killer whales stranded (three of which died, while two were successfully refloated) in the British Virgin Islands a day after Hurricane Marilyn (Mignucci-Giannoni et al. 1999). In 2006, six pygmy killer whales (five of which died) stranded in New Caledonia during and after Hurricane Jim (Clua et al. 2014). Both studies hypothesize that increased energetic costs, as a result of attempting to evade the hurricanes, coupled with animals becoming disoriented and ending up in shallow waters, is what caused them to strand.
While these studies reveal post-storm effects on cetaceans, we still do not know exactly how these individuals behaved during the storms. Did they attempt longer dives to stay away from the rough conditions at the surface, thus becoming disoriented? Or were they behaving normally (i.e. foraging, travelling) and were simply “pushed” into waters that they did not intend to go into? Given that very stormy sea conditions do not allow for visual, boat-based surveys, we need to employ different technologies to study cetacean behavior and distribution during storms.
Passive acoustic monitoring (PAM) is a great tool that can monitor ocean environments for us when the seas are too stormy. Using fixed or mobile platforms, underwater PAM listening devices (hydrophone and data storage) can record sounds in the ocean for us to listen and analyze from shore. With PAM we are able to track the vocalizations of marine mammals as well as other sounds in the environment, such as waves crashing and rain. Anecdotally, we have spent many days at sea in conditions that were too rough for visual observations, but we could safely use our PAM tools to detect cetaceans. So, just because the seas may be too rough to see cetaceans, this fact does not mean that we cannot observe them – we just need to listen instead of look.
There are many tools that can be used to record underwater sounds, including passive acoustic monitoring (PAM; shown in orange), real-time acoustic data collection (green), and active acoustics (blue.) Source: NOAA Fisheries.
A number of studies have investigated whether whales change their vocalization behavior differently in response to changing ambient sound conditions (for example: Dunlop et al. 2010; Fournet et al. 2018). While research on ocean sound levels is often focused on the impact of human-generated or anthropogenic noise, there are also natural, abiotic sound sources (e.g. wind, rain, ice) that can elevate ambient sound levels. One potential animal response to elevated ambient sound levels is to vocalize at a higher intensity, called the Lombard (or cocktail party) effect. This phenomenon is common for us humans – have you ever been at a party and at some point you realize that you are shouting to someone in order to be heard above the noise of the room? That’s the Lombard effect! Humpback whales in Glacier Bay National Park, Alaska, exhibited the Lombard effect in response to both natural and man-made sounds, but the probability of calling was lower when vessels were present compared to times with only natural sounds (Fournet et al. 2018). It is also possible that whales may vocalize at different frequencies, times, or for shorter durations when the ocean becomes louder, which we can easily track with PAM. Unfortunately, PAM is limited to what we are able to hear, so if we do not hear whales we cannot determine if this result is because their vocalizations are masked by higher intensity sounds, if they stopped vocalizing, or if they left the listening area.
Animal-borne tags are another kind of autonomous observation tool that could help us understand cetacean behavior and distribution in storms. Admittedly, the logistics of applying tags before an imminent storm are probably complex. However, the development of medium-duration archival tags may provide a good trade-off between deploying tags long enough before a storm begins, thus providing safe working conditions for the research team, while minimizing potential physical impacts to the animals (Szesciorka et al. 2016). There are currently no published tag studies that document cetacean behavior during storms, but a study of a gray-headed albatross, fitted with a satellite transmitter, that successfully foraged during an Antarctic storm (Catry et al. 2004) shows the promise of using animal-borne tags to answer these questions.
As with many questions about animal behavior, our best option is to combine all of our research tools to piece together evidence about what might be going on in the deep, dark, stormy ocean. Simultaneously collecting acoustic and movement & behavior data through PAM and animal-borne tags, respectively, could allow us to determine how cetaceans behave during storms. While we are probably not poised to tackle these questions right now, perhaps another curious graduate student can take it on for their own PhD research…
References
Catry, P., Phillips, R.A., and J.P. Croxall. Sustained fast travel by a gray-headed albatross (Thalassarchie chrysostoma) riding an Antarctic storm. The Auk 121(4):1208-1213.
Clua, E.E., Manire, C.A., and C. Garrigue. 2014. Biological data of pygmy killer whales (Feresa attenuata) from a mass stranding in New Caledonia (South Pacific) associated with Hurricane Jim in 2006. Aquatic Mammals 40(2):162-172.
Dunlop, R.A., Cato, D.H., and M.J. Noad. 2010. Your attention please: increasing ambient noise levels elicits a change in communication behaviour in humpback whales (Megaptera novaeangliae). Proceedings of the Royal Society B 277(1693):doi.org/10.1098/rspb.2009.2319.
Fandel, A.D., Garrod, A., Hoover, A.L., Wingfield, J.E., Lyubchich, V., Secor, D.H., Hodge, K.B., Rice, A.N., and H. Bailey. 2020. Effects of intense storm events on dolphin occurrence and foraging behavior. Scientific Reports 10:19247.
Fournet, M.E.H., Matthews, L.P., Gabriele, C.M., Haver, S., Mellinger, D.K., and H. Klinck. 2018. Humpback whales Megaptera novaeangliae alter calling behavior in response to natural sounds and vessel noise. Marine Ecology Progress Series 607:251-268.
Mignucci-Giannoni, A.A., Toyos-González, G. M., Pérez-Padilla, J., Rodríguez-López, M. A., and J. Overing. 1999. Mass stranding of pygmy killer whales (Feresa attenuata) in the British Virgin Islands. Journal of the Marine Biological Association of the United Kingdom 80:759-760.
Rosel, P.E., and H. Watts. 2008. Hurricane impacts on bottlenose dolphins in the northern Gulf of Mexico. Gulf of Mexico Science 25:7.
Smith, C.E., Hurley, B.J., Toms, C.N., Mackey, A.D., Solangi, M., and S.A. Kuczaj II. 2013. Hurricane impacts on the foraging patterns of bottlenose dolphins Tursiops truncatus in Mississippi Sound. Marine Ecology Progress Series 487:231-244.
Szesciorka, A.R., Calambokidis, J., and J.T. Harvey. 2016. Testing tag attachments to increase the attachment duration of archival tags on baleen whales. Animal Biotelemetry 4:18.
Last week marked the one year anniversary of the pandemic reality we have all been living. It has been an extremely challenging year, with everyone experiencing different kinds of difficulties and hurdles. One challenge that likely unites the majority of us is having to forego seeing our loved ones. For me personally, this is the longest time I have not seen my family (445 days and counting) and I know I am not alone in this situation. My homesickness started a train of thought about cetacean parental care and inspired me to write a blog about this topic. As you can see from the title, this post focuses on maternal care, rather than parental care. This bias isn’t due to my lack of research on this topic or active exclusion, but rather because there are currently no known cetacean species where paternal participation in offspring production and development extends beyond copulation (Rendell et al. 2019). Thus, this blog is all about the role of mothers in the lives of cetacean calves.
Like humans, cetacean mothers invest a lot of energy into their offspring. Most species have a gestation period of 10 or more months (Oftedal 1997). For baleen whale females in particular, pregnancy is not an easy feat given that they only feed during summer feeding seasons. They must therefore acquire all of the energy they will need for two migrations, birth, and (almost) complete lactation, before they will have access to food on feeding grounds again. For pregnant gray whales, a mere 4% loss in average energy intake on the foraging grounds will prevent her from successfully producing and/or weaning a calf (Villegas-Amtmann et al. 2015), demonstrating how crucial the foraging season is for a pregnant baleen whale. Once a calf is born, lactation ensues, ranging in length between approximately 6-8 months for most baleen whale species to upwards of one or two years in odontocetes (Oftedal 1997). The very short lactation period in baleen whales is offset by the large volume (for blue whales, up to 220 kg per day) and high fat percentage (30-50%) of milk that mothers provide for their calves (Oftedal 1997). In contrast, odontocetes (or toothed whales) have a more prolonged period of lactation with less fatty milk (10-30%). This discrepancy in lactation period lengths is in part because odontocete species do not undertake long migrations, which allows females to feed year-round and therefore allocate energy to nursing young for a longer time.
Blue whale calf nursing in New Zealand in 2016. Footage captured via unmanned aerial system (UAS; drone) piloted by Todd Chandler for GEMM Lab’s OBSIDIAN project. Source: GEMM Lab.
Aside from the energetically costly task of lactation, cetacean mothers must also assist their calves as they learn to swim. Echelon swimming is a common position of mother-calf pairs whereby the calf is in very close proximity to its mother’s mid-lateral flank and provides calves with hydrodynamic benefits. Studies in bottlenose dolphins have shown that swimming in echelon results in a 24% reduction in mean maximum swim speeds and a 13% decrease in distance per stroke (Noren 2008) for mothers, while concurrently increasing average swim speeds and distance per stroke of calves by 28% and 19%, respectively (Noren et al. 2007). While these studies have only been conducted in odontocete species, echelon swimming is also observed in baleen whales (Smultea et al. 2017), indicating that baleen whale females may experience the same reductions in swimming efficiency. Furthermore, mothers will forgo sleep in the first days after birth (killer whales & bottlenose dolphins; Lyamin et al. 2005) and/or shorten their dive foraging times to accommodate calf diving ability (bottlenose dolphins [Miketa et al. 2018] & belugas [Heide-Jørgensen et al. 2001]). Females must endure these losses in foraging opportunities and decreased swimming efficiency when they are at their most nutritionally stressed to ensure the well-being and success of their offspring.
It is at the time of weaning (when a calf becomes independent), that we start to see differences in the maternal role between baleen and toothed whale mothers. Odontocetes have much stronger sociality than baleen whales causing offspring to stay with their mothers for much longer periods. Among the largest toothed whales, such as killer and sperm whales, offspring stay with their mothers in stable matrilineal units for often a lifetime. Among the smaller toothed whales, such as bottlenose dolphins, maternal kin maintain strong bonds in dynamic fission-fusion societies. In contrast, post-weaning maternal care in baleen whales is limited, with the mother-calf pair typically separating soon after the calf is weaned (Rendell et al. 2019).
Conceptual diagram depicting where baleen (Mysticeti) and toothed (Odontoceti) whales fall on the continuum of low to high social structure and matrilineal kinship structure. The networks at the top depict long-term datasets of photo-identified individuals (red nodes = females, blue nodes = males, yellow nodes = calves) with thickness of connecting lines representing strength of association between individuals. Figure and caption [adapted] from Rendell et al. 2019.
The long-term impact of social bonds in odontocetes is evident through examples of vertically transmitted behaviors (from mother to calf) in a number of species. For example, the use of three unique foraging tactics (sponge carrying, rooster-tail foraging, and mill foraging) by bottlenose dolphin calves in Shark Bay, Australia, was only significantly explained by maternal use of these tactics (Sargeant & Mann 2009). In Brazil, individuals of four bottlenose dolphin populations along the coast cooperatively forage with artisanal fishermen, which involves specialized and coordinated behaviors from both species. This cooperative foraging tactic among dolphins is primarily maintained across generations via social learning from mothers to calves (Simões-Lopeset al. 2016). The risky tactic of intentional stranding by killer whales on beaches to capture elephant seal pups requires a high degree of skill and high parental investment to reduce the associated risk of stranding (Guinet & Bouvier 1995).
Evidence for vertical transmission of specialized foraging tactics in baleen whales currently does not exist. Bubble-net feeding is a specialized tactic employed by humpback whales in three oceanic regions where multiple individuals work together to herd and trap prey (Wiley et al. 2011). However, it remains unknown whether this behavior is vertically transmitted. Simultaneous video tags from a mother-calf humpback whale pair in the Western Antarctic Peninsula documented synchrony in dives, with the calf’s track lagging behind the mother’s by 4.5 seconds, suggesting that the calf was following its mother (Tyson et al. 2012). Synchronous diving likely allows calves to observe their mothers and practice their diving, and could offer a pathway for them to mimic foraging behaviors and tactics displayed by mothers.
While there currently may not be evidence for vertical transmission of specialized foraging tactics among the baleen whales, there is documentation of matrilineal fidelity to both foraging (Weinrich 1998, Barendse et al. 2013, Burnham & Duffus 2020) and breeding grounds (Carroll et al. 2015). Matrilineal site fidelity to foraging grounds is not exclusive to baleen whales and has also been documented in a number of odontocete species (Palsbøll et al. 1997, Turgeon et al. 2012).
A gallery of some GEMM Lab documented mother-calf gray whale pairs. All images captured under NOAA/NMFS permit #21678. Exclamation (mom; left) and Angie (calf; right).
Scarlett (mom; left) and Brown (calf; right).
Clouds (mom; back) and Cheetah (calf; front).
Xena (mom; back) and Evie (calf; front).
Knife (mom; left) and Daffodil (calf; right).
Pristine (mom; front) and Three Stars (calf; back).
Triplet (mom; right) and White Knuckle (calf; left).
In the GEMM Lab, we are interested in exploring the potential long-term bonds, role and impact of Pacific Coast Feeding Group (PCFG) gray whale mothers on their calves. GEMM Lab PhD student Clara Bird is digging into whether specialized foraging tactics, such as bubble blasts and headstands, are passed down from mothers to calves. I hope to assess whether using the PCFG range as a foraging ground (rather than the Arctic region) is a vertically transmitted behavior or whether environmental factors may play a larger role in the recruitment and dynamics of the PCFG. It will take us a while to get to the bottom of these questions, so in the meantime hug your loved ones if it’s safe to do so or, if you’re in my boat, continue to talk to them virtually until it is safe to be reunited.
References
Barendse, J., Best, P. B., Carvalho, I., and C. Pomilla. 2013. Mother knows best: occurrence and associations of resighted humpback whales suggest maternally derived fidelity to a southern hemisphere coastal feeding ground. PloS ONE 8:e81238.
Burnham, R. E., and D. A. Duffus. 2020. Maternal behaviors of gray whales (Eschrichtius robustus) on a summer foraging site. Marine Mammal Science 36:1212-1230.
Carroll, E. L., Baker, C. S., Watson, M., Alderman, R., Bannister, J., Gaggiotti, O. E., Gröcke, D. R., Patenaude, N., and R. Harcourt. 2015. Cultural traditions across a migratory network shape the genetic structure of southern right whales around Australia and New Zealand. Scientific Reports 5:16182.
Guinet, C., and J. Bouvier. 1995. Development of intentional stranding hunting techniques in killer whale (Orcinus orca) calves at Crozet Archipelago. Canadian Journal of Zoology 73:27-33.
Heide-Jørgensen, M. P., Hammeken, N., Dietz, R., Orr, J., and P. R. Richard. 2001. Surfacing times and dive rates for narwhals and belugas. Arctic 54:207-355.
Lyamin, O., Pryaslova, J., Lance, V., and J. Siegel. 2005. Continuous activity in cetaceans after birth. Nature 435:1177.
Miketa, M. L., Patterson, E. M., Krzyszczyk, E., Foroughirad, V., and J. Mann. 2018. Calf age and sex affect maternal diving behavior in Shark Bay bottlenose dolphins. Animal Behavior 137:107-117.
Noren, S. R. 2008. Infant carrying behavior in dolphins: costly parental care in an aquatic environment. Functional Ecology 22:284-288.
Noren, S. R., Biedenbach, F., Redfern, J. V., and E. F. Edwards. 2007. Hitching a ride: the formation locomotion strategy of dolphin calves. Functional Ecology 22:278-283.
Oftedal, O. T. Lactation in whales and dolphins: evidence of divergence between baleen- and toothed-species. Journal of Mammary Gland Biology and Neoplasia 2:205-230.
Palsbøll, P. J., Heide-Jørgensen, M. P., and R. Dietz. 1996. Population structure and seasonal movements of narwhals, Monodon monoceros, determined from mtDNA analysis. Heredity 78:284-292.
Rendell, L., Cantor, M., Gero, S., Whitehead, H., and J. Mann. 2019. Causes and consequences of female centrality in cetacean societies. Philosophical Transactions of the Royal Society B 374:20180066.
Sargeant, B. L., and J. Mann. 2009. Developmental evidence for foraging traditions in wild bottlenose dolphins. Animal Behavior 78:715-721.
Simões-Lopes, P. C., Daura-Jorge, F. G., and M. Cantor. 2016. Clues of cultural transmission in cooperative foraging between artisanal fishermen and bottlenose dolphins, Tursiops truncatus (Cetacea: Delphinidae). Zoologia (Curitiba) 33:e20160107.
Smultea, M. A., Fertl, D., Bacon, C. E., Moore, M. R., James, V. R., and B. Würsig. 2017. Cetacean mother-calf behavior observed from a small aircraft off Southern California. Animal Behavior and Cognition 4:1-23.
Turgeon, J., Duchesne, P., Colbeck, G. J., Postma, L. D., and M. O. Hammill. 2011. Spatiotemporal segregation among summer stocks of beluga (Delphinapterus leucas) despite nuclear gene flow: implication for the endangered belugas in eastern Hudson Bay (Canada). Conservation Genetics 13:419-433.
Tyson, R. B., Friedlaender, A. S., Ware, C., Stimpert, A. K., and D. P. Nowacek. 2012. Synchronous mother and calf foraging behaviour in humpback whales Megaptera novaeangliae: insights from multi-sensor suction cup tags. Marine Ecology Progress Series 457:209-220.
Villegas-Amtmann, S., Schwarz, L. K., Sumich, J. L., and D. P. Costa. 2015. A bioenergetics model to evaluate demographic consequences of disturbance in marine mammals applied to gray whales. Ecosphere 6:1-19.
Weinrich, M. 1998. Early experience in habitat choice by humpback whales (Megaptera novaeaengliae). Journal of Mammalogy 79:163-170.
Wiley, D., Ware, C., Bocconcelli, A., Cholewiak, D., Friedlaender, A., Thompson, M., and M. Weinrich. 2011. Underwater components of humpback whale bubble-net feeding behavior. Behavior 148:575-602.
Fall has arrived in the Pacific Northwest. For humans, it means packing away the shorts and sandals, and getting the boots, raincoats and firewood ready. For gray whales, it means gulping down the last meal of zooplankton they will eat for several months and commencing the journey to warmer waters and sunnier skies in Mexico where they will spend the winter fasting, calving, and nursing. While the GEMM Lab may still squeeze in a day or two of field work this week, we are slowly wrapping up the 2020 field season as conditions get rougher and our beloved gray whales gradually depart our waters. This year marked the 6th year of data collection for both of our gray whale projects: the Newport project that investigates the impacts of multiple stressors on gray whale ecology and health, and the Port Orford project that explores fine-scale foraging ecology of gray whales and their zooplankton prey. Since it will be several months before the GEMM Lab heads back out onto the water again, I thought I would summarize our two field seasons, share some highlights, and muse about the drivers of our observations this summer.
Some snapshots of the field team hard at work this summer. Top left: Hunter Warwick operating the drone. Top right: PI Leigh Torres rocking the lab swag to protect against the sun. Bottom left: Lisa Hildebrand waiting for a whale to surface to get photos of it. Bottom right: Alejandro Fernandez Ajo transfers zooplankton from the light trap to sample jars.
Summaries
Our RHIB Ruby zipped around the central and southern Oregon coast on 33 different days. The summer started slow, with several days of field work where we encountered no whales despite surveying our entire study region. Our encounters picked up towards the end of June and by the end of the summer we totaled 107 sightings, encountering 46 unique individuals, 36 of which were resightings of known individuals we have identified in previous years. Our Newport star of the summer was Solé, a female gray whale we have seen every year since 2015, and we also saw many of our other regulars including Casper, Rafael, Spray, Bit, and Heart. None of these whales shone as bright as Solé though. We flew the drone over her 8 times and collected 7 fecal samples (one of which was the biggest whale fecal sample I have ever seen!). In total, we collected 30 fecal samples and flew the drone 88 times. These data will allow us to continue measuring body condition and hormone levels of Pacific Coast Feeding Group (PCFG) gray whales that use the Oregon coast.
Left: Our RHIB Ruby heading out for a day of field work in Port Orford, OR this summer. Right: Solé. Image captured under NOAA/NMFS permit #21678.
Our tandem research kayak Robustus may not be as zippy as Ruby (it is powered by human muscle rather than a powerful outboard engine after all), but it certainly continues to be a trusty vessel for the Port Orford team. The Port Orford research team, named the Theyodelers this year, collected 181 zooplankton samples and conducted 180 GoPro drops during the month of August from Robustus. Despite the many samples collected, the size of our prey samples remained relatively small throughout the whole season compared to previous years. The cliff team surveyed for a total of 117 hours, of which 15 were spent tracking whales with the theodolite and resulted in 40 different tracklines of whale movements. The whale situation in Port Orford was similar to the pattern of whale sightings in Newport, with low whale sightings at the start of the field season. Luckily, by the start of August (which marked the start of data collection for the Theyodelers), the number of whales using the Port Orford area, especially the two study sites, Mill Rocks & Tichenor Cove, had increased. Of the whales that came close enough to shore for us to identify using photo-id, we tracked 5 unique individuals, 3 of which we also saw in Newport this year. The Port Orford star of the summer was Smudge, with his tracklines making up a quarter of all of our tracklines collected. Smudge is also the whale we sighted most often last year in Port Orford.
Top: Theyodelers Mattea Holt Colberg and Liz Kelly sampling atop the kayak Robustus. Bottom: Smudge. Image captured under NOAA/NMFS permit #21678.
Highlights
Many of you may be familiar with the whale Scarlett (formally known as Scarback). Scarlett is a female, at least 24 years old (she was first documented in the PCFG range in 1996), who is well-known (and easily identified) by the large concave injury on her back that is covered in whale lice, or cyamids. No one knows for certain how Scarlett sustained this injury (though there are stories), however what we do know is that it has not prevented this female from reproducing and successfully raising several calves over her lifetime. The GEMM Lab last saw Scarlett with a calf (which we named Brown) in 2016. Since Scarlett is such a famous whale with a unique history, it shouldn’t be a surprise that one of our highlights this summer is the fact that Scarlett showed up with a new calf! In keeping with a “shades of red” theme, Leigh came up with the name Rose for the new calf. In July, the mom-calf pair put on quite a cute performance, with Rose rising up on Scarlett’s back, giving the team a glimpse of its face. The Scarlett-Rose highlight doesn’t end there though. Just last week, we had a very brief encounter in choppy, swelly waters with a small whale. The whale surfaced just twice allowing us to capture photo-id images, and as we were looking around to see where it would come up a third time, it suddenly breached approximately 20 m from the boat. Lo-and-behold, after comparing our photos of the whale to our catalogue, we realized that this elusive, breaching whale was Rose! I am excited to see whether Rose will return to the Oregon coast next summer and become a PCFG regular just like her mom.
Top: Scarlett, easily identified by the large concave injury on her back covered in cyamids. Bottom: Scarlett with her new calf Rose riding her back, giving us a glimpse of its face. Images captures under NOAA/NMFS permit #21678.
The highlight of the field season in Port Orford is the trial, failures and small successes of a new element to the project. There is still a lot that we do not know and understand about PCFG gray whales. One such thing is the way in which gray whales maneuver their large bodies in shallow rocky habitats, often riddled with kelp, and how exactly they capture their zooplankton prey in these environments. Using drones has certainly helped bring some light into this darkness and has led to the documentation of many novel foraging behaviors (Torres et al. 2018). However, the view from above is unable to provide the fine-scale interactions between whales, kelp, reefs, and zooplankton. Instead, we must somehow find a way to watch the whales underwater. Enter CamDo. CamDo is a technology company that designs specialty products to allow for GoPro cameras to be used for time-lapsed recordings over long periods of time in harsh environmental conditions. One of their products is a housing specifically designed for long-term filming underwater – exactly what we need! The journey was not as easy as simply purchasing the housing. We also needed to build a lander for the housing to sit on (thankfully our very own Todd Chandler designed and built something for us), and coordinate with divers and a vessel to deploy and retrieve the set-up, as well as undertake weekly battery and SD cards swaps (thankfully Dave Lacey of South Coast Tours and a very generous group of divers* donated their time and resources to make this happen). We unfortunately had some technological difficulties and bad visibility for the first 4 weeks (precisely why this CamDo effort was a pilot season this year), however we had some small success in the last 2 weeks of deployment that give us hope for the future. The camera recorded a lot of things: thick layers of mysids, countless rockfish and lingcod, several swimming and foraging murres, a handful of harbor seals, and two encounters of the species we were hoping to film – gray whales! While the footage is not the ‘money shot’ we are hoping to film (aka, a headstanding gray whale eating zooplankton right in front of the camera), the fact that we captured gray whales in the first place has showed us that this set-up is a promising investment of time, money and effort that will hopefully deliver next year.
Top left: CamDo atop its lander designed and built by Todd Chandler. Top right: Divers getting ready to get into the water for a battery and SD card swap. Bottom: Dave Lacey and two of our divers with the Black Pearl, South Coast Tour’s boat.
Musings
You may have picked up on the fact that we had slow starts to our field seasons in both Newport and Port Orford. Furthermore, while the number of whale sightings did increase in both locations throughout the field seasons, the number of sightings and whales per day were lower than they have been in previous years. For example, in 2018, we identified 15 different individuals in the month of August in Port Orford (compared to just 5 this year). In 2019, 63 unique whales were seen in Newport (compared to 46 this year). Interestingly, we had a greater diversity of encountered individuals at the start and end of the season in Newport, with a relatively small number of different individuals in July and August. While I cannot provide a definitive reason (or reasons) as to why patterns were observed (we will need to analyze several years of our data to try and understand why), I have some hypotheses I wish to share with you.
As I mentioned in a previous blog, this summer the coastal upwelling along the Oregon coast was delayed (Figure 1). Typically, peak upwelling occurs during the month of June or shortly thereafter, bringing nutrient-rich, deep waters to the surface and, when mixed with sunlight, a lot of productivity. This productivity sets off a chain of reactions — the input of nutrients leads to increased phytoplankton production, which in turn leads to increased zooplankton production, resulting in growth and development of larger organisms that consume zooplankton, such as rockfish and gray whales. If the timing of upwelling is delayed, then so too is this chain of reactions. As you can see from Figure 1, the red lines show that the peak upwelling this year occurred far later in the summer than any year in the last 10 years, with the exception of 2012. Gray whales may have cued into this delay and therefore also delayed their arrival to the PCFG feeding grounds, hence causing us to have low sighting rates at the start of our season. However, this is mostly speculative as we still do not understand the functional mechanisms by which cetaceans, such as gray whales, detect prey across different scales, and to what extent oceanographic conditions like upwelling may play a role in prey availability (Torres 2017).
Figure 1. 10 year time series of the Coastal Upwelling Transport Index (CUTI). CUTI represents the amount of upwelling (positive numbers) or downwelling (negative numbers). The light-colored lines representthe CUTI at that point in time while the dark, bold line represents the long-term average.The vertical red lines represent the point of peak upwelling in that summer and the horizontal green line shows the peak level of upwelling in 2020 relative to all previous years.
Furthermore, the green line in Figure 1shows that even after peak upwelling was reached this year, upwelling conditions were lower than all the other peaks in the previous 10 years. We know that weak upwelling is correlated to poor body condition of PCFG gray whales in subsequent years (Soledade Lemos et al. 2020). Upon arriving to the Oregon coast feeding grounds, gray whales may have noticed that it was shaping up to be a poor prey year (we certainly noticed it in Port Orford in the emptiness of our zooplankton net). Faced with this low resource availability, individuals had to make important decisions – risk staying in a currently prey-poor environment or continue the journey onward, searching for better prey conditions elsewhere. This conundrum is known as the marginal value theorem, whereby an individual must decide whether it should abandon the patch it is currently foraging on and move on to search for a new patch without knowing how far away the next patch may be or its value relative to the current patch (Charnov 1976). If we think of the Oregon coast as the ‘current patch’, then we can see how the marginal value theorem translates to the situation gray whales may have found themselves in at the start of the summer.
Yet, an individual gray whale does not make these decisions in a vacuum. Instead, all gray whales in the same area are faced with the same conundrum. Seminal work by Pianka (1974) showed that when resources, such as food, are abundant, then competition between predators is low because there is enough food to go around. However, when resources dwindle, competition increases and the niches of predators begin to overlap more and more. With Charnov and Pianka’s theories in mind, we can see two groups of gray whales emerge from our 2020 field work observations: those that stayed in the ‘current patch’ (Oregon) and those that decided to seek out a new patch in hopes that it would be a better one. Solé certainly belongs in the first group. We saw her consistently throughout the whole summer. In fact, she was oftentimes so predictable that we would find her foraging on the same reef complex every time we went out to survey. Smudge may also belong in this group, however it is hard to say definitively since we only survey in Port Orford in late July and August. In contrast, I would place whales such as Spray and Heart in the second group since we saw them early in the summer and then not again until mid-to-late September. Where did they go in the interim? Did they go somewhere else in the PCFG range? Or did they venture all the way up to Alaska to the primary Eastern North Pacific (ENP) gray whale feeding grounds? Did their choice to search for food elsewhere pay off?
As I said earlier, these are all just musings for now, but the GEMM Lab is already hard at work trying to answer these questions. Stay tuned to see what we find!
* Thanks to all the divers who assisted with the pilot CamDo season: Aaron Galloway, Ross Whippo, Svetlana Maslakova, Taylor Eaton, Cori Kane, Austin Williams, Justin Smith
References
Charnov, E.L. 1976. Optimal Foraging, the Marginal Value Theorem. Theoretical Population Biology 9(2):129-136.
Pianka, E.R. 1974. Niche Overlap and Diffuse Competition. PNAS 71(5):2141-2145.
Soledade Lemos, L., Burnett, J.D., Chandler, T.E., Sumich, J.L., and L.G. Torres. 2020. Intra- and inter-annual variation in gray whale body condition on a foraging ground. Ecosphere 11(4):e03094.
Torres, L.G. 2017. A sense of scale: Foraging cetaceans’ use of scale-dependent multimodal sensory systems. Marine Mammal Science 33(4):1170-1193.
Torres, L.G., Nieukirk, S.L., Lemos, L., and T.E. Chandler. 2018. Drone Up! Quantifying Whale Behavior From a New Perspective Improves Observational Capacity. Frontiers in Marine Science: https://doi.org/10.3389/fmars.2018.00319.
When humans count calories it is typically to regulate and limit calorie intake. What I am wondering about is whether gray whales are aware of caloric differences in the prey that is available to them and whether they make foraging decisions based on those differences. In last week’s post, Dawn discussed what makes a good meal for a hungry blue whale. She discussed that total prey biomass of a patch, as well as how densely aggregated that patch is, are the important factors when a blue whale is picking its next meal. If these factors are important for blue whales, is it same for gray whales? Why even consider the caloric value of their prey?
Gray and blue whales are different in many ways; one way is that blue whales are krill specialists whereas gray whales are more flexible foragers. The Pacific Coast Feeding Group (PCFG) of gray whales in particular are known to pursue a more varied menu. Previous studies along the PCFG range have documented gray whales feeding on mysid shrimp (Darling et al. 1998;Newell 2009), amphipods (Oliver et al. 1984; Darling et al. 1998), cumacean shrimp (Jenkinson 2001; Moore et al. 2007;Gosho et al. 2011), and porcelain crab larvae (Dunham and Duffus 2002), to name a few. Based on our observations in the field and from our drone footage, we have observed gray whales feeding on reefs (likely on mysid shrimp), benthically (likely on burrowing amphipods), and at the surface on crab larvae (Fig. 1). Therefore, while both blue and PCFG whales must make decisions about prey patch quality based on biomass and density of the prey, gray whales have an extra decision to make based on prey type since their prey menu items occupy different habitats that require different feeding tactics and amount of energy to acquire them. In light of these reasons, I hypothesize that prey caloric value factors into their decision of prey patch selection.
Figure 1. Gray whales use several feeding tactics to obtain a variety of coastal Oregon zooplankton prey including jaw snapping (0:12 of video), drooling mud (0:21), and head standing (0:32), to name a few.
This prey selection process is crucial since PCFG gray whales only have about 6 months to consume all the food they need to migrate and reproduce (even less for the Eastern North Pacific (ENP) gray whales since their journey to their Arctic feeding grounds is much longer). You may be asking, well if feeding is so important to gray whales, then why not eat everything they come across? Surely, if they ate every prey item they swam by, then they would be fine. The reason it isn’t quite this simple is because there are energetic costs to travel to, search for, and consume food. If an individual whale simply eats what is closest (a small, poor-quality prey patch) and uses up more energy than it gains, it may be missing out on a much more beneficial and rewarding prey patch that is a little further away (that patch may disperse or another whale may eat it by the time this whale gets there). Scientists have pondered this decision-making process in predators for a long time. These ponderances are best summed up by two central theories: the optimal foraging theory (MacArthur & Pianka 1966) and the marginal value theorem (Charnov 1976). If you are a frequent reader of the blog, you have probably heard these terms once or twice before as a lot of the questions we ask in the GEMM Lab can be traced back to these concepts.
Optimal foraging theory (OFT) states that a predator should pick the most beneficial resource for the lowest cost, thereby maximizing the net energy gained. So, a gray whale should pick a prey patch where it knows that it will gain more energy from consuming the prey in the patch than it will lose energy in the process of searching for and feeding on it. Marginal value theorem elaborates on this OFT concept by adding that the predator also needs to consider the cost of giving up a prey patch to search for a new one, which may or may not end up being more profitable or which may take a very long time to find (and therefore cost more energy).
The second chapter of my thesis will investigate whether individual gray whales have foraging preferences by relating feeding location to prey quality (community composition) and quantity (relative density). However, in order to do that, I first must know about the quality of the individual prey species, which is why my first chapter explores the caloric content of common coastal zooplankton species in Oregon that may serve as gray whale prey. The lab work and analysis for that chapter are completed and I am in the process of writing it up for publication. Preliminary results (Fig. 2) show variation in caloric content between species (represented by different colors) and reproductive stages (represented by different shapes), with a potential increasing trend throughout the summer. These results suggest that some species and reproductive stages may be less profitable than others based solely on caloric content.
Figure 2. Mean caloric content (J/mg) of coastal Oregon zooplankton (error bars represent standard deviation) from May-October in 2017-2018. Colors represent species and shapes represent reproductive stage.
Now that we have established that there may be bigger benefits to feeding on some species over others, we have to consider the availability of these zooplankton species to PCFG whales. Availability can be thought of in two ways: 1) is the prey species present and at high enough densities to make searching and foraging profitable, and 2) is the prey species in a habitat or depth that is accessible to the whale at a reasonable energetic cost? Some prey species, such as crab larvae, are not available at all times of the summer. Their reproductive cycles are pulsed (Roegner et al. 2007) and therefore these prey species are less available than species, such as mysid shrimp, that have more continuous reproduction (Mauchline 1980). Mysid shrimp appear to seek refuge on reefs in rock crevices and among kelp, whereas amphipods often burrow in soft sediment. Both of these habitat types present different challenges and energetic costs to a foraging gray whale; it may take more time and energy to dislodge mysids from a reef, but the payout will be bigger in terms of caloric gain than if the whale decides to sift through soft sediment on the seafloor to feed on amphipods. This benthic feeding tactic may potentially be a less costly foraging tactic for PCFG whales, but the reward is a less profitable prey item.
My first chapter will extend our findings on the caloric content of Oregon coastal zooplankton to facilitate a comparison to the caloric values of the main ampeliscid amphipod prey of ENP gray whales feeding in the Arctic. Through this comparison I hope to assess the trade-offs of being a PCFG whale rather than an ENP whale that completes the full migration cycle to the primary summer feeding grounds in the Arctic.
References
Charnov, E. L. 1976. Optimal foraging: the marginal value theorem. Theoretical Population Biology 9:129-136.
Darling, J. D., Keogh, K. E. and T. E. Steeves. 1998. Gray whale (Eschrichtius robustus) habitat utilization and prey species off Vancouver Island, B.C. Marine Mammal Science 14(4):692-720.
Dunham, J. S. and D. A. Duffus. 2002. Diet of gray whales (Eschrichtius robustus) in Clayoquot Sound, British Columbia, Canada. Marine Mammal Science 18(2):419-437.
Gosho, M., Gearin, P. J., Jenkinson, R. S., Laake, J. L., Mazzuca, L., Kubiak, D., Calambokidis, J. C., Megill, W. M., Gisborne, B., Goley, D., Tombach, C., Darling, J. D. and V. Deecke. 2011. SC/M11/AWMP2 submitted to International Whaling Commission Scientific Committee.
Jenkinson, R. S. 2001. Gray whale (Eschrichtius robustus) prey availability and feeding ecology in Northern California, 1999-2000. Master’s thesis, Humboldt State University.
MacArthur, R. H., and E. R. Pianka. 1966. On optimal use of a patchy environment. American Naturalist 100:603-609.
Mauchline, J. 1980. The larvae and reproduction in Blaxter, J. H. S., Russell, F. S., and M. Yonge, eds. Advances in Marine Biology vol. 18. Academic Press, London.
Moore, S. E., Wynne, K. M., Kinney, J. C., and C. M. Grebmeier. 2007. Gray whale occurrence and forage southeast of Kodiak Island, Alaska. Marine Mammal Science 23(2)419-428.
Newell, C. L. 2009. Ecological interrelationships between summer resident gray whales (Eschrichtius robustus) and their prey, mysid shrimp (Holmesimysis sculpta and Neomysis rayii) along the central Oregon coast. Master’s thesis, Oregon State University.
Oliver, J. S., Slattery, P. N., Silberstein, M. A., and E. F. O’Connor. 1984. Gray whale feeding on dense ampeliscid amphipod communities near Bamfield, British Columbia. Canadian Journal of Zoology 62:41-49.
Roegner, G. C., Armstrong, D. A., and A. L. Shanks. 2007. Wind and tidal influences on larval crab recruitment to an Oregon estuary. Marine Ecology Progress Series 351:177-188.
There is something special about the Oregon Coast. It’s like nowhere else in the world. When Lisa told me that gray whales are understudied on our coastline, I secretly and selfishly thought to myself, “I hope it stays that way”. Then I would have a chance to be a pioneer one day too, studying something along this rugged coast full of life, death and everything in between, that no one has answered before. Of course, I only feel this way half of the time.
Yet, the more time I spend in Port Orford, the more I realize that our coastline truly is one of those last frontiers. A place where fundamental questions have yet to be explored, where the passing of seasons brings with it a violent change in conditions. From sunny summer days on the Port Orford beaches taking in the soft glistening of sunlight illuminating Redfish Rocks Marine Reserve, to cold, dark and stormy months with no end in sight and nothing but the sound of wind curving around the bends of your home and rain puttering against the windows.
Noah reading a book on the cliff site with a view of Mill Rocks in the background. Source: N. Dolinajec.
But no matter the season, no matter the conditions, the Oregon Coast harnesses something truly special, truly extraordinary. A cyclical diversity of life.
Since I was a kid, the Oregon Coast has inspired me. Not always to think about wildlife, in fact, mostly in other ways. To contemplate more primal philosophical questions. At 28 years old, it’s been a longer road than expected to get to this point, working with these amazing people, in this amazing place, on this amazing project. And the more time that passes, the more failures, missteps and dysfunctional experiences I absorb, the more that I learn about what really needs to change. In the world of course, but, mostly in science.
In the past few years, as I eek closer to 30, and I begin to look back on some of the adventures I have taken in my life, I take heavy note of where I am now, sitting on a kayak in Mill Rocks sampling for gray whale prey abundance and distribution, or atop the cliff, gazing out into the open ocean waiting patiently and graciously (at least trying to be) for a small poof of water spray from the beating surface of the sea. That little poof? It may not seem like much but it’s a sign of life. Of an age-old journey, one we know very little about. And here I am, a part of it, albeit a small one, but nevertheless, forever a part of that great journey.
A foggy view of Mill Rocks from the cliff (left) and a shot from the kayak (right). Source: N. Dolinajec.
And without losing sight of my job, sampling for zooplankton or tracking the whales as they move across the open water, I’ve found myself thinking about the depth of being involved in such an ancient process, and considering a very important question. One that doesn’t spend nearly enough time in the day-to-day conversation of an academic…
What exactly is a scientist? And how does one become a scientist?
The academic path to the sciences is exclusionary, beyond any reasonable level. It discriminates on gender, race, experience and age. Making the sciences, which are meant as a tool to better the world and make useful contributions to society and the future, feel inaccessible for so many people full of potential but without the right boxes ticked on a form.
How many beautiful ideas have been left to decay because of the ego that science has built for itself?
A sign that sits in the front window of the OSU Port Orford Field Station. Source: N. Dolinajec.
Don’t get me wrong, I love science, it has given me joy that other things in life cannot. It has shown me both the complexity of the world and the simplicity of how we view it. And I believe that science can still be the future. But in order for science to command our future, to guide us in the right direction, it cannot be a hierarchy of antiquated procedures any longer. We must open our arms, our minds and our resources to take chances on students, far and wide, that may lack traditional training but instead have other skills or experiences to offer science. Science needs an overhaul. Science needs diversity.
After all, change of perspective can be a profound driver of scientific results, can it not?
Here in Port Orford, in this bizarre year of 2020, we have the beginning, the makings if-you-will, of that very diversity that I am speaking of. The four of us, ‘The Theyodelers’ as we righteously call ourselves, each come from such drastically different places in life only to meet under the same roof for 6 weeks and miraculously not only survive together, but thrive together.
‘The Theyodelers’ after the 2020 (virtual) Port Orford Community Presentation, from left to right: Dr. Leigh Torres, Lisa Hildebrand, Liz Kelly, Mattea Holt Colberg, Noah Dolinajec, Tom Calvanese, Tom McCambridge (front). Source: L. Hildebrand.
And that, that essence of positivity that we have been able to build around one another this season, is exactly what I mean when I say that science needs an overhaul.
We do not all find our way to this moment, doing science in such an inspiring place, in the same way. Some of us are born with the innate ability to see the world through objective eyes, the kind of mind that makes great science happen from an early age. And others find our way to science after being enlightened by trials and travails, failures and mistakes, missed opportunities and missteps.
No matter the journey, we all ended up here. Watching these great gray giants on their journeys.
And it all comes full circle doesn’t it?
Each of our journeys, human or whale, can lead to the very same point despite beginning at very different places. And in that diversity of experience, of life, of age, of color, is where we find our brightest moments, our grandest ideas and our future, driven by science.





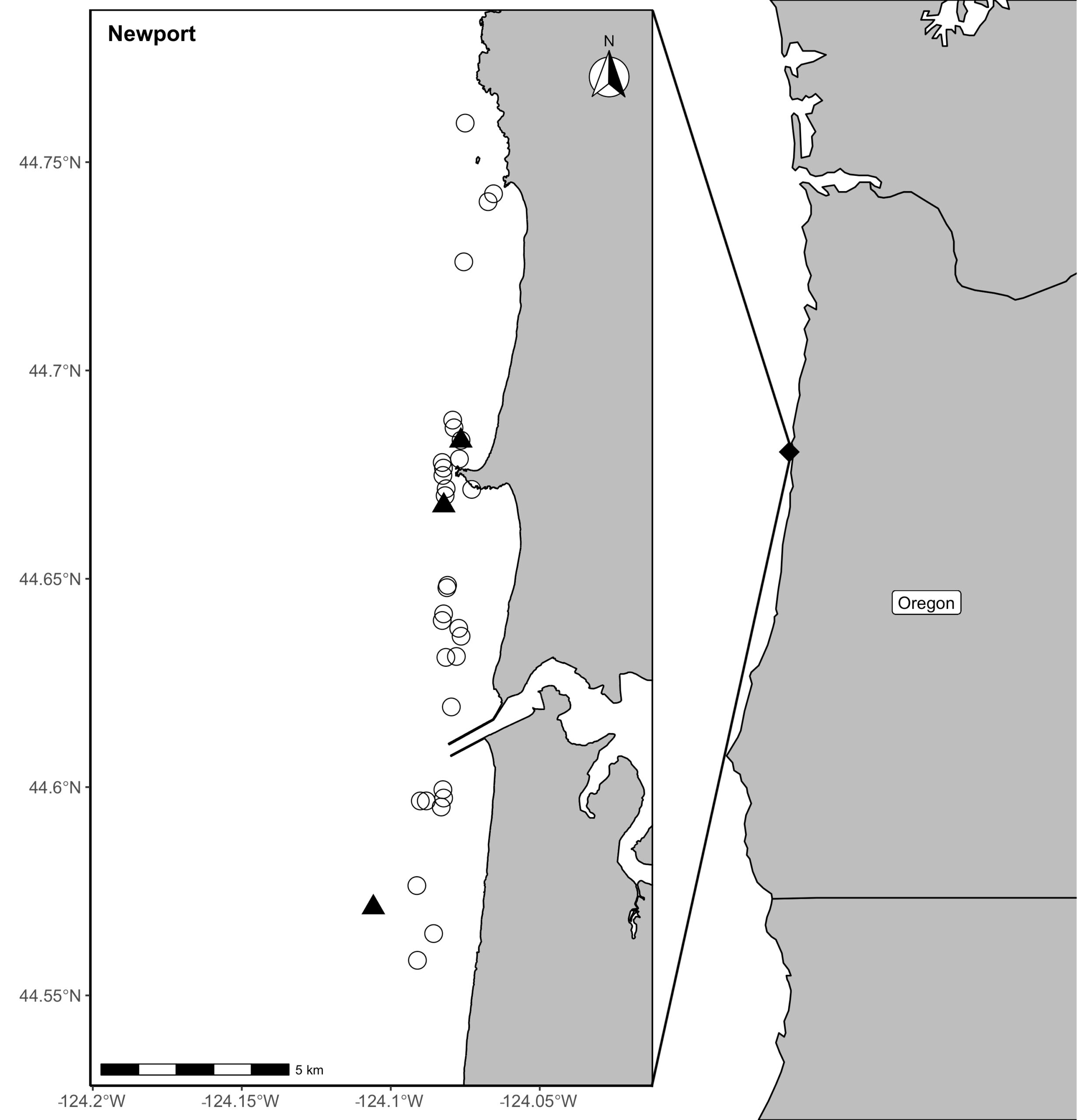
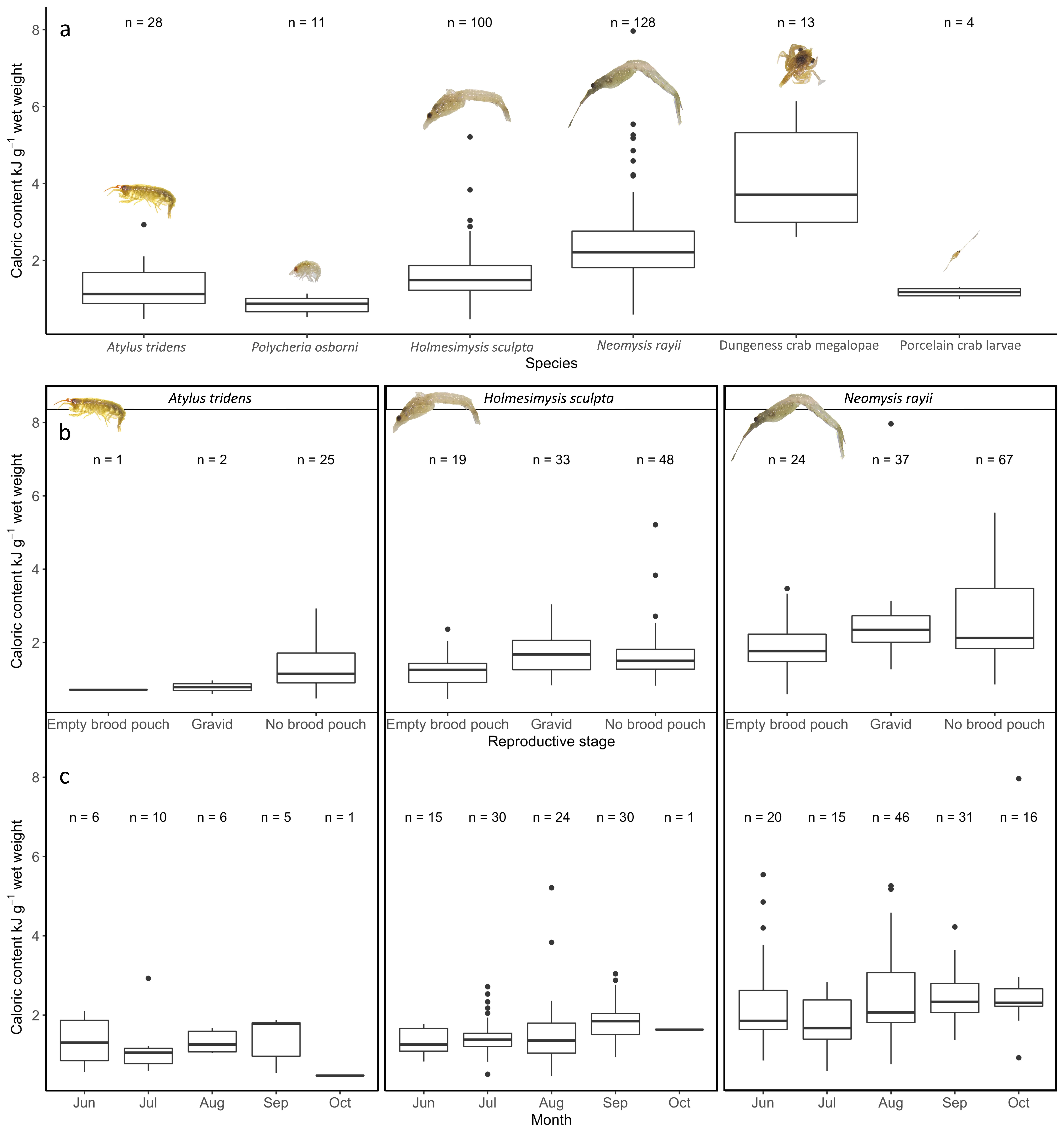


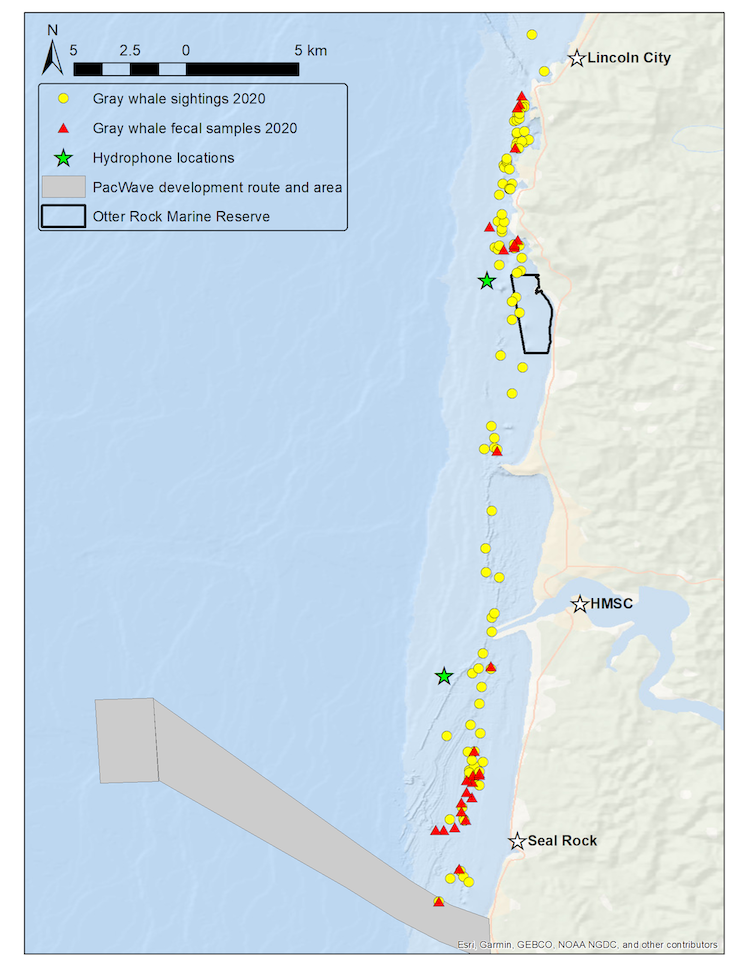

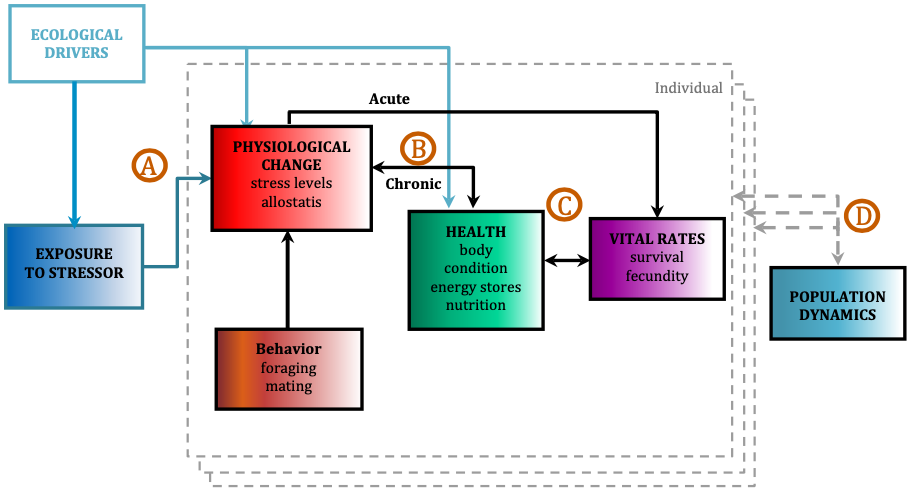































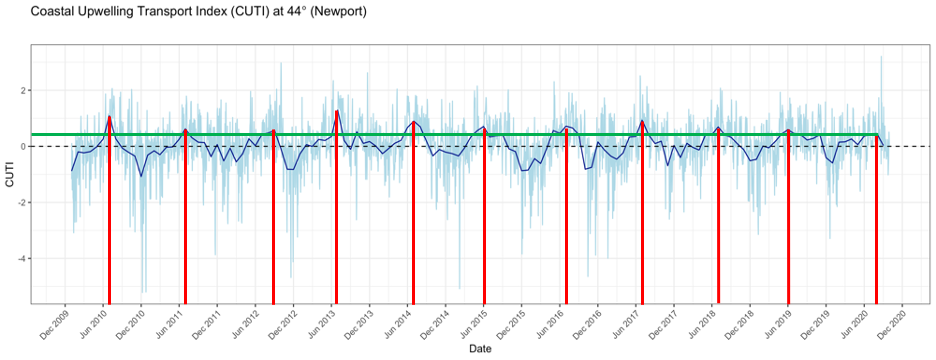
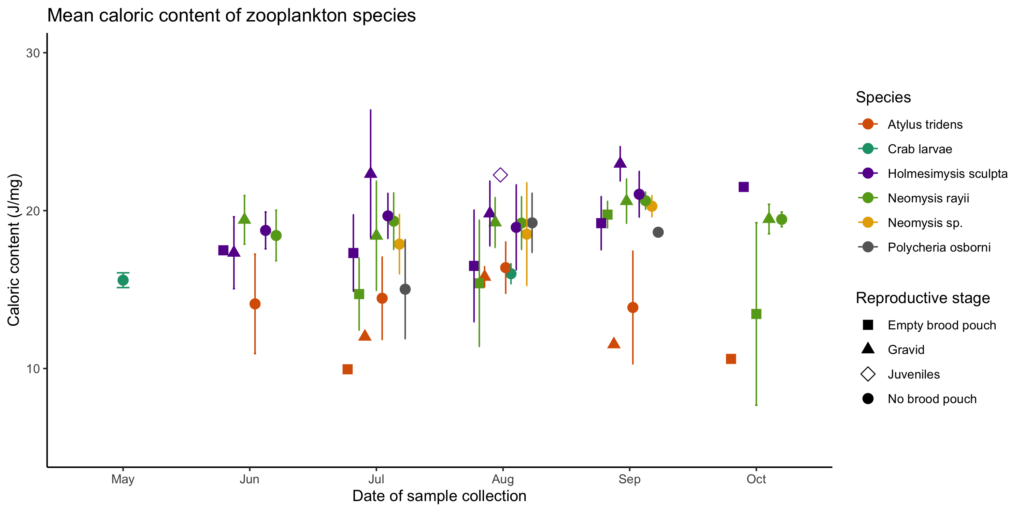





{kind=link}