Dr. Enrico Pirotta (CREEM, University of St Andrews) and Dr. Leigh Torres (GEMM Lab, MMI, OSU)
The health of animals affects their ability to survive and reproduce, which, in turn, drives the dynamics of populations, including whether their abundance trends up or down. Thus, understanding the links between health and reproduction can help us evaluate the impact of human activities and climate change on wildlife, and effectively guide our management and conservation efforts. In long-lived species, such as whales, once a decline in population abundance is detected, it can be too late to reverse the trend, so early warning signals are needed to indicate how these populations are faring.
We worked on this complex issue in a study that was recently published in the Journal of Animal Ecology. In this paper, we developed a new statistical approach to link three key components of the health of a Pacific Coast Feeding Group (PCFG) gray whale (namely, its body size, body condition, and stress levels) to a female’s ability to give birth to a calf. We were able to inform these metrics of whale health using an eight-year dataset derived from the GRANITE project of aerial images from drones for measurements of body size and condition, and fecal samples for glucocorticoid hormone analysis as an indicator of stress. We combined these data with observations of females with or without calves throughout the PCFG range over our study period.
We found that for a female to successfully have a calf, she needs to be both large and fat, as these factors indicate if the female has enough energy stored to support reproduction that year (Fig. 1). Remarkably, we also found indication that females with particularly high stress hormone levels may not get pregnant in the first place, which is the first demonstration of a link between stress physiology and vital rates in a baleen whale, to our knowledge.
Figure 1. Taken from Pirotta et al. (2025), Fig. 5. Combined relationship of PCFG gray whale length and nutritional state (combination of body size and condition) in the previous year with calving probability, colored by whether the model estimated an individual to have calved or not at a given reproductive opportunity.
Our study’s findings are concerning given our previous research indicating that gray whales in this PCFG sub-group have been growing to shorter lengths over the last couple of decades (Pirotta et al. 2023), are thinner than animals in the broader Eastern North Pacific gray whale population (Torres et al, 2022), and show an increase in stress-related hormones when exposed to human activities (Lemos et al, 2022; Pirotta et al. 2023). Furthermore, in our recent study we also documented that there are fewer young individuals than expected for a growing or stable population (Fig. 2), which can be an indicator of a population in decline since there may not be many individuals entering the reproductive adult age groups. Altogether, our results act as early warning signals that the PCFG may be facing a possible population decline currently or in the near future.
Figure 2. Taken from Pirotta et al. (2025), Fig. 1. Age structure diagram for 139 PCFG gray whales in our dataset. Each bar represents the number of individuals of a given age in 2023, with the color indicating the proportion of individuals of that age for which age is known (vs. estimated from a minimum estimate following Pirotta, Bierlich, et al., 2024). The red line reports a smooth kernel density estimate of the distribution.
These findings are sobering news for Oregon residents and tourists who enjoy watching these whales along our coast every summer and fall. We have gotten to know many of these whales so well – like Scarlett, Equal, Clouds, Lunita, and Pacman, who you can meet on our IndividuWhale website – that we wonder how they will adapt and survive as their once reliable habitat and prey-base changes. We hope our work sparks collective and multifaceted efforts to reduce impacts on these unique PCFG whales, and that we can continue the GRANITE project for many more years to come to monitor these whales and learn from their response to change.
This work exemplifies the incredible value of long-term studies, interdisciplinary methods, and effective collaboration. Through many years of research on this gray whale group, we have collected detailed data on diverse aspects of their behavior, ecology and life history that are critical to understanding their response to disturbance and environmental change, which are both escalating in the study region. We are incredibly grateful to the following members of the PCFG Consortium for contributing sightings and calf observation data that supported this study: Jeff Jacobsen, Carrie Newell, NOAA Fisheries (Peter Mahoney and Jeff Harris), Cascadia Research Collective (Alie Perez), Department of Fisheries and Oceans, Canada (Thomas Doniol-Valcroze and Erin Foster), Mark Sawyer and Ashley Hoyland, Wendy Szaniszlo, Brian Gisborne, Era Horton.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly alert when we make a new post! Just add your name into the subscribe box below!
References:
Lemos, Leila S., Joseph H. Haxel, Amy Olsen, Jonathan D. Burnett, Angela Smith, Todd E. Chandler, Sharon L. Nieukirk, Shawn E. Larson, Kathleen E. Hunt, and Leigh G. Torres. “Effects of Vessel Traffic and Ocean Noise on Gray Whale Stress Hormones.” Scientific Reports 12, no. 1 (2022): 18580. https://dx.doi.org/10.1038/s41598-022-14510-5.
Pirotta, Enrico, Alejandro Fernandez Ajó, K. C. Bierlich, Clara N Bird, C Loren Buck, Samara M Haver, Joseph H Haxel, Lisa Hildebrand, Kathleen E Hunt, Leila S Lemos, Leslie New, and Leigh G Torres. “Assessing Variation in Faecal Glucocorticoid Concentrations in Gray Whales Exposed to Anthropogenic Stressors.” Conservation Physiology 11, no. 1 (2023). https://dx.doi.org/10.1093/conphys/coad082.
Torres, Leigh G., Clara N. Bird, Fabian Rodríguez-González, Fredrik Christiansen, Lars Bejder, Leila Lemos, Jorge Urban R, et al. “Range-Wide Comparison of Gray Whale Body Condition Reveals Contrasting Sub-Population Health Characteristics and Vulnerability to Environmental Change.” Frontiers in Marine Science 9 (2022). https://doi.org/10.3389/fmars.2022.867258. https://www.frontiersin.org/article/10.3389/fmars.2022.867258
When most people think about monitoring the health of a 40-ton gray whale, they picture blubber thickness, dive patterns, or perhaps growth rates. But what if some of the most telling signs are found not in the whale’s bulk, but right on the surface–embedded in its skin, and even crawling across it?
As part of the GRANITEproject, my research focuses on using a long-term photographic dataset (>347,000 photos from 10 years!) to evaluate epidermal indicators of stress and health in Pacific Coast Feeding Group (PCFG) gray whales (Eschrichtius robustus) foraging off the Oregon coast. My central questions ask:
Can we use features visible on the skin like epidermal diseases, lesion severity, scarring from orcas, boats, and fishing gear, and potentially cyamid loads as biomarkers of physiological stress or nutritional status?
How do these skin-based indicators correlate with environmental variables, prey availability, fecal hormones, and overall body condition?
By tracking these patterns across individuals and years, my goal is to understand how gray whales are responding to a changing ocean and whether their skin can tell us more about what’s under the surface.
What are cyamids?
Cyamids, more commonly known as “whale lice”, are small crustaceans that live exclusively on marine mammals. Despite their nickname, cyamids are not true lice—they’re actually amphipod ectoparasites, more closely related to beach hoppers than anything you’d find in your hair. For gray whales (Eschrichtius robustus), these tiny passengers are a constant presence throughout their lives.
Figure 1. Gray whale blow hole area covered in barnacles and cyamids. The circle inset shows a zoomed in area where you can see the orange cyamids aggregating near the more yellow barnacles.
Each whale can host thousands of cyamids at a time, with individuals often clustering in specific areas of the body that provide physical refuge from the currents: around the blowhole, in the crevices of flukes, along the rostrum, genital slits, and especially around wounds or skin irregularities (Figure 1). Unlike barnacles, which attach directly to the skin and remain stationary while they feed on nutrients in the passing water, cyamids grasp onto the whale’s body using claw-like appendages, feeding on sloughed skin and bodily fluids. This relationship is generally not thought to be harmful to the whale, but high cyamid loads can be indicative of poor health, injury, or compromised immune function.
There are several species of cyamids, and many are host-specific—meaning they’ve evolved alongside particular whale species. In gray whales, the most common is Cyamus scammoni, which specializes on gray whales and is rarely found elsewhere. Other species found on gray whales include Cyamus kessleri, and the rarer Cyamus ceti. Cyamids are transmitted primarily from mother to calf, which helps explain their host fidelity, but horizontal transmission (between unrelated individuals) may also occur during close contact which can explain some rare occurrences of cyamids that are found outside of their general host species. In fact, Cyamus ceti was only found once on gray whales in 1861 but is generally thought to be specific to bowhead whales, giving us potential insight into interspecies interactions (bowhead and gray whales can spatially overlap on Arctic foraging grounds).
Figure 2. Cyamus scammoni close up photographs of (A) aggregation, (B) juvenile stage, (C) dorsal side of an adult, and (D) ventral side of an adult showcasing the cyamids corkscrew shaped gills (Takeda et al. 2005)
Because cyamids are permanent residents of the whale’s skin, they offer a unique window into both individual whale life histories and broader ecological trends. Their location, abundance, and distribution can potentially inform us about wound healing, residency duration in foraging areas, and even stress or health status—which makes them an unexpectedly valuable focal point in drone and photograph-based monitoring efforts like in the GRANITE project.
Cyamid Life History
Cyamids are obligate ectoparasites, meaning they spend their entire life on a whale and cannot survive independently in the open ocean. Unlike free-swimming crustaceans, cyamids are permanently attached to the skin or embedded within crevices of the whale’s body, often clinging to roughened areas, scars, embedded barnacles, or calloused skin where they can anchor themselves more securely.
They begin life as tiny juveniles, hatching from eggs carried in the brood pouch of a female cyamid. Rather than undergoing a larval phase in the water column like many marine invertebrates, cyamids develop directly into miniature versions of adults and remain on the whale from birth. This direct development is essential because there’s no safe habitat for a larval cyamid in the open ocean: the host whale is both nursery and home.
Most transmission occurs from mother to calf during the close physical contact of early life. Calves born in the warm lagoons of Baja California, Mexico where gray whales calve and nurse during the winter inherit their cyamid colonies during nursing, rubbing, and swimming alongside their mothers. These early colonizers will multiply as the calf grows and can remain with the whale for years, forming the basis of a persistent, host-specific population.
For Cyamus scammoni specifically (our gray whale specific cyamid), adults will breed in the summer just before the southbound migration. Females will have around 1,000 eggs in their brood pouch, although only about a 60% are fertilized (Leung, 1976). These eggs will hatch in the fall while the gray whales take on their southbound migration but they will stay in the safety of the brood pouch for around 2 to 3 months. The juveniles will be released in the winter, when gray whales arrive in the Baja lagoons where they will then find shelter within the crevices of their host gray whale. Juveniles reach maturity during the northbound migration and will be a full-grown brood upon arrival to summer grounds. While the cycle takes about 8 months to complete, there are juveniles found along the gray whales year-round, leading us to believe that there is likely overlap between broods. For our less abundant Cyamus kessleri, the life cycle is very similar, but the juveniles reach maturity before the gray whales northbound migration to summer feeding grounds. Also, there are around 300 eggs in the Cyamus kessleri brood pouches that have a higher rate of fertilization (75-80%) than Cyamus scammoni (60%) (Leung, 1976)
In short, the life of a cyamid is fully bound to the life of a whale. Every migration, dive, foraging event, and scar the whale experiences becomes part of the cyamid’s environment. By studying them, we gain another lens through which to interpret the health, behavior, and ecology of gray whales on the Oregon Coast.
Uses in Cetacean Health Assessments
As we’ve established, cyamids have unique life histories as ectoparasites and may be valuable indicators in cetacean health assessments across multiple whale species. Because they often congregate around wounds, lesions, and areas of poor skin integrity, their presence and distribution can reveal important clues about a whale’s physical condition, injury history, and immune response. However, studies that have made these connections have variable results.
In species like North Atlantic right whales (Pettis et al. 2004, Pirotta et al. 2023), harbor porpoises (Lehnert et al. 2021), and gray whales (Raverty et al. 2024), researchers have used visual surveys and photographic analysis to quantify cyamid loads in living, stranded, and hunted whales. Researchers can score cyamid presence by identifying attachment sites (e.g. blowhole, scar, dorsal ridge) and estimating the relative coverage by using standardized reference images to maintain consistency. In these studies, whales with heavy cyamid coverage, especially in sensitive regions like the blowhole, mouthline, and genital area, often show signs of poor health or stress, such as emaciation, scarring from entanglement, or chronic skin conditions. Cyamid coverage is sometimes used alongside body condition indices and lesion scoring to build a more complete health profile (Pirotta et al. 2023). There are also studies that show no connections, or even positive connections between body condition and cyamid coverage (Von Duyke et al. 2016).
While cyamids are often associated with injured, inflamed, or otherwise damaged skin, there is no evidence that points towards cyamids directly damaging the skin themselves. However, more work needs to be done to assess their role in the healing processes. Additionally, it’s been noted that more work is needed on the role of cyamids and disease spread (Overstreet et al. 2009). For the PCFG, there is an iconic whale we call “Scarlett” (also known as “Scarback”) who has a large scar on the right side of her back that is highly identifiable due to the orange swarm of cyamids that are constantly surrounding the edges of the wound. She has managed to survive and thrive, producing many calves over the years, but questions remain: How are the cyamids affecting the healing process? Are they increasing or decreasing the risk of infection? How does the frequency of large injuries like this on whales contribute to the cyamid population over evolutionary time?
Figure 3. Right side of PCFG icon, “Scarlett” showing her massive scar covered with orange aggregations of cyamids.
Because whales are complex, highly mobile, long-lived creatures with a constant population of cyamid hitchhikers their skin condition is likely representative of specific to life history, phylogeography, and demographic traits of individuals. While we know that cyamids generally eat sloughed or damaged skin on the whale, what this behavior and symbiosis means for each whale’s individual physiology can be highly complex. Through our high-resolution drone and lateral imagery of the same individuals over time paired with other data sources, such as body condition and prey availability, cyamid scores can offer key insights into how environmental stressors and foraging success affect individual and population-level whale health.
These tiny crustaceans, clinging to the folds and scars of their hosts, might seem like background noise in a study focused on body condition or foraging ecology—but they’re far from incidental. In my research, I’ve come to see cyamids as part of the bigger story:silent indicators of stress, recovery, movement, and resilience. By pairing imagery of PCFG gray whale skin with data on prey availability and environmental conditions, I’m working to understand how foraging success and anthropogenic stressors (such as vessel traffic and entanglements) manifest not just in a whale’s body condition, but in the skin itself. The presence, distribution, and density of cyamids may offer yet another layer of insight into how gray whales are coping with changing ocean conditions. It’s a reminder that even the smallest details, like a patch of whale lice, can help us ask bigger questions about the health, resilience, and future of these cetaceans.
References
Callahan, C.M., n.d. MOLECULAR SYSTEMATICS AND POPULATION GENETICS OF WHALE LICE (AMPHIPODA: CYAMIDAE) LIVING ON GRAY WHALE ISLANDS.
Lehnert, K., IJsseldijk, L.L., Uy, M.L., Boyi, J.O., van Schalkwijk, L., Tollenaar, E.A.P., Gröne, A., Wohlsein, P., Siebert, U., 2021. Whale lice (Isocyamus deltobranchium & Isocyamus delphinii; Cyamidae) prevalence in odontocetes off the German and Dutch coasts – morphological and molecular characterization and health implications. International Journal for Parasitology: Parasites and Wildlife 15, 22–30. https://doi.org/10.1016/j.ijppaw.2021.02.015
Leung, Y., 1976. Life cycle of cyamus scammoni (amphipoda: cyamidae), ectoparasite of gray whale, with a remark on the associated species. Scientific Reports of the Whales Research Institute 28, 153–160.
Overstreet, R.M., Jovonovich, J., Ma, H., 2009. Parasitic crustaceans as vectors of viruses, with an emphasis on three penaeid viruses. Integrative and Comparative Biology 49, 127–141. https://doi.org/10.1093/icb/icp033
Pettis, H.M., Rolland, R.M., Hamilton, P.K., Brault, S., Knowlton, A.R., Kraus, S.D., 2004. Visual health assessment of North Atlantic right whales (Eubalaena glacialis) using photographs. Can. J. Zool. 82, 8–19. https://doi.org/10.1139/z03-207
Pirotta, E., Schick, R.S., Hamilton, P.K., Harris, C.M., Hewitt, J., Knowlton, A.R., Kraus, S.D., Meyer-Gutbrod, E., Moore, M.J., Pettis, H.M., Photopoulou, T., Rolland, R.M., Tyack, P.L., Thomas, L., 2023. Estimating the effects of stressors on the health, survival and reproduction of a critically endangered, long-lived species. Oikos 2023, e09801. https://doi.org/10.1111/oik.09801
Raverty, S., Duignan, P., Greig, D., Huggins, J.L., Huntington, K.B., Garner, M., Calambokidis, J., Cottrell, P., Danil, K., D’Alessandro, D., Duffield, D., Flannery, M., Gulland, F.M., Halaska, B., Lambourn, D.M., Lehnhart, T., Urbán R., J., Rowles, T., Rice, J., Savage, K., Wilkinson, K., Greenman, J., Viezbicke, J., Cottrell, B., Goley, P.D., Martinez, M., Fauquier, D., 2024. Gray whale (Eschrichtius robustus) post-mortem findings from December 2018 through 2021 during the Unusual Mortality Event in the Eastern North Pacific. PLoS One 19, e0295861. https://doi.org/10.1371/journal.pone.0295861
Stimmelmayr, R., Gulland, F.M.D., 2020. Gray Whale (Eschrichtius robustus) Health and Disease: Review and Future Directions. Front. Mar. Sci. 7. https://doi.org/10.3389/fmars.2020.588820
Takeda, M., Ogino, M., n.d. Record of a Whale Louse, Cyamus scammoni Dall (Crustacea: Amphipoda: Cyamidae), from the Gray Whale Strayed into Tokyo Bay, the Pacific Coast of Japan.
Von Duyke, A.L., Stimmelmayr, R., Sheffield, G., Sformo, T., Suydam, R., Givens, G.H., George, J.C., 2016. Prevalence and Abundance of Cyamid “Whale Lice” (Cyamus ceti) on Subsistence Harvested Bowhead Whales (Balaena mysticetus). Arctic 69, 331–340.
Würsig, B., Thewissen, J.G.M., Kovacs, K.M., 2017. Encyclopedia of Marine Mammals. Elsevier Science & Technology, Chantilly, UNITED STATES.
Baleen whales must navigate a seemingly featureless world to locate the resources they need to survive. The task of finding prey to feed on in the vast seascapes relies on the use of several sensory modalities that operate at different scales (Torres 2017; Figure 1). For example, baleen whale vision is believed to be rather limited, with the ability to see objects about 10-100 meters away. Yet, baleen whale somatosensory perception of oceanographic stimuli is thought to be on the order of 100-1000s kilometers. This diversity in sensory ability has led scientists to believe that whales, in fact all animals, perceive cues and make decisions at several scales. As ecologists, we endeavor to understand why and when animals are found (or not found) in certain locations as this knowledge allows us to better manage and conserve animal populations. With this information we can aim to minimize potential anthropogenic disturbance and protect important resource areas, such as foraging or nursing grounds. In order to accomplish this goal, we ourselves must conduct studies and test hypotheses at several scales (Levin 1992; Hobbs 2003). As someone who tackles spatial foraging ecology questions, I am particularly interested in understanding whale behavior and movement in the context of feeding. Since accurately measuring predator and prey distribution at the same scales can be challenging, we often resort to environmental variables to serve as proxies for prey, whereby we look for correlations between environmental variables and whales to understand and predict the distribution of our population.
Figure 1. Schematic of hypothetical interchange of sensory modalities used by baleen whales to locate prey at variable scales. X-axis represents log distance to prey from micro (left) to macro (right). Y-axis represents the relative use of each sensory modality between 0 (no contribution) to 10 (highest contribution). Each line and color represent a different sensory modality. Figure taken and caption adapted from Torres 2017.
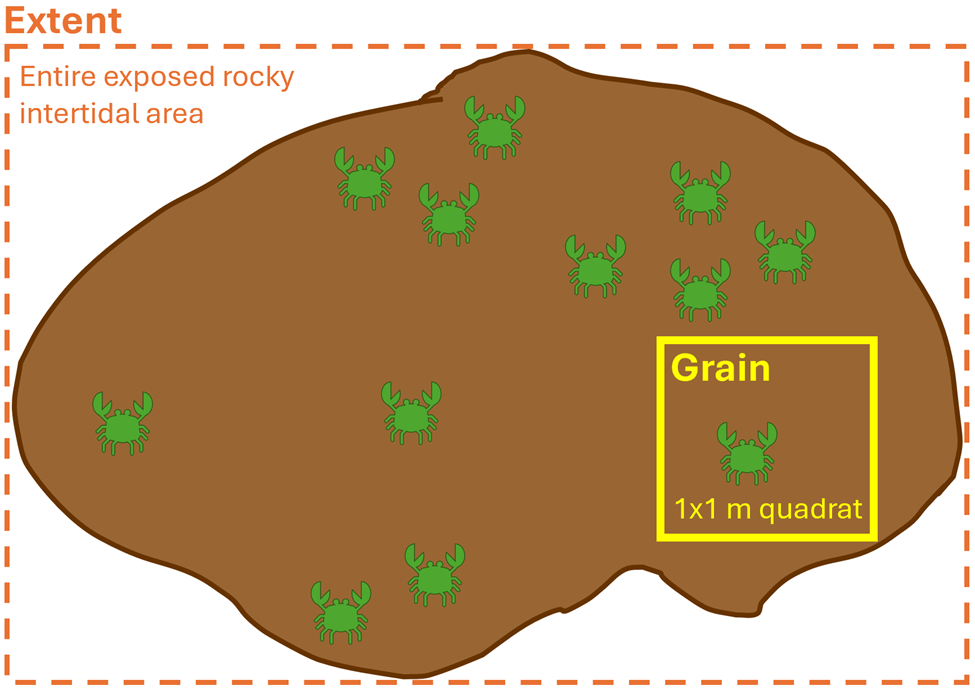
What do I mean when I use the word ‘scale’? The term scale is typically explained by two components: grain and extent (Wiens 1989). The grain is the finest resolution measured; in other words, how detailed we are measuring. The extent is the overall coverage of what we are measuring. These components can be applied to both spatial scale and temporal scale. For example, spatially, if we were using a 1×1 meter sampling quadrat to count the number of crabs on a rocky shore, then our grain would be the 1 m2 quadrat and the extent would be the entire exposed rocky intertidal area that we are surveying (Figure 2). Temporally, if we placed a temperature logger at the mouth of Yaquina Bay that took a temperature recording every minute for two years, then our grain would be one minute and the extent would be two years. So, when designing a study, it is imperative for us to decide on the spatiotemporal scales of the ecological questions we are asking and the hypotheses we are testing, as it will inform what data we need to collect. When making this decision, it is important to think about the scale at which the ecological process happens, as opposed to the scale at which we can observe the process (Levin 1992). In other words, we need to think from the perspective of our study species, as opposed to from our own human perspective. Making informed and ecologically reasonable decisions regarding the choice of scale relies on having prior knowledge of an animal’s biology, such as knowing that baleen whales might see a prey patch that is 50 meters away, but it may also somatosensorily perceive an oceanic front where zooplankton prey aggregate from 500 kilometers away.
Figure 2. Schematic of spatial scale where the extent (depicted by dashed orange box) is the entire exposed rocky intertidal area being surveyed and the grain (solid yellow box) is the 1×1 m quadrat being used to count crabs.
There is a wealth of studies that have explored space use patterns of wildlife relative to environmental variables to better understand foraging behavior. I want to share a couple from the marine mammal realm with you that I find particularly fascinating. In their 2018 study, González García and colleagues used opportunistic sightings of blue whales around the Azorean islands of Portugal and modeled their distribution patterns relative to physiographic and oceanographic variables summarized at different spatial (fine [1-10 km] and meso [10-100 km]) and temporal (daily, weekly, monthly) scales. The two variables that were most correlated with blue whale occurrence was distance from the coast and eddy kinetic energy (a measure of mesoscale variability of ocean dynamics). Both of these variables were interestingly found to be scale invariant, meaning that no matter which spatial and temporal scale was investigated, the relationship between blue whales and these two variables stayed the same; blue whale occurrence increased with increasing distance from the coast and was maximal at an eddy kinetic energy value of 0.007 cm2/s2 (Figure 3).
Figure 3. Functional response curves between presence of Azorean blue whales and distance to the coast (panel 1 on left) and eddy kinetic energy (panel 2 on right). The top row of each panel represents the low spatial scale and the bottom row represents the high spatial scale. Each column represents a different temporal scale (from left to right: daily, weekly, monthly). Note that the general shape of the relationship remains similar across all spatiotemporal scales and that the peak of the curves tend to occur at the same values for distance to coast and eddy kinetic energy across all scales. Figures taken from González García et al. 2018.
However, not all studies find scale invariant relationships. For example, Cotté and co-authors (2009) found that habitat use of Mediterranean fin whales was very much scale dependent. At a large scale (700-1,000 km and annual), fin whales were more densely aggregated during the summer in the Western Mediterranean where there was consistently colder water than in the winter. However, at a meso scale (20-100 km and weekly-monthly), fin whale densities were highest in areas where there were steep changes in temperature, as opposed to consistently cold temperatures. The authors explain that these differences in fin whale density and temperature at different scales are likely due to whale movement being driven by annually persistent prey abundance at the large scale, but at the meso scale, where prey aggregations are less predictable, the fin whales’ distribution becomes more driven by areas of physical ocean mixing.
As I investigate the environmental drivers of individual gray whale space use using our 8-year GRANITE (Gray whale Response to Ambient Noise Informed by Technology and Ecology) dataset, these studies (and many more) are at the top of my mind to interpret the patterns we are detecting. Our goal is to quantify and describe what environmental conditions (1) lead to a higher probability of a gray whale being seen in our central Oregon coast study area (~70 km) at a daily scale, and (2) influence space use patterns (activity range, residency, activity center) of different individual whales at annual scales. Our results show both consistency and variation in the environmental drivers of gray whales across these scales, leading me to deeply consider how gray whales make decisions at different points in their lives, based on information gained through various senses, to maximize their chances of capturing food. Previous work from the GEMM Lab on the relationships between gray whales and prey, at both fine (read more here) and large (read more here) scales have guided my work by providing specific hypotheses regarding environmental variables and lag times for me to test. Investigating the environmental drivers of animal space use and behavior is exciting work as it reveals that no single environmental variable determines animal distribution, but rather that multiple processes are happening concomitantly that animals respond to at different scales continually. It is only by studying animal space use patterns across spatiotemporal scales that we can begin to understand their complex decision-making patterns.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog below and get a monthly message when we post a new blog. Just add your name and email into the subscribe box below.
References
Cotté, C., Guinet, C., Taupier-Letage, I., Mate, B., & Petiau, E. (2009). Scale-dependent habitat use by a large free-ranging predator, the Mediterranean fin whale. Deep Sea Research Part I: Oceanographic Research Papers, 56(5), 801-811.
González García, L., Pierce, G. J., Autret, E., & Torres-Palenzuela, J. M. (2018). Multi-scale habitat preference analyses for Azorean blue whales. PLoS One, 13(9), e0201786.
Hobbs, N. T. (2003). Challenges and opportunities in integrating ecological knowledge across scales. Forest Ecology and Management, 181(1-2), 223-238.
Levin, S. A. (1992). The problem of pattern and scale in ecology: the Robert H. MacArthur award lecture. Ecology, 73(6), 1943-1967.
Torres, L. G. (2017). A sense of scale: Foraging cetaceans’ use of scale‐dependent multimodal sensory systems. Marine Mammal Science, 33(4), 1170-1193.
Wiens, J. A. (1989). Spatial scaling in ecology. Functional ecology, 3(4), 385-397.
As the sun set on February 16th, the R/V Star Keys pulled into Wellington Harbour, marking the end of the 2025 SAPPHIRE field season. The crew and science team returned to shore after a packed, productive, and successful three weeks at sea studying the impacts of environmental change on blue whales and krill in the South Taranaki Bight, Aotearoa New Zealand.
A blue whale comes up for air in the South Taranaki Bight.
In stark contrast to the 2024 field season, which featured dense and seemingly endless layers of gelatinous salps in the water and no krill or blue whales in the South Taranaki Bight, the 2025 field season was filled with blue whales and krill. In our three weeks aboard our research vessel Star Keys this year, we observed 66 blue whales, most of which were lunge feeding at the surface on dense patches of krill. We also collected krill for on-board respiration experiments and to be frozen to measure their lengths, weights, and caloric content. We recovered two hydrophones that recorded blue whale calls for the past year, and replaced them with two more. We collected identification photos, skin and blubber tissue samples for genetic and hormone analysis, and flew drones over almost all whales we encountered to measure body condition and morphology. We conducted water column profiles to measure the oceanography of the region, and mapped the prey field as we surveyed using a scientific echosounder.
Map of our survey effort (gray tracklines), blue whale sightings (red circles), and hydrophone locations (purple stars).
Around the world, we are currently bearing witness to environmental change. Our survey last year in 2024 was a reminder of the challenges these blue whales face to survive and thrive in an increasingly unpredictable ocean. This year was a poignant example of the vibrant marine life that exists here in the South Taranaki Bight when ocean conditions align more closely with what is expected, and of the incredible resilience of these animals as they navigate changing waters. These contrasting conditions over multiple years are key to our understanding as we study the impacts of climate change on krill and blue whales through the SAPPHIRE project.
Drone image of a blue whale coming to the surface.
The fieldwork we do to collect these data is motivated by scientific questions, management needs, and fascination with this ecosystem. But ultimately, what makes fieldwork possible and memorable is the people. We are deeply grateful for the many partners on the SAPPHIRE project. The 2025 science team was made up of Leigh Torres, Dawn Barlow, KC Bierlich, Kim Bernard (Oregon State University), Mike Ogle, and Ros Cole (Department of Conservation). The outstanding crew of the R/V Star Keys (Western Work Boats), Josh Fowden, Dave Futter, and Jordy Maiden-Drum, kept us safe, sailing, fed, and happy for three intense weeks. We are also grateful for our shore support, including our colleagues at Cornell University’s Yang Center for Conservation Bioacoustics, NIWA, the Marine Mammal Institute at Oregon State University, and the University of Auckland. Importantly, we deeply appreciate our many stakeholders who help us share, learn, and make our findings meaningful, including the Department of Conservation, the people of Aotearoa, and iwi across our study region, especially Ngāruahine who hosted us at the Rangatapu Marae for a profound hui with a powerful pōwhiri and critical wānanga of knowledge sharing.
Drone image of a blue whale mom and calf pair.
Now the next phase of the work begins. We have many terabytes of data to process, analyze, interpret, and share. We will certainly have our hands full. But while we are at our computers back in Oregon, we will be holding the memories of this field season close: The brilliant turquoise glow of a blue whale just below the surface, the sound of the deep exhalation as the whale comes up for air, and the awe of looking into a blue whale’s eye as it engulfs a dense swarm of krill; The golden sunset lighting and moon rise over Cape Farewell, and Mount Taranaki towering over the blue waters of the South Taranaki Bight; The giddy exclamations or silent awe of those of us privileged to spend time in these waters observing these animals, and the visions that linger just behind our eyelids as we fell into an exhausted sleep. We will see what the next year holds for the SAPPHIRE team and the blue whales and krill of the South Taranaki Bight.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly alert when we make a new post! Just add your name into the subscribe box below!
By Rachel Kaplan, PhD candidate, Oregon State University College of Earth, Ocean, and Atmospheric Sciences and Department of Fisheries, Wildlife, and Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
What does a whale look for at mealtime? Is it a lot of food, its quality, or the type of food? An improved understanding of what makes krill swarms, an important prey item, appetizing for humpback whales can help us anticipate where and when we will see them in our ocean backyard, the Northern California Current (NCC) foraging grounds. In a new paper, we found that humpback whale presence in the NCC is tied to several different metrics of krill swarm quality and quantity, particularly species composition (what types of krill are in the swarm), energetic density (the caloric richness of the average mouthful), and biomass (how much krill is in the swarm). Interestingly, relationships between humpback whales and these krill swarm quality metrics are variable in time and space, dependent on whether the whale is foraging on or off the continental shelf and if it is early or late in the foraging season.
This study required a special, fine-scale dataset of simultaneous observations of krill and whales at sea. While GEMM Lab members conducted marine mammal surveys, we simultaneously observed the prey that whales had access to, using active acoustics (essentially a fancy fish finder) to profile the water column and net tows to collect krill. When we put all these data streams together, we found that increases in biomass, energetic density, and the amount of a particular species, Thysanoessa spinifera, in a krill swarm were positively related to humpback whale presence. These results suggest that humpback whales balance multiple prey quality factors to select feeding areas that offer both plentiful and high-quality krill.
Figure 1. Top photo: Marine mammal observers Clara Bird (left) and Dawn Barlow (right) collect humpback whale distribution data. Bottom photo: At the same time, Talia Davis (left) and Rachel Kaplan (right) collect krill samples.
Species composition
Euphausia pacifica and T. spinifera are the two most common krill species in the NCC region, and other research has shown that many krill foragers, including blue whales, seabirds, and fish, preferentially consume T. spinifera. Although this pickiness is well-warranted – individual T. spinifera tend to be larger than E. pacifica and much higher in calories during the late foraging season – targeting this juicy prey item could place humpback whales in competition with these other species, which may make it harder for them to find a square meal. Nevertheless, we found positive relationships between the proportion of T. spinifera in a krill swarm and humpback whale presence, suggesting humpback whales do in fact preferentially prey upon T. spinifera, particularly during the late foraging season (about July-November).
Energetic density
Humpback whales’ preference for T. spinifera during the late foraging season may be due to its higher caloric content. Although the two krill species offer a similar number of calories early in the foraging season,we found that the energetic density of T. spinifera was elevated during the late foraging season, after productive upwelling conditions have revved up the food web over several months. Krill swarm energetic density had a positive effect on humpback whale occurrence, particularly in the late season when T. spinifera and E. pacifica have significantly different caloric contents. Interestingly, this positive relationship was not present onshore during the early season, when the two krill species have similar caloric contents.
Figure 2. In terms of caloric content, Thysanoessa spinifera krill like this one are the winners in the NCC region! They pack on the milligrams through the productive summer season, making them advantageous prey for hungry whales.
Humpback whales also target forage fish on the continental shelf that have higher energetic densities than krill, indicating that whales may selectively forage on fish – even though it is more energetically expensive to capture them. Variation in seasonal and spatial relationships with krill swarm energetic density may explain why humpback whales prey-switch, selecting prey based on availability and quality. As flexible foragers, humpback whales can consistently target higher-quality swarms that offer more energy per lunge.
Biomass
Biomass, or the total amount of krill in a swarm, was the single best predictor of humpback whale presence that we tested. This result emphasizes the importance of large krill swarms in explaining where humpback whales forage. We found that krill swarm biomass tended to be higher offshore, where swarms were also located deeper in the water column. During the late season offshore, krill quality (elevated due to higher late season caloric contents) together with quantity (higher offshore biomass) may make these offshore swarms the most favorable for foraging whales, despite being deeper.
Figure 3. When humpback whales “fluke,” as seen in this picture, it may indicate the beginning of a foraging dive to capture prey.
Future food webs
Environmental conditions are changing in the NCC, with events like marine heatwaves and strong El Niño events shifting food webs. E. pacifica and T. spinifera may respond to climate change differently based on their life history strategies. Distributional shifts, such as the disappearance of T. spinifera from the NCC during the 2014–2015 “Blob” marine heatwave that transformed the northeast Pacific Ocean, could diminish or entirely remove this key prey item. As a result of such climate and environmental changes, humpback whales may encounter lower quality prey and/or shifts in prey distribution that could make it harder for them to find a meal. In changing oceans, better understanding krill prey quality for humpback whales will shape improved tools for conservation management.
References
Chenoweth, E., Boswell, K., Friedlaender, A., McPhee, M., Burrows, J., Heintz, R., and Straley, J. 2021. Confronting assumptions about prey selection by lunge‐feeding whales using a process‐based model. Funct. Ecol., 35.
Croll, D., Marinovic, B., Benson, S., Chavez, F., Black, N., Ternullo, R., and Tershy, B. 2005. From wind to whales: trophic links in a coastal upwelling system. Mar. Ecol. Prog. Ser., 289: 117–130.
Derville, S., Buell, T. V., Corbett, K. C., Hayslip, C., and Torres, L. G. 2023. Exposure of whales to entanglement risk in Dungeness crab fishing gear in Oregon, USA, reveals distinctive spatio-temporal and climatic patterns. Biol. Conserv., 281: 109989.
Fiedler, P. C., Reilly, S. B., Hewitt, R. P., Demer, D., Philbrick, V. A., Smith, S., Armstrong, W., et al. 1998. Blue whale habitat and prey in the California Channel Islands. Deep Sea Res. Part II, 45: 1781–1801.
Fisher, J. L., Menkel, J., Copeman, L., Shaw, C. T., Feinberg, L. R., and Peterson, W. T. 2020. Comparison of condition metrics and lipid content between Euphausia pacifica and Thysanoessa spinifera in the northern California Current, USA. Prog. Oceanogr., 188.
Murdoch, W. W. 1969. Switching in General Predators: Experiments on Predator Specificity and Stability of Prey Populations. Ecol. Monog., 39: 335–354.
Nickels, C. F., Sala, L. M., and Ohman, M. D. 2018. The morphology of euphausiid mandibles used to assess selective predation by blue whales in the southern sector of the California Current System. J. Crustacean Biol., 38: 563–573.
Price, S. E., Savoca, M. S., Kumar, M., Czapanskiy, M. F., McDermott, D., Litvin, S. Y., Cade, D. E., et al. 2024. Energy densities of key prey species in the California Current Ecosystem. Front. Mar. Sci., 10: 1345525.
Robertson, R. R., and Bjorkstedt, E. P. 2020. Climate-driven variability in Euphausia pacifica size distributions off northern California. Prog. Oceanogr., 188.
Santora, J. A., Mantua, N. J., Schroeder, I. D., Field, J. C., Hazen, E. L., Bograd, S. J., Sydeman, W. J., et al. 2020. Habitat compression and ecosystem shifts as potential links between marine heatwave and record whale entanglements. Nat Commun, 11: 536.
Spitz, J., Trites, A. W., Becquet, V., Brind’Amour, A., Cherel, Y., Galois, R., and Ridoux, V. 2012. Cost of Living Dictates what Whales, Dolphins and Porpoises Eat: The Importance of Prey Quality on Predator Foraging Strategies. PLoS ONE, 7: e50096.
Tanasichuk, R. 1998a. Interannual variations in the population biology and productivity of Thysanoessa spinifera in Barkley Sound, Canada, with special reference to the 1992 and 1993 warm ocean years. Mar. Ecol. Prog. Ser., 173: 181–195.
Videsen, S. K. A., Simon, M., Christiansen, F., Friedlaender, A., Goldbogen, J., Malte, H., Segre, P., et al. 2023. Cheap gulp foraging of a giga-predator enables efficient exploitation of sparse prey. Sci. Adv., 9: eade3889.
Weber, E. D., Auth, T. D., Baumann-Pickering, S., Baumgartner, T. R., Bjorkstedt, E. P., Bograd, S. J., Burke, B. J., et al. 2021. State of the California Current 2019–2020: Back to the Future With Marine Heatwaves? Front. Mar. Sci., 8.
By Lindsay Wickman, Postdoctoral Scholar, Oregon State University Department of Fisheries, Wildlife, and Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
Earlier this month, most of the GEMM Lab and I attended the 25th Biennial Conference on the Biology of Marine Mammals in Perth, Western Australia. This year’s theme, “Fishing for Change,” acknowledged that incidental entanglement in fishing gear is currently the most pervasive threat to marine mammals (e.g., Avila et al., 2018). While many presentations on the prevalence and impacts of entanglement on marine mammals were sobering, it was also inspiring to be surrounded by so many dedicated people working to address this urgent issue. For me, one of the most memorable anecdotes was an incredible whale disentanglement story shared by Paul Cottrell, a Marine Mammal Coordinator at the Department of Fisheries and Oceans Canada (DFO) in British Columbia.
An Incredible Story of Whale Disentanglement
During August 2024, DFO and a local NGO (Straitwatch) responded to a report of two humpback whales entangled in the same fishing gear near Quadra Island, B.C., Canada. Their photo-identification histories revealed that one whale had migrated from Hawaii, while the other had come from Mexico. Now tied together, the two whales’ fates became intertwined, forcing them to coordinate their movements. This situation obviously raised concerns about their welfare and survival, but I also had to silently wonder, “Did the two whales ever argue about where to migrate next? Would they choose Hawaii or Mexico?”
Thanks to the rescuers’ efforts, both whales were freed and able to make their own choice about where to spend the breeding season. As Paul explained, successfully disentangling one whale is challenging and dangerous, so freeing two was an impressive feat. After the rescue, a video showed the whales continuing to swim together synchronously, as if they did not realize they were no longer connected!
Most Entangled Whales are Out of Sight
The story above exemplifies a “confirmed” entanglement—these whales were seen dragging fishing gear and the event was reported by concerned citizens. However, most entanglement events are never witnessed, for several reasons.
When a whale becomes entangled in fishing gear, it rarely remains anchored in place. Instead, the whale often breaks part of the gear, dragging it behind as it swims. The likelihood of observing the entangled whale subsequently depends on both the chance of it being seen and the observer’s awareness and willingness to report the event (Robbins and Mattila, 2004).
An entangled humpback whale drags gear off of San Diego, California. Credit: Keith Yip, taken under NOAA Permit #18786.
Once entangled, many become a “dead whale swimming,” eventually succumbing to starvation and/or infections (Dolman and Moore, 2017). Many entanglements involve the mouth, severely impacting the whale’s ability to feed (Moore and van der Hoop, 2012). The additional drag imposed by entanglement is comparable to the energetic costs of migration or reproduction, causing a significant depletion in their energy reserves (van der Hoop et al. 2015). Serious injuries include amputations, hemorrhage, and infections (Cassoff et al., 2011).
Although some carcasses of entangled whales wash ashore, most are lost at sea and never recovered. For example, even with relatively intensive monitoring for North Atlantic right whale (NARW) carcasses, Pace et al. (2021) estimated that recovered carcasses represented just 36% of the total deaths. These recovered carcasses may also underestimate the toll of entanglement; entanglement accounted for 51% of mortality in the carcasses vs. 87% of serious injuries observed in living NARWs (Pace et al., 2021).
For whales that manage to dislodge the gear and survive, scars can provide clues to their past entanglement history. Injuries from the fishing lines can leave indentations where they cut through skin and blubber, and healed wounds often result in white pigmented scars that wrap around the body (especially the flukes and peduncle; e.g., Robbins and Matilla 2004). The widespread prevalence of these scars suggests that in many cases, whales can actually dislodge the gear on their own. For example, a study of entanglement scars on humpback whales in the Gulf of Maine revealed that 10% of adults and 30% of juveniles acquired new entanglement scars between 2009-2010. Without scarring analyses though, most of these entanglements would have been missed; just 7% of these individuals with entanglement-related scars were seen while entangled (Robbins 2012).
A humpback whale fluke and peduncle showing scarring likely caused by a past entanglement. Credit: GEMM lab, taken under NOAA Permit # 27426 issued to MMI.
Unfortunately, scars are not the only long-term consequence of non-lethal entanglement events. Previously entangled NARWs have lower survival rates than unaffected individuals (Robbins et al. 2015, Reed et al. 2024), and long-term stress responses can impact their future health and reproductive success (Pettis et al. 2004). It is tempting to assume that only severe entanglements affect future reproduction and survival, but a lack of extensive external injuries doesn’t necessarily mean that the impact of the entanglement event is more minor (Robbins and Matilla, 2004). For example, Reed et al. (2024) found that NARWs with entanglement injuries classified as minor were less likely to transition from a “non-breeder” to “breeder” status than those with severe injuries.
Tracking Unseen Entanglements: Project SLATE
Since reported entanglements and recovered carcasses reveal just a fraction of actual entanglements, researchers are continuing to innovate ways of documenting these “unseen” entanglement events.
As discussed in a previous blog post, photos of entanglement scars on the flukes and peduncles of humpback whales are being utilized in Project SLATE to detect trends in entanglement off the coast of Oregon, USA. Analyzing images of whales for signs of past entanglements is a meticulous process that may not seem as thrilling as responding to an actual disentanglement event. However, in areas with lower population densities, such as the Oregon coast, reported entanglements are undoubtedly an underestimate of the true number of events. Thus, tracking scarring rates can provide more comprehensive data on entanglement prevalence in Oregon than confirmed reports alone.
What to do if you see an entangled whale
If you happen to observe an entangled whale, please do not attempt to disentangle it yourself. Whale disentanglement is dangerous and complex, so best left to the experts! When well-meaning citizens attempt a disentanglement on their own, it can also result in an “incomplete disentanglement,” where some, but not all, gear is removed from the whale. Incomplete disentanglements just make it harder for responders to subsequently find and successfully rescue the whale.
Instead, report the entanglement by promptly calling:
Entanglement Reporting Hotline: 1-877-SOS-WHAL or 1-877-767-9425
or U.S. Coast Guard: VHF Ch. 16
Videos or photos showing the entangling gear is very helpful to trained responders, but remember to stay at least 100 yards from the whale, and beware of snagging your vessel in the lines. Visit NOAA Fisheries for more information.
A NOAA-led team disentangle a humpback whale near Dutch Harbor, Alaska. Credit: Andy Dietrick/NOAA, taken under NOAA Permit #18786.
References:
Avila, I.C., Kaschner, K., Dormann, C.F.. (2018). Current global risks to marine mammals: Taking stock of the threats. Biol. Conserv. 221, 44–58.
Dolman, Sarah J., and Michael J. Moore. (2024). Chapter 4: Welfare implications of cetacean bycatch and entanglements. In A. Butterworth (Ed.) Marine Mammal Welfare: Human Induced Change in the Marine Environment and Its Impacts on Marine Mammal Welfare (pp. 41-65).
Moore, M. J., and van der Hoop, J. M. (2012). The painful side of trap and fixed net fisheries: chronic entanglement of large whales. Journal of Marine Sciences, 2012.
Pace III, R. M., Williams, R., Kraus, S. D., Knowlton, A. R., & Pettis, H. M. (2021). Cryptic mortality of North Atlantic right whales. Conservation Science and Practice, 3(2), e346
Reed, J., New, L., Corkeron, P., Harcourt, R. (2024). Disentangling the influence of entanglement on recruitment in North Atlantic right whales. Proc. R. Soc. B Biol. Sci. 291.
Robbins, J., Mattila, D. (2004). Estimating humpback whale (Megaptera novaeangliae) entanglement rates on the basis of scar evidence. Rep. to Northeast Fish. Sci. Center, Natl. Mar. Fish. Serv. 43EANF030121 22p.
Robbins, J. (2012). Scar-Based Inference Into Gulf of Maine Humpback Whale Entanglement : 2010. Report to the Northeast Fisheries Science Center National Marine Fisheries Service, EA133F09CN0253 Item 0003AB, Task 3.
van der Hoop JM, Corkeron P, Kenney J, Landry S, Morin D, Smith J, Moore MJ. (2015). Drag from fishing gear entangling North Atlantic right whales. Mar Mamm Sci 32(2):619–642.
By Nicole Principe, first-year PhD student, OSU Dept of Fisheries, Wildlife and Conservation Sciences, GEMM Lab
Humans rely on oceans and coastal ecosystems for a variety of resources, such as tourism and recreation, fishing and aquaculture, transport of goods, and resource extraction. However, each use is contributing to new and cumulative stressors that are impacting marine mammals. The health of marine mammal populations can often serve as indicators of overall environmental health. Therefore, studying the stressors they face can help provide insights into the broader impacts on marine ecosystems and determine if conservation or management measures are necessary. As a master’s student at the College of Charleston in South Carolina and subsequently the stranding and research technician with the Lowcountry Marine Mammal Network (LMMN), I saw first-hand how some of these stressors affect local marine mammal populations.
In my role as the stranding and research technician with LMMN, I led the response and recovery of all deceased marine mammals, mainly bottlenose dolphins (Tursiops erebennus), in South Carolina to determine cause of death and identify main sources of mortality. Threats to these cetaceans can be environmental or anthropogenic in origin. Carefully examining and sampling every individual during a necropsy was critical to determine the presence of infectious disease, the contaminant and microplastic load, and any sign human interaction. While deaths from environmental causes can be more challenging for humans to mitigate, direct threats from human activity can be lessened with conservation actions and increased education to the public. LMMN responds to several strandings of dolphins each year that are the result of entanglement or boat strike. South Carolina has one of the highest rates of crab pot entanglements. In some cases, the call came quick enough that a disentanglement was possible, but in others, we found the animal already deceased with rope and gear still attached. Hundreds, if not thousands, of commercial and recreational crab pots are deployed within South Carolina estuaries, yet there are currently no regulations in place to help mitigate the threat of entanglement.
Images: Conducting a necropsy in the lab (left) and doing a preliminary examination in the field (right) on stranded bottlenose dolphins. Photo credit: Lowcountry Marine Mammal Network
LMMN also conducts land and boat-based surveys to better understand strand feeding, which is a unique foraging strategy utilized by a small number of dolphins in South Carolina. When dolphins strand feed, they herd and trap fish up onto mudbanks or shorelines. The dolphins chase after the fish, briefly stranding themselves as they try to catch them. It is an incredible behavior to witness and because of this, it has become highly publicized as a tourist activity. There are areas where the public can walk right up as dolphins are attempting to hunt and many instances of people trying to touch, feed, or otherwise harass the dolphins have been reported. I also conducted a small study where I used drones to identify human interferences towards dolphins strand feeding and found that boaters and kayakers were often approaching the animals too closely, following them, or speeding through the inlet when animals were present. The write up on that project can be found here. High levels of human disturbance towards dolphins strand feeding could lead individuals to abandon otherwise suitable habitat, causing them to expend more energy to look for food elsewhere.
Images: Bottlenose dolphins strand feeding (left) and an instance of a beachgoer illegally harassing dolphins while they hunt (right). Photo credit: Glen Cox & Lucy Grierson.
To help mitigate threats to dolphins from entanglements, boat strikes, and illegal harassment, the LMMN team and I created an educational workshop called W.A.V.E., which stands for Wildlife Awareness and Viewing Etiquette. These half-day workshops are tailored to both recreational boaters/public and commercial tour operators and fishermen and cover topics ranging from the importance of marine mammals in our ecosystem, the Marine Mammal Protection Act, global and local threats, and ways we can view marine wildlife that reduce disturbance. It is my hope that with more education and awareness about how humans use our waterways and interact with wildlife in negative ways, it can lead to positive changes. For more information about LMMN’s W.A.V.E. Workshops, head to their website.
Image: Successful W.A.V.E. Workshop with local eco-tour operators. Photo credit: Lowcountry Marine Mammal Network
In addition to cumulative stressors from human interactions, I also began to contemplate the role of climate change as a threat to the lives of marine mammals during my master’s research on dolphin distribution within the Charleston Estuary System (CES). A main question I was investigating was if and why some dolphins travel into low salinity waters high in the estuarine system. Bottlenose dolphins have evolved in marine and estuarine environments where salinity levels are typically ~30 parts per thousand (ppt). While dolphins can withstand short durations of exposure to low salinity (defined as 15 ppt), prolonged exposure to freshwater can result in negative health consequences, such as sloughing of skin and ulcerative lesions, changes in pathophysiology, and eventual mortality (Ewing et al., 2017). Over the past 20 years, many intermittent dolphin sightings and strandings occurred in riverine areas of the CES where salinity levels were below 10 ppt. To better understand how and why dolphins use this risky habitat, I conducted drone surveys across the CES for a year. I did find dolphin groups traveling and feeding in low salinity waters, however, the encounters were only during months with warmer water temperatures (Principe et al., 2023). We hypothesize that environmental conditions during those months may lead to decreased prey availability in the lower, more suitable parts of the estuary, forcing dolphins to travel further up the rivers to access higher abundances of prey (especially mullet). Other studies in different regions have found similar results of dolphins traveling into low salinity water during warmer months potentially in response to prey (Mintzer and Fazioli, 2021; Takeshita et al., 2021).
Images: Drone snapshot of six bottlenose dolphins traveling together (left) and a dolphin observed with skin lesions in low salinity habitat (right). Photo credit: Nicole Principe, NMFS Permit 21938
These results lead to questions as to how prey and dolphin movements will shift under future climate change scenarios. Increasing warm water temperatures may lead to further shifts in prey distribution, potentially driving more estuarine dolphins to utilize upper riverine habitats to find food. Just since 2022, four dolphins were observed in freshwater habitat for several weeks. Two were eventually found and confirmed deceased and two went missing and are presumed deceased. If more dolphins use and remain in these low salinity habitats for extended periods, negative health consequences could lead to population impacts and signal a need for more conservation and management actions.
It is quickly becoming evident that climate change is threatening marine mammals, at both local and global scales. More research is needed to better understand how changing environmental conditions is impacting the availability and quality of prey and how large marine predators are shifting in response. For my PhD, I am working with the GEMM Lab on the SAPPHIRE (Synthesis of Acoustics, Physiology, Prey, and Habitat in a Rapidly changing Environment) project, where we are researching how changing ocean conditions affect the availability of krill, and blue whale behavior, health, and reproduction in New Zealand. The South Taranaki Bight (STB) region experiences a productive coastal upwelling system that supports enhanced primary productivity (Chiswell et al. 2017) and dense aggregations of prey (Bradford-Grieve et al., 1993). Pygmy blue whales (Balaenoptera musculus brevicauda) in this region are not known to migrate and instead use the STB region year-round for foraging and reproduction (Torres, 2013; Barlow et al., 2022). After a marine heatwave in the Tasman Sea in 2015-2016, there were less krill aggregations due to lessened upwelling (Barlow et al., 2020), which caused reduced foraging effort, and subsequently reduced reproductive activity by blue whales (Barlow et al. 2023). Continued field work and data analysis will help us to develop Species Health Models that will predict how these prey and predator populations will respond to future environmental change.
Overall, it is clear that human activity is leading to direct and indirect impacts on marine mammal populations at many different scales, from an individual human harassing a foraging dolphin to global climate change impacts on blue whale population dynamics. Ongoing research is essential in understanding these impacts better and thus inform development of effective conservation strategies to protect both marine mammals and the environment.
References
Barlow DR, Bernard KS, Escobar-Flores P, Palacios DM, Torres LG (2020) Links in the trophic chain: Modeling functional relationships between in situ oceanography, krill, and blue whale distribution under different oceanographic regimes. Mar Ecol Prog Ser 642:207–225.
Barlow DR, Klinck H, Ponirakis D, Branch TA, Torres LG (2023) Environmental conditions and marine heatwaves influence blue whale foraging and reproductive effort. Ecol Evol 13:e9770.
Barlow DR, Klinck H, Ponirakis D, Holt Colberg M, Torres LG (2022) Temporal occurrence of three blue whale populations in New Zealand waters from passive acoustic monitoring. J Mammal 104(1): 29–38.
Bradford-Grieve JM, Murdoch RC, Chapman BE (1993) Composition of macrozooplankton assemblages associated with the formation and decay of pulses within an upwelling plume in greater cook strait, New Zealand. New Zeal J Mar Freshw Res 27(1): 1–22.
Chiswell SM, Zeldis JR, Hadfield MG, Pinkerton MH (2017) Wind-driven upwelling and surface chlorophyll blooms in greater Cook Strait. New Zeal J Mar Fresw Res 51(4): 465–489.
Ewing RY, Mase-Guthrie B, McFee W, Townsend F, Manire CA, Walsh M,
Borkowski R, Bossart GD, Schaefer AM (2017). Evaluation of serum for pathophysiological effects of prolonged low salinity water exposure in displaced bottlenose dolphins (Tursiops truncatus). Front Vet Sci 4
Hornsby F, McDonald T, Balmer BC, Speakman T, Mullin K, Rosel P, Wells R, Telander A, Marcy P, Schwacke L (2017) Using salinity to identify common bottlenose dolphin habitat in Barataria Bay, Louisiana, USA. Endanger Species Res 33: 833–192.
Mintzer VJ, Fazioli KL (2021) Salinity and water temperature as predictors of bottlenose dolphin (Tursiops truncatus) encounter rates in upper Galveston Bay, Texas. Front Mar Sci 8
Principe N, McFee W, Levine N, Balmer B, Ballenger J (2023). Using Unoccupied Aerial Systems (UAS) to Determine the Distribution Patterns of Tamanend’s Bottlenose Dolphins (Tursiops erebennus) across Varying Salinities in Charleston, South Carolina. Drones 7(12): 10.3390/drones7120689.
Takeshita R, Balmer BC, Messina F, Zolman ES, Thomas L, Wells RS, Smith CR, Rowles TK, Schwacke LH (2021). High site-fidelity in common bottlenose dolphins despite low salinity exposure and associated indicators of compromised health. PLoS ONE, 16(9), e0258031.
Torres LG (2013) Evidence for an unrecognised blue whale foraging ground in New Zealand. New Zeal J Mar Freshw Res 47:235–248.
By Natalie Nickells, visiting PhD Student, British Antarctic Survey
For the last three months, I’ve been lucky enough to be welcomed into the GEMM lab as a visiting PhD student to work on the acoustic data from hydrophones in CATS tags deployed on gray whales. This work has been a huge change for me! I’ve gone from studying Antarctic baleen whale foraging, the topic of my PhD, from a distance at my desk in Cambridge England, to studying PCFG gray whales in Newport- and finally being in the same country, state, and even county to the whales I am studying! Unlike my Antarctic research, where whale blows in the distance become tiny points in a sea of data, listening to the CATS tag data has allowed me to really connect with these animals on an emotional level, as I’ve spent days, weeks and months listening to the world as they hear it.
Humans are fundamentally visual creatures- we take in information through sight first, with hearing probably our second, or for some even third, sense in line. However, for marine mammals, the same cannot be said: their world is auditory first. This fact is an important realisation to get our heads around, highlighted beautifully by the phrase “the ears are the window to the soul of the whale” (Sonic Sea (2017)) or Tim Donaghy’s emotive statement that “a deaf whale is a dead whale”. High levels of ocean noise therefore have a huge impact on baleen whales. Imagine trying to do your groceries or find a friend while blindfolded or in a thick fog– you might struggle to access food or communicate with others, and your stress would certainly be high. To succeed, you would likely need to change your behaviour.
Behavioural changes in response to ocean noise are observed in baleen whales: for example, humpback whales change their foraging behaviour when ship noise increases (Blair et al., 2016), and gray whales have been shown to call more frequently and possibly more loudly in conditions of high ocean noise (Dahlheim & Castellote, 2016). However, even in the absence of notable behaviour change due to ocean noise, North Atlantic right whales may still be experiencing a stress response. When shipping traffic in the Bay of Fundy significantly decreased in the aftermath of 9/11, North Atlantic right whales in the area had decreased chronic stress levels (Rolland et al., 2012).
Previous work by the GEMM lab observed this stress response to ocean noise in gray whales. They found a correlation between high levels of glucocorticoid (a stress indicator) in male gray whale faeces with high vessel noise and vessel counts in the area. Vessel noise was measured using two static hydrophones off the Oregon coast, and it was assumed all animals in the area experienced the same noise (Lemos et al., 2022; Pirotta et al., 2023). However, a static hydrophone is an imperfect measure of the sound levels a mobile animal experiences, particularly as we might expect animals to change behaviour when disturbed (Sullivan & Torres, 2018). This previous work became the starting point for the question I have addressed during my time in the GEMM Lab: can we measure and characterise the sound levels an individual whale was exposed to? Enter CATS tags. These are suction-cup tags fitted with a host of sensors, which have been used by the GEMM lab since 2021 (see Image 1). So far, they have mostly been used for their accelerometry data (Colson et al. (in press), see also Kate’s blog post). However, the GEMM lab had the foresight to put hydrophones on these tags, and as a result I was welcomed into the lab by a bumper-crop of hydrophone data just waiting to be analysed!
Image 1: A gray whale (“Slush”) being tagged with a CATS tag and Natalie (right) with the same tag.
This tag data is particularly valuable, not only for its ability to follow the acoustic world of an individual whale, but also due to the whole suite of data that comes with the acoustics: essentially, the acoustic data comes with behavioural data. Or at least, it comes with data from which we can infer behaviour (Colson et al, in press)! Incorporating behaviour into passive acoustics work hugely strengthens its ecological usefulness (Oestreich et al., 2024). We can hear what an individual whale is hearing, and we can also infer what they were doing before, during, and after they heard or made that sound. Having behavioural data also means that we can ground-truth the sounds we hear. When hearing an interesting sound, I can go back to the video data and accelerometer data to check what the whale sees, what its body-position is doing (e.g., is it headstand foraging?) and the speed and direction of its travel. Context is key!
The importance of context was highlighted in my very first week here in the GEMM lab. I became very interested in a sound I could hear frequently when the whale would surface- a distorted bark-like noise, but the whale was surely too far offshore for any barking dog to be heard? And almost every time the whale surfaced? After a few days pondering, I shared my mystery with Leigh, who laughingly revealed that one of the whale-watching boats in this area has a ‘whale-alerting’ dog on board! Sometimes if it sounds like a dog… it’s a dog! Besides my slightly anticlimactic discovery of dogs barking, committing time to listening to the tags and hearing what the whales hear, has been a magical experience. My favourite hydrophone sound, that still gets me excited when I hear it, is the gray whale ‘bongo call’- or as it’s more formally known in the literature, M1 vocalisation (Guazzo et al., 2019). I’ll let you decide which name is more appropriate! I first heard this call when investigating a time on “Scarlett’s” tag when we knew her 14 year-old daughter “Pacman” had been close: about 15 minutes before “Pacman” appears on the video, Scarlett makes this call (you can play the clip below to listen). In “Lunita’s” tag, we even hear this call three times in a row!
Image 2: A ‘bongo call’ made by “Scarlett” when her daughter “Pacman” was nearby.
Relatively little research has been done on gray whale calls compared to other more studied species like humpbacks. Most of this research has taken place on gray whale migratory routes (Guazzo et al., 2019, 2017; Burnham et al. 2018) or in captivity (Fish et. al, 1974 ) so these tag recordings could be a valuable addition to a small sample from the foraging grounds (Clayton et al., 2023; Haver et al., 2023)- as well as being very personally exciting to hear!
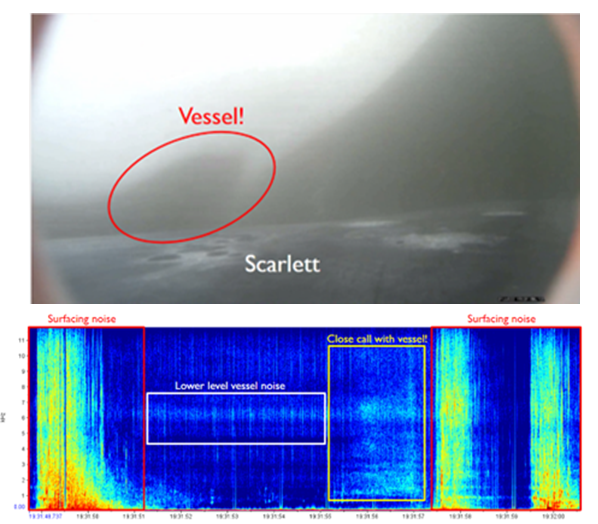
We’ve also been able to use the tag hydrophone data to look at close calls with ships. As I was going through the data on “Scarlett’s” tag, I noticed a spike in vessel noise. Looking at the video from the same timestamp, I could see a small vessel passing directly over her as she surfaced. At the time this vessel passed over her, the tag was only 0.8 m under the surface of the water!
Image 3: A close encounter between a small vessel and “Scarlett”, shown both on the video from the CATS tag (top) and the spectrogram (bottom). The close call is outlined in a yellow box, when a greater intensity of noise occurred as illustrated by the brighter colour intensity compared to the white box (quieter vessel noise). Brighter colours denote a louder volume. The red boxes show surfacing noise- this can essentially be ignored when interpreting the echogram for our purposes.
Sometimes vessels may be more distant, but possibly equally harmful: we have seen vessel noise from larger and presumably more distant vessels dominate the soundscape in some of the tag data. Remembering that to a whale, the sonic world is as important as the visual world is to us, this elevated background noise from ships could have major consequences. So, the first step is to try to quantify the gray whales’ exposure to this vessel noise. I’ve been running some systematic sampling on the tag data to try to quantify background noise levels, and how this changes depending on the time of day: do individual whales experience the same daily spikes in ocean noise that were detected on the static hydrophones, at around 6am and noon due to vessel traffic (Haver et al., 2023)? If not, are they taking evasive action to avoid these spikes? These are just some of the questions that these CATS tags can help us answer, although ideally we need longer acoustic data recordings to capture day and night data, as well as potentially improving the hydrophones on the CATS tags themselves to minimise the impacts of tag interference and random noise.
When explaining to the public what it is to be a PhD student, I often refer to myself as a ‘scientist in training’, or to young children, a ‘baby scientist’. As I look toward my departure from the GEMM lab, I hope to have developed into at least a scientific toddler, having gained the ability to walk through reams of acoustic data with (relative) independence. More than that, I’m excited to take home a refreshed sense of curiosity about what drives marine mammals to behave as they do, an openness to collaboration and new approaches, and a large dose of ‘American emotion’! Let’s hope my British colleagues can handle it!
My heartfelt thanks to all those who welcomed me so warmly at the GEMM lab and Oregon State University, particularly my mentors Leigh Torres and Samara Haver.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly alert when we make a new post! Just add your name into the subscribe box below!
Bibliography
Sonic Sea (2017) Directed by Michelle Dougherty [Film] Distributed by the Natural Resources Defense Council.
Burnham, R., Duffus, D. & Mouy, X. (2018) Gray Whale (Eschrictius robustus) Call Types Recorded During Migration off the West Coast of Vancouver Island. Frontiers in Marine Science. 5, 329. doi:10.3389/fmars.2018.00329.
Colson, K., E. Pirotta L. New, D Cade, J Calambokidis, K. Bierlich, C Bird, A Fernandez Ajó, L. Hildebrand, A. Trites, L. Torres. (in press). Using accelerometry tags to quantify gray whale foraging behavior. Marine Mammal Science.
Clayton, H., Cade, D.E., Burnham, R., Calambokidis, J. & Goldbogen, J. (2023) Acoustic behavior of gray whales tagged with biologging devices on foraging grounds. Frontiers in Marine Science. 10, 1111666. doi:10.3389/fmars.2023.1111666.
Dahlheim, M. & Castellote, M. (2016) Changes in the acoustic behavior of gray whales Eschrichtius robustus in response to noise. Endangered Species Research. 31, 227–242. doi:10.3354/esr00759.
Fish, J.F., Sumich, J.L. & Lingle, G.L. (n.d.) Sounds Produced by the Gray Whale, Eschrichtius robustus.
Guazzo, R., Schulman-Janiger, A., Smith, M., Barlow, J., D’Spain, G., Rimington, D. & Hildebrand, J. (2019) Gray whale migration patterns through the Southern California Bight from multi-year visual and acoustic monitoring. Marine Ecology Progress Series. 625, 181–203. doi:10.3354/meps12989.
Guazzo, R.A., Helble, T.A., D’Spain, G.L., Weller, D.W., Wiggins, S.M. & Hildebrand, J.A. (2017) Migratory behavior of eastern North Pacific gray whales tracked using a hydrophone array S. Li (ed.). PLOS ONE. 12 (10), e0185585. doi:10.1371/journal.pone.0185585.
Haver, S.M., Haxel, J., Dziak, R.P., Roche, L., Matsumoto, H., Hvidsten, C. & Torres, L.G. (2023) The variable influence of anthropogenic noise on summer season coastal underwater soundscapes near a port and marine reserve. Marine Pollution Bulletin. 194, 115406. doi:10.1016/j.marpolbul.2023.115406.
Lemos, L.S., Haxel, J.H., Olsen, A., Burnett, J.D., Smith, A., Chandler, T.E., Nieukirk, S.L., Larson, S.E., Hunt, K.E. & Torres, L.G. (2022) Effects of vessel traffic and ocean noise on gray whale stress hormones. Scientific Reports. 12 (1), 18580. doi:10.1038/s41598-022-14510-5.
Oestreich, W.K., Oliver, R.Y., Chapman, M.S., Go, M.C. & McKenna, M.F. (2024) Listening to animal behavior to understand changing ecosystems. Trends in Ecology & Evolution. S0169534724001459. doi:10.1016/j.tree.2024.06.007.
Pirotta, E., Fernandez Ajó, A., Bierlich, K.C., Bird, C.N., Buck, C.L., Haver, S.M., Haxel, J.H., Hildebrand, L., Hunt, K.E., Lemos, L.S., New, L. & Torres, L.G. (2023) Assessing variation in faecal glucocorticoid concentrations in gray whales exposed to anthropogenic stressors S. Cooke (ed.). Conservation Physiology. 11 (1), coad082. doi:10.1093/conphys/coad082.
Rolland, R.M., Parks, S.E., Hunt, K.E., Castellote, M., Corkeron, P.J., Nowacek, D.P., Wasser, S.K. & Kraus, S.D. (2012) Evidence that ship noise increases stress in right whales. Proceedings of the Royal Society B: Biological Sciences. 279 (1737), 2363–2368. doi:10.1098/rspb.2011.2429.
Sullivan, F.A. & Torres, L.G. (2018) Assessment of vessel disturbance to gray whales to inform sustainable ecotourism. The Journal of Wildlife Management. 82 (5), 896–905. doi:10.1002/jwmg.21462.
Dr. Alejandro A. Fernández Ajó, Postdoctoral Scholar, Marine Mammal Institute – OSU Department of Fisheries, Wildlife, & Conservation Sciences, Geospatial Ecology of Marine Megafauna (GEMM) Lab.
Whales are exposed to an increasing number of human-induced stressors—ranging from pollution and bycatch to the impacts of climate change on prey quality and distribution. Understanding how these factors affect whale health is critical for their conservation. The use of alternative approaches (i.e., alternative to blood samples) for gathering physiological information on large whales using a variety of non-lethal and non to minimally invasive sample matrices (i.e., blubber biopsies, blow, and fecal samples) provides a window into their endocrine state, allowing researchers to assess how these animals respond to both short-term and long-term stressors, and assess their reproductive and nutritional status. However, a lack of standardized protocols might hinder the comparability of results across studies, making it difficult to draw broad conclusions about the health and reproductive parameters of different whale populations.
Dr. Logan Pallin and I organized a lab exchange, funded by The Company of Biologists, to start a new collaboration aimed at bridging this gap by validating and standardizing methods for endocrine assessments in whale blubber. This is not just a technical exercise; it is a foundational step towards building equity and capacity in laboratories worldwide to conduct reliable and comparable endocrine assessments, enhancing the opportunities for multi-lab collaborations. Through this exchange, we aim to consolidate a standardized approach that will yield consistent results between laboratories, enabling better comparisons across different large whale populations. Hosted by the University of California Santa Cruz Biotelemetry and Behavioral Ecology Lab (UCSC-BTBEL Lab) under the mentorship of Dr. Logan Pallin, this experience is instrumental in advancing my research on large whale ecology and conservation.
Dr. Logan Pallin and Alejandro Fernandez Ajó conducting hormone extractions from gray whale blubber samples (left). Preparing a microtiter assay plate for hormone quantification in blubber (right).
During this exchange at the BBE Lab, I had the privilege of working closely with Dr. Logan Pallin, whose expertise in large whale endocrinology (particularly analyzing blubber biopsies) has been instrumental in shaping modern approaches to whale research. The lab’s cutting-edge equipment and Logan’s extensive experience with hormone extraction and quantification methods provided an ideal setting for refining our protocols. Our work focused on the extraction and quantification of progesterone from gray whale blubber samples provided by the Oregon State University Marine Mammal Stranding Network, part of MMI. These large blubber sections allow for repeated sub-sampling to ensure that the selected immunoassays reliably detect and measure the hormones of interest, while also assessing potential sources of variability when applying a standardized protocol. We initially focused our tests and validations on progesterone, as it is the precursor of all major steroid hormones and serves as an indicator of reproductive state in females.
A fieldwork day off Monterrey Bay, California with Dr. Logan Pallin, and PhD candidate Haley Robb. Blubber. Blubber biopsies can be obtained from free swimming whales with minimally invasive methods. From each sample we can derive multiple information about the reproductive status, genetics and overall health of the individuals.
The broader impact of our work The successful validation and standardization of these protocols represents a significant advancement in whale conservation physiology. Once these methods are established, we plan to acquire funds to apply them to a larger collection of blubber samples. We hope to expand our work to include other species and regions, building a broader network of researchers dedicated to studying large whales in a rapidly changing world, and to assess hormone profiles in relation to factors like reproductive success, body condition, and exposure to stressors such as vessel traffic and environmental changes.
During our fieldwork in Monterey Bay, we had fascinating encounters with Minke whales (Balaenoptera acutorostrata, top left), a large group of Risso’s dolphins (Grampus griseus, bottom left), playful Humpbacks (Megaptera novaeangliae, top right), and a Blue whale (Balaenoptera musculus, no photo).
As I conclude this lab exchange, I am filled with excitement for the future. The knowledge and skills gained during this experience will undoubtedly shape the next phase of my research, allowing me to contribute more effectively to the conservation of these incredible animals. I look forward to applying these standardized methods to ongoing and future projects, and to continuing this fruitful collaboration with the BBE Lab. This journey has reinforced the importance of collaboration, standardization, and innovation in the field of conservation physiology. By working together, we can better understand the complex lives of large whales and take meaningful steps towards their protection in an increasingly challenging environment.
Acknowledgments: This exchange was made possible by the support of The Company of Biologists Traveling Fellowship Grant. I would like to thank Dr. Ari Friedlaender (BBE Lab PI) for facilitating this exchange, and Dr. Leigh Torres (GEMM Lab PI) and Dr. Lisa Balance (MMI director) for their support in helping me expand my collaboration network and skillsets. Special thanks to PhD student Haley Robb for her assistance in the laboratory and fieldwork, and a heartfelt thank you to Dr. Logan Pallin for generously sharing his knowledge and time.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly message when we post a new blog. Just add your name and email into the subscribe box below.
By Rachel Kaplan, PhD candidate, Oregon State University College of Earth, Ocean, and Atmospheric Sciences and Department of Fisheries, Wildlife, and Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
I moved to Corvallis exactly four years ago, in the deep, dark midst of the Covid pandemic, and during the added chaos of the 2020 Labor Day Fires, some of the worst in Oregon’s history. I vividly remember attending our virtual lab meeting sitting on the floor surrounded by boxes, while my labmates told me their own stories (many, surprisingly!) of moving during natural disasters. At the time, beginning graduate school represented so many big changes in my life: I had quit my job, sold my furniture, and moved across the country, hoping to explore an area of research that had been calling to me for years, and to gain a new skillset and confidence.
Highlight: A very pandemic cruise. My first day of marine mammal fieldwork in 2021, at sea with (now Dr.) Dawn Barlow.
Now, I’m starting the fifth year of my PhD, thinking about all that has happened and all that is to come. Graduate school is full of milestones to mark time and progress: I’ve taken the courses required for my program, sat for a written exam to test my broad knowledge of oceanography, and written a dissertation proposal. Earlier this year, I spent two months buried in the literature on oceanography, krill, and whale ecology in preparation for my oral qualifying exam. I’ve stared at the water for dozens of hours watching for whales off the Oregon coast, and experienced polar night studying winter krill in Antarctica. I’ve conquered my fear of learning to code, and felt constant, profound gratitude for the amazing people I get to work with.
The last four years have been incredibly busy and active, but now more than ever, it feels like the time to really do. I can see the analytical steps ahead for my final two dissertation chapters more clearly than I’ve been able to see either of the other two chapters that have come before. One of my favorite parts of the process of research is discussing analytical decisions with my labmates and supervisors, and experiencing how their brains work. Much of our work hinges on modeling relationships between animals and their environment. A model, most fundamentally, is a reduced-scale representation of a system. As I’ve learned to use statistical models to understand relationships between krill and whales, I have simultaneously been building a mental model of the Northern California Current (NCC) ecosystem and the ecological relationships within it. Just as I have long admired in my supervisors and labmates, I can now feel my own mind becoming more playful as I think about this ocean environment, the whales and krill that make a living in the NCC, and the best way to approach studying them analytically.
Highlight: Working on my dissertation proposal during a friend’s 2022 wedding celebration in Utah.
Graduate school demands that you learn and work to constantly exceed your own bounds, and pushing to that extent for years is often stressful and even existentially threatening. However, this process is also beautiful. I have spent the last four years growing in the ways that I’ve long wanted to, and reveled in feeling my mind learn to play. I wouldn’t give up a moment of the time I’ve spent in the field, the relationships I’ve built with my labmates, or the confidence I’ve developed along the way.
As I look ahead to this next, final, year of graduate school, I hope to use what I’ve learned every day – and not just about how to conduct research, but about myself. I want to always remember that krill, whales, and the ocean ecosystem are incredible, and that it is a privilege to study them. I hope to work calmly and intentionally, and to continue appreciating this process of research and growth.
Highlight: My first in-person oral presentation, at the 2024 ICES-PICES International Zooplankton Production Symposium in Hobart, Tasmania.