By Dr. Lisa Hildebrand
I successfully defended my PhD 55 days ago and I have been grappling with a lot of complex emotions since then. I feel pride in having accomplished such a milestone, sadness to have left such an incredible lab and workplace, excitement to see what the future holds, and, admittedly, also relief to have a bit of time off. I have also been reflecting about all the things I have learned during my seven-year journey of being a graduate student. Naturally there is all the ecological, quantitative and communication knowledge and skills I learned that have shaped me into the scientist I am today. However, there are so many lessons that I have learned that are less tangible but equally as important in building a successful career in academia, I think.
Feeling stupid is kind of the point
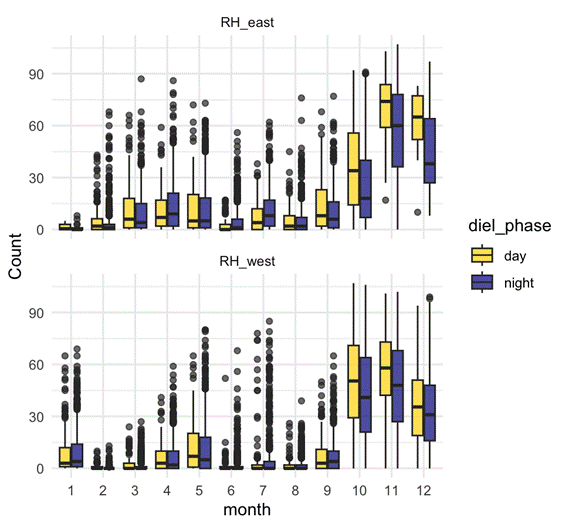
Graduate school is riddled with unknowns. As we embark on these journeys there is much to be learned and it can be exhausting not knowing a lot of the time. During many stints of my graduate school journey, I felt stupid, with varying magnitudes of stupidity. Whether I felt stupid because I had to Google “how to save one object in R environment” for the thousandth time or because I simply could not grasp why a Bayesian posterior distribution was so much better than a frequentist p-value result. If you’re struggling with feeling stupid in academia, know that it is actually an important part of the process – sitting in the discomfort of not knowing something is what drives us to get answers. Of course, in graduate school, it isn’t that we just don’t know the answer to a scientific question (for example, in my case, how do different individual gray whales use space during a foraging season?), but we also don’t know how to get the answers to that question right away. Before I could even begin to tackle that question for one of my PhD chapters, I had to learn about how to quantify survey effort from survey conditions, the principles of Bayesian statistics, theoretical spatial ecology, coding in Python, capture-recapture models, and so much more. Each of those pursuits involved weeks, if not months, of feeling stupid. But feeling stupid throughout the pursuits was kind of the point. There’s an incredibly excellent (and short) article on this topic by Martin Schwartz that I cannot recommend highly enough.
Asking questions isn’t a weakness, it’s a strength
Somewhat related to the importance of feeling stupid, is realizing that it’s ok, neigh even necessary, to ask for help. Learning to be an independent problem-solver is of course a huge part of being a graduate student. But it’s also important to learn that at some points, continuing to bang your head against the keyboard because you can’t figure something out, is not the most productive use of your time. I have been so lucky to have been surrounded by so many knowledgeable, generous and kind people throughout my graduate career, ranging from fellow graduate students to lab mates, to project colleagues, and my committee members and advisor. I have benefitted so much from countless Zoom meetings or whiteboarding sessions where people have shared their wisdom with me in helping me solve a problem. I know this quote is probably a cliché but I think it holds so much truth: “Ask a question and be a fool for five minutes, don’t ask a question and be a fool for life.”. More often than not, a question that you ask is one that someone else probably also had in mind but was too shy or embarrassed to ask. So, be the risk taker and ask that question!

Learning takes time
The months leading up to a defense are usually a massive crunch. There are analyses that are still getting figured out, chapters that need to be written, citations that need to be integrated, and a big beefy Word document that needs to be formatted according to very specific thesis guidelines. By the end of a degree, a graduate student has learned so much and honed so many skills that tasks are happening much more efficiently than they did the first time around. A few weeks before my defense, I had a conversation with Marine Mammal Institute research associate Selene Fregosi where I lamented about not having achieved enough in the first three years of my PhD seeing as I was able to complete two data analyses and chapters in the final six months of my PhD. It left me wondering, what did I spend all those years doing at the start of my PhD?! Selene chuckled in response and she said, “Lisa, you were learning!”. Selene helped me realize that it was those years spent in the slow learning process that allowed me to be fast yet effective in the final months before my defense, enabling the completion of two chapters in a much shorter amount of time than my first chapters took. That flip in perspective was a much kinder way of looking at the years gone by. Dovetailed with this lesson is that everything takes way longer than you think it will, especially if it involves learning, so budget for learning time!
Adaptability is key
I consider myself a Type A personality. I like being organized, I like having a plan, and I like knowing what is around every corner. Unfortunately, a graduate student trajectory (and life!) rarely has a defined, clear, unchanging path. I recently looked back at a document titled ‘PhD Chapter Ideas’ that I made in June 2020 while I was in the final six months of my Master’s degree and starting to brainstorm with Leigh about what my PhD dissertation could entail. Of the six ideas listed in that document, only two of them sort of align with the chapters that ended up in my final dissertation five years later. My dissertation morphed quite a bit over the years and this change is inevitable. So many things can go wrong during scientific endeavors: equipment can malfunction and fail, data may not be available in time or impossible to acquire, proposed collaborations may not take place, weather, funding hiccups, computational limitations, to name a few. While it is important to be persistent and to find solutions to problems that arise, sometimes the solution to a problem may be pivoting and embracing change.

Thinking ahead
As I begin the next, somewhat daunting, journey of entering the job market, I am reminded that graduate school has shaped me in ways I did not always recognize at the time. I take comfort in knowing that graduate school has prepared me in ways that extend far beyond technical training. Alongside tools and knowledge, I leave with resilience, perspective, and a collection of hard-earned lessons that will continue to guide me as I navigate what comes next.