By Lisa Hildebrand, PhD student, OSU Department of Fisheries, Wildlife, & Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
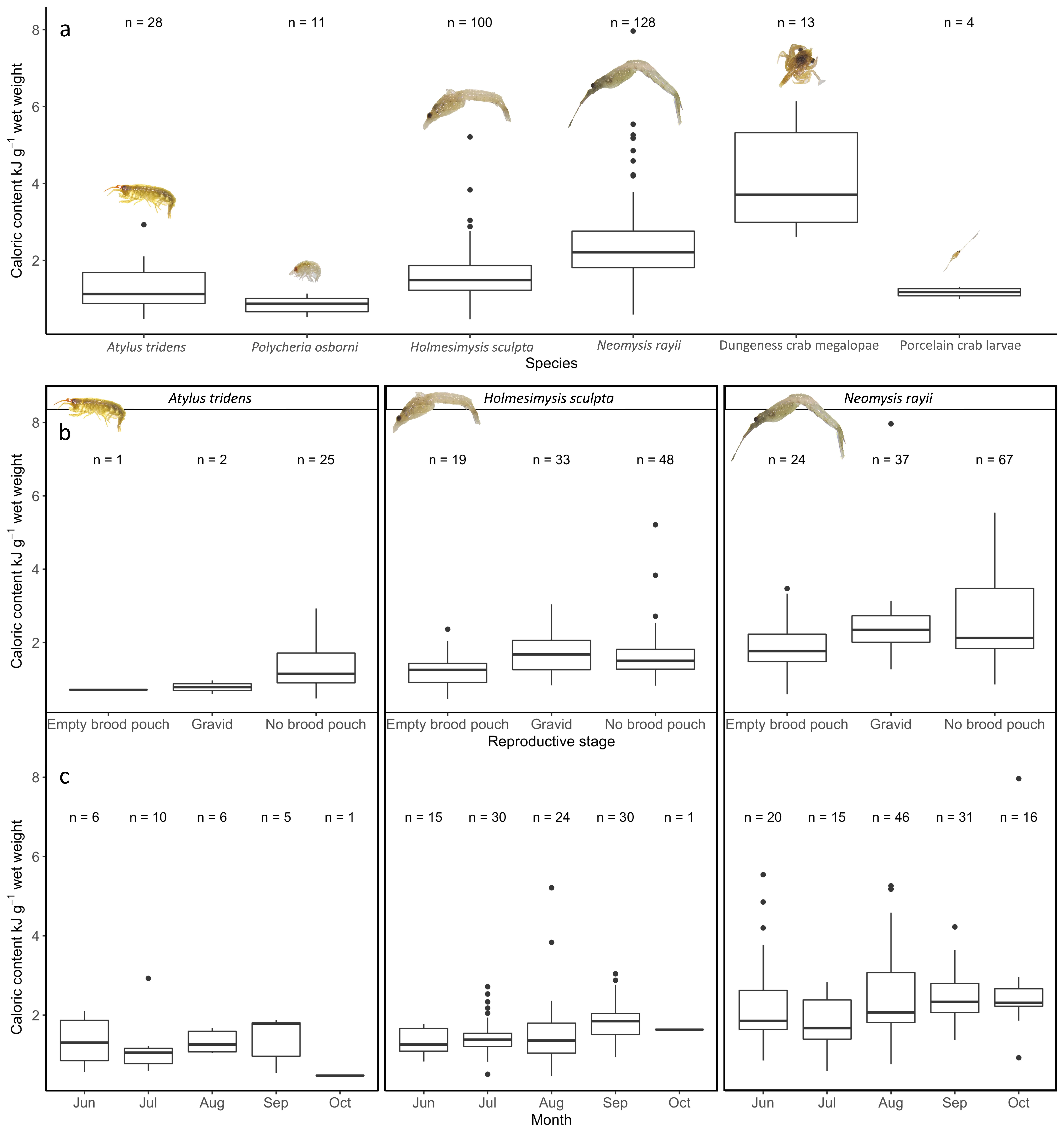
Since we find ourselves well into the cozy winter season, I thought it was an appropriate time to update you all on our project COZI (Coastal Oregon Zooplankton Investigation). COZI is a cross-college collaborative effort, led by GEMM PI Leigh Torres, that aims to better understand the quality of Oregon coast zooplankton prey and its impacts on gray whale foraging ecology and health. Leigh is joined by three other early-career female scientists, Dr. Sarah Henkel, Dr. Kim Bernard, and Dr. Susanne Brander, that each contribute a different area of expertise to the project. The quartet recently graced the cover of the Oregon Stater in an article all about COZI written by Nancy Steinberg (which I highly recommend reading!). To date, the COZI team (which includes myself as well as many other students) has found that the caloric content of the six predominant zooplankton species in Oregon coastal waters differs significantly, with Dungeness crab megalopae coming out on top as a caloric goldmine (Hildebrand et al. 2021). We found that these Oregon prey are calorically competitive with the predominant benthic amphipod that gray whales feed on in the Arctic, which has interesting implications for foraging ground selection and use of gray whales in the eastern North Pacific (read about it in detail in my blog about the publication). Now that we know that Oregon zooplankton quality differs in terms of calories, we are curious to determine whether these species are impacted by microplastics in the environment, to what extent, and how gray whales may be affected.

To answer these questions, we are analyzing both zooplankton and gray whale fecal samples for microplastics to see what kind, and how many, microplastics we find, and whether microplastics biomagnify up the food chain. The lab analysis has just been completed and we are working on interpreting the results. We can’t let the cat out of the bag yet, but a little sneak-peek of what we have found is that there are different levels of microplastic loads by zooplankton species, which also end up in the whale poop. So, until we finalize those results for sharing, I am going to review the field of microplastics research, with a particular focus on cetaceans. Avid readers of our blog may recall that I wrote a blog about marine plastics at the start of 2019. In that blog, I mentioned that a GoogleScholar search of “microplastics marine” generated 7,650 results. To get an idea of how microplastics research in the marine environment has progressed since I wrote my 2019 blog, I conducted the same GoogleScholar search for this blog but I limited the results to studies published between 2019-2021. GoogleScholar presented me with a whopping 18,000+ results, which shows the rapidity at which the field of marine microplastics research has grown in the last couple of years. The studies span all kinds of topics from distribution & occurrence, to chemical behaviors & interactions with other toxins, to sources & sinks (to name a few!). The results encompass both laboratory and field studies investigating samples from all five oceans of the world. Unfortunately, the title of my blog from two years ago still rings very true: plastics truly are ubiquitous in the marine environment.
In my last blog, I listed three cetacean species that had been found to contain microplastics: a True’s beaked whale (Lusher et al. 2015), a humpback whale (Besseling et al.2015) and an Indo-Pacific humpback dolphin (Zhu et al.2018). Reflective of the marine microplastics field in general, this list has also grown considerably in the last two years. Since 2019, microplastics have been detected in harbor porpoises (Philipp et al. 2021), common dolphins (Nelms et al. 2019), striped dolphins (Novillo et al. 2020), bottlenose dolphins (Battaglia et al. 2020), Atlantic white-sided dolphins (Nelms et al. 2019), beluga whales (Moore et al. 2020), and Bryde’s & sei whales (Zantis et al. 2021). At this point, I would posit that the main reason this list is not longer is due to the time it takes to collect and analyze samples for microplastics, rather than microplastics being absent in other cetacean species. During my research for this blog, I noticed that the studies on microplastics in cetaceans are starting to shift from focusing on simply determining microplastic occurrence to attempting to estimate levels of exposure and/or ingestion, determine the main source (from water vs. from prey), and long-term consequences.

A study published this year examined fecal samples of Bryde’s and sei whales in coastal waters in New Zealand and detected 32 ± 24 microplastics per 6 g of feces (Zantis et al. 2021). By extrapolating these values to the proportions of prey species in the whales’ diet, the authors estimate that these whales consume over 24,000 pieces of microplastics per mouthful of prey, or more than 3 million microplastics per day. Another study (Shetty 2021) in the same geographic region investigated the levels of microplastics in coastal surface waters, which allowed the authors to estimate whether the source of the microplastics that the Bryde’s and sei whales ingest come from the water or the prey. They found that the estimated level of microplastics that the whales consume daily from their prey is four orders of magnitude higher than the microplastic levels in the coastal waters. This finding strongly suggests that the predominant mode of exposure of large filter feeders, such as baleen whales, for microplastic pollution comes from their prey through biomagnification (not just from the ambient sea water).

COZI aims to conduct similar analyses as these studies described above to understand the exposure of coastal Oregon zooplankton to microplastics and how this may be affecting gray whales. Stay tuned for those results!
I am aware that I have painted a very bleak (but true) picture of microplastic pollution in our oceans in this blog but there are things you can do to help reduce microdebris in the environment!
- A major source of pollution in the ocean comes from microfibers through our laundry. You can help stop this pathway by simply using a Cora Ball or installing a filter (such as this one) in your washing machine that captures microfleece & polyester fibers.
- Minimize your use of single-use plastics. There are so many ways to do so including reuseable water bottles, travel mugs for coffee or tea, fabric totes as shopping bags, carry a set of utensils for takeout food, beeswax wraps instead of plastic wrap or sandwich bags.
- Use public transport when possible as another huge source of microplastics comes from tire treads! This solution also helps reduce your carbon footprint.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly message when we post a new blog. Just add your name into the subscribe box below.
References
Battaglia, F.M., Beckingham, B.A., & McFee, W.E. 2020. First report from North America of microplastics in the gastrointestinal tract of stranded bottlenose dolphins (Tursiops truncatus). Marine Pollution Bulletin 160:111677.
Besseling, E., et al. 2015. Microplastic in a macro filter feeder: humpback whale Megaptera novaeangliae. Marine Pollution Bulletin 95: 248-252.
Hildebrand, L., Bernard, K.S., & Torres, L.G. 2021. Do gray whales count calories? Comparing energetic values of gray whale prey across two different feeding grounds in the eastern North Pacific. Frontiers in Marine Science. https://doi.org/10.3389/fmars.2021.683634
Lusher, A.L., et al. 2015. Microplastic and macroplastic ingestion by a deep diving, oceanic cetacean: the True’s beaked whales Mesoplodon mirus. Environmental Pollution 199: 185-191.
Moore, R.C., et al. 2020. Microplastics in beluga whales (Delphinapterus leucas) from the eastern Beaufort Sea. Marine Pollution Bulletin 150:110723.
Nelms, S.E., et al. 2019. Microplastics in marine mammals stranded around the British coast: ubiquitous bus transitory? Scientific Reports 9:1075.
Novillo, O., Raga, J. A., & Tomás, J. 2020. Evaluating the presence of microplastics in striped dolphins (Stenella coeruleoalba) stranded in the western Mediterranean Sea. Marine Pollution Bulletin 160:111557.
Philipp, C., et al. 2021. First evidence of retrospective findings of microplastics in harbor porpoises (Phocoena phocoena) from German waters. Frontiers in Marine Science. https://doi.org/10.3389/fmars.2021.682532
Shetty, D. 2021. Incidence of microplastics in coastal inshore fish species and surface waters in the Hauraki Gulf, New Zealand. Master’s thesis, University of Auckland, New Zealand.
Zantis, L.J., et al. 2021. Assessing microplastic exposure of large marine filter-feeders. Science of The Total Environment 151815.
Zhu, J., et al. 2018. Cetaceans and microplastics: First report of microplastic ingestion by a coastal delphinid, Sousa chinensis. Science of the Total Environment 659: 649-654.










































{kind=link}