As I sat down to write this blog, I realized that it is the first post I have written in 2023! This is largely because I have spent the last seven weeks preparing for (and partly taking) my PhD qualifying exams, an academic milestone that involves written and oral exams prepared by each committee member for the student. The point of the qualifying exams is for the student’s committee to determine the student’s understanding of their major field, particularly where and what the limits of that understanding are, and to assess the student’s capability for research. How do you prepare for these exams? Reading. Lots of reading and synthesis of the collective materials assigned by each committee member. My dissertation research covers a broad range of Pacific Coast Feeding Group (PCFG) gray whale ecology, such as space use, oceanography, foraging theory and behavioral responses to anthropogenic activities. Accordingly, my assigned reading lists were equally broad and diverse. For today’s blog, I am going to share some of the papers that have stuck with me and muse about how these topics relate to my study system, the Pacific Coast Feeding Group (PCFG) of gray whales.
Space use & home range
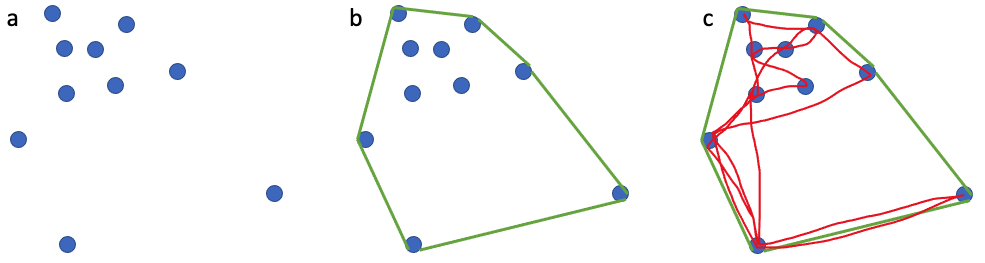
For decades, ecologists have been interested in defining an animal’s use of space through time, often referred to as an animal’s home range. The seminal definition of a home range comes from Burt (1943) who outlined it as “the area traversed by an individual in its normal activities of food gathering, mating, and caring for young.”. I like this definition of a home range because it is biologically grounded and based on an animal’s requirements. However, quantifying an animal’s home range based on this definition is harder than it may sound. In an ideal world, it could be achieved if we were able to collect location data that is continuous (i.e., one location per second), long-term (i.e., at least half the lifespan of an animal) and precise (i.e., correct to the nearest meter) together with behavior for an individual. However, a device that could collect such data, particularly for a baleen whale, does not currently exist. Instead, we must use discontinuous (i.e., one location per hour, day or month) and/or short-term (i.e., <1 year) data with variable precision to calculate animal home ranges. A very common and simple analytical method that is used to calculate an animal’s home range is the minimum convex polygon (MCP). MCP draws the smallest polygon around points with all interior angles less than 180º. While this method is appealing and widely used, it often overestimates the home range by including areas not used by an animal at all (Figure 1).
Figure 1. (a) 10 point locations where an individual was observed; (b) the home range as determined by the minimum convex polygon method; (c) the red path shows the movements the animal actually took. Note the large white area in (c) where the animal never went even though it is considered part of the animal’s home range.
This example is just one of many where home range estimators inaccurately describe an animal’s space use. However, this does not mean that we should not attempt to make our best approximations of an animal’s home range using the tools and data we have at our disposal. Powell & Mitchell perfectly summarized this sentiment in their 2012 paper: “Understanding animal’s home ranges will be a messy, irregular, complex process and the results will be difficult to map. We must embrace this messiness as it simply represents the real behaviors of animals in complex and variable environments.”. For my second dissertation chapter, I am investigating individual PCFG gray whale space use patterns by calculating activity centers and ranges. The activity center is simply the geographic center of all points of observation (Hayne, 1949) and the range is the distance from the activity center to the most distant point of observations in either poleward direction. While the actual activity center is probably relatively meaningless to a whale, we hope that by calculating these metrics we can identify different strategies of space use that individuals employ to meet their energetic requirements (Figure 2).
Figure 2. Sightings of nine different PCFG individuals across our GRANITE study area. Each circle represents a location where an individual was sighted and circles are color-coded by year. Plotting the raw data of sighting histories of these individuals hints at patterns in space use by different individuals, which I will explore further in my second dissertation chapter.
Non-stationary responses to oceanography
Collecting spatiotemporally overlapping predator-prey datasets at the appropriate scales is notoriously challenging in the marine environment. As a result, marine ecologists often try to find patterns between marine species and oceanographic and/or environmental covariates, as these can sometimes be easier to sample and thus make marine species predictions simpler. This approach has been applied successfully in hundreds, if not thousands, of studies (e.g., Barlow et al., 2020; Derville et al., 2022). Unfortunately, these relationships are not always proving to be stable over time, a phenomenon called non-stationarity. For example, Schmidt et al. (2014) showed that the reproductive successes of Brandt’s cormorants and Cassin’s auklets on southeast Farallon Island were positively correlated with each other from 1975 to 1995 and were associated with negative El Niño-Southern Oscillation. However, around the mid-1990s this relationship broke down and by 2002, the reproductive successes of the two species were significantly negatively correlated (Figure 3). Furthermore, the relationships between reproductive success and most physical oceanographic conditions became highly variable from year to year and were non-stationary. Thus, if the authors continued to use the relationships defined early on in the study (1975-1995) to predict seabird reproductive success relative to ocean conditions from 2002-2012, their predictions would have been completely wrong. After reading this study, I thought a lot about what the oceanographic conditions have been since the GEMM Lab started studying PCFG gray whales vs. the years prior. Leigh launched the GRANITE project in 2016, right at the tail end of the record marine heatwave in the Pacific, known as “the Blob”. While we do not have as long of a dataset as the Schmidt et al. (2014) study, I wonder whether we might find non-stationary responses between PCFG gray whales and environmental and/or oceanographic variables, given how the effects of the Blob lingered for a long time and we may have captured the central Oregon coast environment shifting from ‘weird to normal’. Non-stationarity is something I will at least keep in mind when I am working on my third dissertation chapter which will investigate the environmental and oceanographic drivers of PCFG gray whale space use strategies.
Figure 3. Figure and caption taken from Schmidt et al. (2014).
There are so many more studies and musings that I could write about. I keep being told by others who have been through this qualifying exam process that this is the smartest I am ever going to be, and I finally understand what they mean. After spending almost two months in my own little study world, my research, and where it fits within the complex web of ecological knowledge, has snapped into hyperfocus. I can see clearly where past research will guide me and where I am blazing a new trail of things never attempted before. While I still have the oral portion of my exams before me (in fact, it’s tomorrow!), I am already giddy with excitement to switch back to analyzing data and making progress on my dissertation research.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly message when we post a new blog. Just add your name and email into the subscribe box below.
References
Barlow, D.R., Bernard, K.S., Escobar-Flores, P., Palacios, D.M., Torres, L.G. 2020. Links in the trophic chain: modeling functional relationships between in situ oceanography, krill, and blue whale distribution under different oceanographic regimes. Marine Ecology Progress Series 642: 207−225.
Burt, W.H. 1943. Territoriality and home range concepts as applied to mammals. Journal of Mammalogy 24(3): 346-352. https://doi.org/10.2307/1374834
Derville, S., Barlow, D.R., Hayslip, C., Torres, L.G. 2022. Seasonal, annual, and decadal distribution of three rorqual whale species relative to dynamic ocean conditions off Oregon, USA. Frontiers in Marine Science 9. https://doi.org/10.3389/fmars.2022.868566
Hayne, D.W. 1949. Calculation of size of home range. Journal of Mammalogy 30(1): 1-18.
Powell, R.A., Mitchell, M.S. 2012. What is a home range? Journal of Mammalogy 93(4): 948-958. https://doi.org/10.1644/11-MAMM-S-177.1
Schmidt, A.E., Botsford, L.W., Eadie, J.M., Bradley, R.W., Di Lorenzo E., Jahncke, J. 2014. Non-stationary seabird responses reveal shifting ENSO dynamics in the northeast Pacific. Marine Ecology Progress Series 499: 249-258. https://doi.org/10.3354/meps10629
Another year has come and gone, and with the final days of 2022 upon us, it is my honor and pleasure to present to you, dear reader, this summary of achievements by the GEMM Lab this year. It has been another big year for us, so snuggle up with your favorite holiday drink and enjoy our recap of 2022!
Leigh working hard during GRANITE field work
2022 was a huge year of milestones for each lab member. The biggest happened just a few weeks ago when, on December 1st, our primary investigator (PI) and the captain at the GEMM Lab helm, Leigh Torres, started her sabbatical!!! Leigh, who received tenure and became an Associate Professor in 2020, was eligible for a sabbatical this year and took the opportunity to take a very well-deserved three months in New Zealand with her family. Leigh established the GEMM Lab in 2014, and it has since grown into a 13-person strong team that aims to advance marine science and conservation through innovative and engaged research across 11 active projects. I know I speak for all my lab mates when I say that we are incredibly grateful and thankful for Leigh, who always prioritizes us, even when she is busy with other things. Leigh, enjoy New Zealand and your time off! Your crew will man the GEMM Lab ship while you are away, under the leadership of the four postdocs, your first mates. Speaking of which…
Dawn Barlow defended her PhD dissertation “Ecology and Distribution of Blue Whales in New Zealand Across Spatial and Temporal Scales” in April and became the latest Dr. of the GEMM Lab! Dawn’s achievements were recognized by OSU’s College of Agricultural Sciences as she was awarded the prestigious Savery Outstanding Doctoral Student Award in the spring. Finishing her PhD also marked the culmination of a decade of blue whale research in New Zealand, which began with Leigh’s hypothesis of a resident blue whale population in the region. Thankfully, we have not had to say goodbye to Dawn as she is now a GEMM Lab postdoctoral scholar (more below). The milestones kept coming after Dawn’s defense as PhD student Clara Bird became PhD candidate Clara Bird in April after passing her qualifying exams. Four of us – MS students Allison Dawn and Imogen (also called Miranda) Lucciano, and PhD students Rachel Kaplan and myself – successfully defended our research proposals to our committees and had fruitful discussions about how to best accomplish our ambitious proposed research. Morgan O’Rourke-Liggett rejoined the GEMM Lab after being the undergraduate intern in the 2017 TOPAZ/JASPER (Theodolites Overlooking Predators and Prey / Journey for Aspiring Students Pursuing Ecological Research) field season, and they completed their graduate certificate in Geographic Information Systems in the Fall. For their capstone project, Morgan is now working on accounting for GRANITE (Gray whale Response to Ambient Noise Informed by Technology and Ecology) survey effort in order for us to then understand whether and how distribution patterns of gray whales have changed. Finally, Imogen completed her Graduate Certificate of Wildlife Management and moved into an M.Sc. program. Hip-hip-hurrah for all of these degree milestones!
Clockwise: New Dr. Dawn Barlow with her committee after successfully defending her PhD dissertation; Lisa (me) celebrating after a successful PhD research review; Dawn and Leigh during a successful OPAL field day; the R/V Bell M. Shimada science team on the September cruise (Rachel is second from left in the back row); the 2022 TOPAZ/JASPER field team hard at work
This year, it felt like someone in the GEMM Lab was always either preparing for fieldwork, in the field, or completing the post-fieldwork tasks of gear maintenance and data download. This reality is not surprising given that we have five active projects that involve fieldwork, which keep us busy on the ocean. Another two successful gray whale field seasons are on the books! Our project GRANITE wrapped its 7th consecutive year of field work in Newport on October 15th, while the integrated projects TOPAZ/JASPER completed an 8th consecutive field season in Port Orford at the end of August. The GRANITE field team grew with the addition of Master’s student Kate Colson, who is co-advised by Leigh and Dr. Andrew Trites at University of British Columbia. Down south in Port Orford, Allison successfully led her first solo field season after taking over the project from me last year. But the nearshore is not the only place that captured the GEMM Lab’s attention. HALO (Holistic Assessment of Living marine resources off Oregon) completed three survey cruises in January, June, and July, which included the successful recovery and replacement of three hydrophones, providing Imogen and Cornell PhD student Marissa Garcia with their long-awaited acoustic data. Imogen oversees cruise coordination for this GEMM Lab effort, and several lab members have gone to sea for HALO, including Imogen, Rachel, Dawn and Leigh. We also continued our participation in the Northern California Current (NCC) cruises, where we collect marine mammal and krill data for the OPAL (Overlap Predictions About Large whales) project. Dawn, Rachel and Clara all headed out together on NOAA’s R/V Bell M. Shimada in May, while Rachel was the sole GEMM Lab representative on the September cruise. Offshore biopsy efforts and U.S. Coast Guard helicopter flights also contributed data to OPAL through the year. Finally, Leigh and Dawn also participated in the MMI-wide MOSAIC (Marine Offshore Species Assessment to Inform Clean energy) cruises in August and October. Despite spending so many hours on the water, we were productive onshore too…
Gray whale suction cup taggingHALO hydrophones back!GRANITE field teamDawn during OPAL field workRachel, Dawn & Clara on the NCC cruise
Our faraway postdoc Solène, who has been working remotely from New Caledonia, has made steady progress on the OPAL project. Her biggest achievement this year was finishing the first, NOAA section 6-funded component, and helping to acquire funding for the second phase of the project, which Rachel started work on for her PhD. We were lucky to have Solène visit the lab in January, where she met the new and reunited with the old faces of the GEMM Lab. While her time in Oregon was only 6 weeks or so, we managed to rope her into her first and second gray whale paper (stay tuned for that sometime in 2023). And to top off our quest of making Solène an Oregonian, we are so thrilled to announce that she and her husband Micah have finally acquired their visas to move here in just a few weeks, landing in January 2023!!
Solène & Micah after receiving their visa to come to the USA in January 2023
We have been, and continue to be, busy processing and analyzing all of the rich datasets that we collect during our intense field efforts. While I do not have time to mention all of the work that occurs in the lab and on our computers, I want to highlight some of them. Our postdoctoral scholar Alejandro A. Fernández Ajó is currently back at his graduate institute, Northern Arizona University, conducting lab work to analyze the 63 fecal samples collected from 26 individual gray whales during our 2022 GRANITE field season. Rachel and her amazing team of krill interns have been doing lots of bomb calorimetry all year to better understand the caloric value of different krill species and cohorts. Imogen spent a month at Cornell University in Ithaca, New York, to hone her skills for baleen whale recognition in acoustics data and to become well acquainted with OSU affiliates Dr. Holger Klinck, PhD student Marissa Garcia, and other researchers at the K. Lisa Yang Center of Conservation Bioacoustics.
Even with all these projects underway, it seems that we cannot go a full year in the GEMM Lab without launching new endeavors. 2022 saw the creation of two more projects. For her postdoctoral research, Dawn is leading the newly-launched EMERALD (Examining Marine mammal Ecology through Region-wide Assessment of Long-term Data), which investigates spatiotemporal distribution patterns in harbor porpoise and gray whales in the nearshore NCC waters. Secondly, postdoctoral scholar KC Bierlich and Leigh have received funding to kickstart MMI’s Center of Drone Excellence (CODEX), which will launch in 2023. CODEX will focus on developing open-source tools and software to help analyze drone imagery, with the aim of offering online tutorials and hosting workshops. Both EMERALD and CODEX are funded by sales and renewals of the special Oregon gray whale license plate, which benefits MMI. We gratefully thank all the gray whale license plate holders, who made this research possible, and encourage any Oregonians that don’t have a whale on their tail yet, to do so in 2023!
Describing a year in the life of the GEMM Lab would not be complete without mentioning our outreach and education efforts as well. Allison, Clara, and I put on our teaching hats and gave guest lectures and labs for Dr. Renee Albertson and Dr. Kate Stafford’s marine mammal classes here at OSU as well as host an Introduction to R/RStudio workshop for undergraduates in our roles as coordinators for the Fisheries & Wildlife Mentorship Program. Alejandro gave a virtual talk to graduate students at the University of Pretoria South Africa about conservation physiology, highlighting his research with southern right whales. KC was invited to talk about using drones and computer science to study whales at Newport High School’s Computer Science Course and Oregon Sea Grant’s Whale Ecology Homeschool Program. He also gave the keynote presentation at the 25th Annual Salmon Bowl, part of the National Oceanic Sciences Bowl, which was hosted by OSU in February. Clara and myself were both invited speakers for Cape Perpetua’s monthly speaker series, where we presented our PhD research. Furthermore, GEMM Lab members also presented our work at numerous scientific conferences including the Society of Marine Mammal conference, Ocean Sciences, PICES annual meeting, and TWS Oregon Chapter, to name a few. The dissemination of our work to the scientific community and the public is a central focus of our lab, and we also prioritize providing hands-on opportunities and experiences to students eager to participate in ecological research. We mentored a total of 12 students in 2022, from high school to graduate level, who were involved in all aspects of our research including kayaking in Port Orford to collect prey samples, meticulously measuring drone images of whales, and spending hours hunched over microscopes identifying tiny crustaceans.
Clockwise: 2022 TOPAZ/JASPER team (Charlie, Luke, Allison, Nicola, Zoe); REU student Braden Virgil discussing his poster; krill interns Abby and Henley; REU student Celest with mentors Clara and Leigh
We have once again been prolific writers, contributing 19 total peer-reviewed publications to 15 different scientific journals. If you are in the mood for some holiday reading, you will find the full list of publications at the end of this post. All authors in bold are (or were) GEMM Lab members when the work occurred.
And YOU, our awesome, supportive readers, have once again been busy, with a whopping 25,368 views of our blog this year!!! Thank you for joining us on our 2022 journey! We hope you have enjoyed the tales that we have told and the knowledge we have (hopefully) conveyed. On one final note, if you are still looking for that perfect holiday gift for the whale-lover in your life, and if you want to support our research, consider adopting a whale from our IndividuWhale website. As a small incentive, if you adopt a whale before the end of the year, you will be entered into our Oregon South Coast Whale Watch Experience giveaway! We will reveal the giveaway winner in January 2023. We wish you all restful, happy, and most importantly, healthy holidays, and hope you will join us again in 2023!
The GEMM Lab with their white elephant gifts during our annual holiday party
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly message when we post a new blog. Just add your name and email into the subscribe box below.
Publications
Barlow, D.R., Klinck, H., Ponirakis, D., Holt Colberg, M., Torres, L.G. (In Press). Temporal occurrence of three blue whale populations in New Zealand waters from passive acoustic monitoring. Journal of Mammalogy.
Barlow, D.R., Estrada Jorge, M., Klinck, H., Torres, L.G. (2022). Shaken, not stirred: blue whales show no acoustic response to earthquake events. Royal Society Open Science. 9:220242.
Bierlich, K.C., Hewitt, J., Schick R.S., Pallin, L., Dale, J., Friedlaender, A.S., Christiansen, F., Sprogis K.R., Dawn, A.H., Bird, C.N., Larsen, G., Nichols, R., Shero, M., Goldbogen, J.A., Read, A., Johnston, D.W. (2022). Seasonal gain in body condition of foraging humpback whales along the Western Antarctic Peninsula. Frontiers in Marine Science. 9, 1–16. https://www.frontiersin.org/articles/10.3389/fmars.2022.1036860/full
Cade, D.E., Kahane-Rapport, S.R., Gough, W.T., Bierlich, K.C., Linksy, J.M.J., Johnston, D.W., Goldbogen, J.A., Friedlaender, A.S. (in press). Ultra-high feeding rates of Antarctic minke whales imply a lower limit for body size in engulfment filtration feeders. Nature Ecology and Evolution.
D’Agostino, V.C., Fernández Ajó, A., Degrati, M. et al. Potential endocrine correlation with exposure to domoic acid in Southern Right Whale (Eubalaena australis) at the Península Valdés breeding ground. Oecologia 198, 21–34 (2022). https://doi.org/10.1007/s00442-021-05078-4
Derville, S., Barlow, D.R., Hayslip, C., Torres, L.G. (2022). Seasonal, annual, and decadal shifts of three baleen whale species relative to dynamic ocean conditions off Oregon, USA. Frontiers in Marine Science 9:868566.
Goetz, K.T., Stephenson, F., Hoskins, A., Bindoff, A.D., Orben, R.A., Sagar, P.M., Torres, L.G., et al. (2022). Data quality influences the predicted distribution and habitat of four southern-hemisphere Albatross species. Frontiers in Marine Science 9:782923. https://doi:10.3389/fmars.2022.782923
Gough, W.T., Cade, D.E., Czapanskiy, M.F., Potvin, J., Fish, F.E, Kahane-Rapport, S.R., Savoca, M.S., Bierlich, K.C., Johnston, D.W., Friedlaender, A.S., Szabo, A., Bejder, L., Goldbogen, J.A., (2022). Fast and Furious: Energetic tradeoffs and scaling of high-speed foraging in rorqual whales. Integrative Organismal Biology, 4(1) obac038, https://doi.org/10.1093/iob/obac038
Green, C-P., Ratcliffe, N., Mattern, T., …, Torres, L.G., Hindell, M.A. (2022). The role of allochrony in influencing interspecific differences in foraging distribution during the non-breeding season between two congeneric crested penguin species. PLoS ONE https://doi.org/10.1371/journal.pone.0262901
Hildebrand, L., Sullivan, F.A., Orben, R.A., Derville, S., Torres, L.G. (2022). Trade-offs in prey quantity and quality in gray whale foraging. Marine Ecology Progress Series 695:189-201. https://doi.org/10.3354/meps14115
Hunt, K.E., Buck, C.L., Ferguson, S.H., Fernández Ajó, A., Heide-Jørgensen, M.P., Matthews, C.J.D. (2002). Male Bowhead Whale Reproductive Histories Inferred from Baleen Testosterone and Stable Isotopes, Integrative Organismal Biology, 4: obac014, https://doi.org/10.1093/iob/obac014
Lemos, L.S., Haxel, J.H., Olsen, A., Burnett, J.D., Smith, A., Chandler, T.E., Nieukirk, S.L., Larson, S.E., Hunt, K.E., Torres, L.G. (2022). Effects of vessel traffic and ocean noise on gray whale stress hormones. Scientific Reports 12:18580.
Mouton, T.L., Stephenson, F., Torres, L.G., Rayment, W., Brough, T., McLean, M., Tonkin, J.D., Albouy, C., Leprieur, F. (2022). Spatial mismatch in diversity facets reveals contrasting protection for New Zealand’s cetacean biodiversity. Biological Conservation 267:109484. https://doi.org/10.1016/j.biocon.2022.109484
Nazario, E.C., Cade, D.E., Bierlich, K.C., Czapanskiy, M.F., Goldbogen, J.A., Kahane-Rapport, S.R., van der Hoop, J.M., San Luis, M.T., Friedlaender, A.S. (2022). Baleen whale inhalation variability revealed using animal-borne video tags. PeerJ 10:e13724 https://doi.org/10.7717/peerj.13724
Pallin, L., Bierlich, K.C., Durban, J. Fearnbach, H., Savenko, O., C.S. Baker, E. Bell, Double, M.C., de la Mare, W., Goldbogen, J., Johnston, D., Kellar, N., Nichols, R., Nowacek, D., Read, A.J., Steel, D., Friedlaender, A. (2022) Demography of an ice-obligate mysticete in a region of rapid environmental change. Royal Society of Open Science. 9(11). https://doi.org/10.1098/rsos.220724
Reisinger, R.R., Brooks, C.M., Raymond, B., …, Torres, L.G., et al. (2022). Predator-derived bioregions in the Southern Ocean: Characteristics, drivers and representation in marine protected areas. Biological Conservation 272:109630. https://doi.org/10.1016/j.biocon.2022.109630
Rivers, J.W., Guerrero J.B., Brodeur, R.D., …, Torres. L.G., Barth, J.A. (2022). Critical research needs for forage fish within inner shelf marine ecosystems. Fisheries 47(5):213-221. https://doi.org/10.1002/fsh.10725
Obtaining enough food is crucial for predators to ensure adequate energy gain for maintenance of vital functions and support for energetically costly life history events (e.g., reproduction). Foraging involves decisions at every step of the process, including prey selection, capture, and consumption, all of which should be as efficient as possible. Making poor foraging decisions can have long-term repercussions on reproductive success and population dynamics (Harris et al. 2007, 2008, Grémillet et al. 2008), and for marine predators that rely on prey that is spatially and temporally dynamic and notoriously patchy (Hyrenbach et al. 2000), these decisions can be especially challenging. Prey abundance and density are frequently used as predictors of marine predator distribution, movement, and foraging effort, with predators often selecting highly abundant or dense prey patches (e.g., Goldbogen et al. 2011, Torres et al. 2020). However, there is increased recognition that prey quality is also an important factor to consider when assessing a predator’s ecology and habitat use (Spitz et al. 2012), and marine predators do show a preference for higher quality prey items (e.g., Haug et al. 2002, Cade et al. 2022). Moreover, negative impacts of low-quality prey on the health and breeding success of some marine mammals (Rosen & Trites 2000, Trites & Donnelly 2003) have been documented. Therefore, examining multiple prey metrics, such as prey quantity and quality, in predator ecology studies is critical.
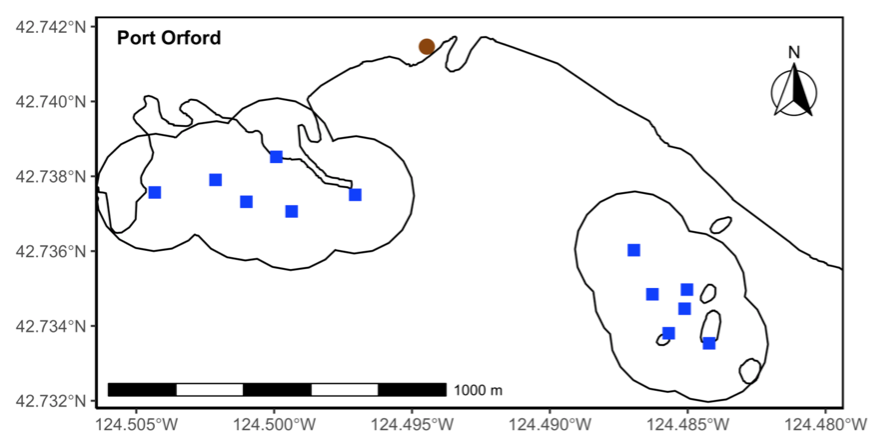
Figure 1. Site map of the Port Orford TOPAZ/JASPER integrated projects. Blue squares represent the location of the 12 sampling stations within the 2 study sites (site boundaries demarcated with black lines). Brown dot represents the cliff-top observation site where theodolite tracking occurred.
Our integrated TOPAZ/JASPER projects in Port Orford do just this! We collect both prey quantity and quality data from a tandem research kayak, while we track Pacific Coast Feeding Group (PCFG) gray whales from shore. The prey and whale sampling overlap spatially (and often temporally within the same day). This kind of concurrent predator-prey sampling at similar scales is often logistically challenging to achieve, yet because PCFG gray whales have an affinity for nearshore, coastal habitats, it is something we have been able to achieve in Port Orford. Since 2016, a field team comprised of graduate, undergraduate, and high school students has collected data during the month of August to investigate gray whale foraging decisions relative to prey. Every day, a kayak team collects GoPro videos (to assess relative prey abundance; AKA: quantity) and zooplankton samples using a tow net (to assess prey community composition; AKA: quality through caloric content of different species) (Figure 1). At the same time, a cliff team surveys for gray whales from shore and tracks them using a theodolite, which provides us with tracklines of individual whales; We categorize each location of a whale into three broad behavior states (feeding, searching, transiting) based on movement patterns. Over the years, the various students who have participated in the TOPAZ/JASPER projects have written many blog posts, which I encourage you to read here (particularly to get more detailed information about the field methods).
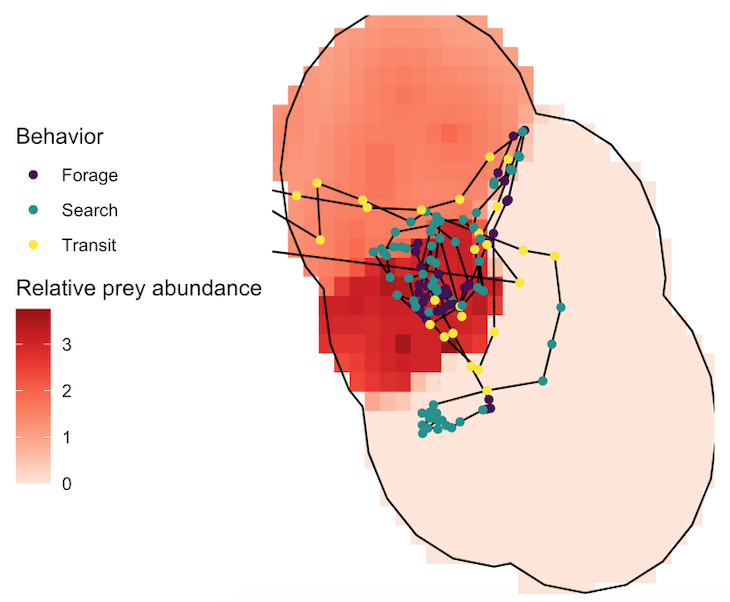
Figure 2. An example daily layer of relative prey abundance (increasing color darkness corresponds with increasing abundance) in one study site with a whale theodolite trackline recorded on the same day overlaid and color-coded by behavioral state.
Several years of data are needed to conduct a robust analysis for our ecological questions about prey choice, but after seven years, we finally had the data and I am excited to share the results, which are due to the many years of hard work from many students! Our recent paper in Marine Ecology Progress Series aimed to determine whether PCFG gray whale foraging decisions are driven by prey quantity (abundance) or quality (caloric content of species) at a scale of 20 m (which is slightly less than 2 adult gray whale body lengths). In this study, we built upon results from my previous Master’s publication, which revealed that there are significant differences in the caloric content between the six common nearshore zooplankton prey species that PCFG gray whales feed on (Hildebrand et al. 2021). Therefore, in this study we addressed the hypothesis that individual whales will select areas where the prey community is dominated by the mysid shrimp Neomysis rayii, since it is significantly higher in caloric content than the other two prey species we identified, Holmesimysis sculpta (a medium quality mysid shrimp species) and Atylus tridens (a low quality amphipod species) (Hildebrand et al. 2021). We used spatial statistics and model to make daily maps of prey abundance and quality that we compared to our whale tracks and behavior from the same day. Please read our paper for the details on our novel methods that produced a dizzying amount of prey layers, which allowed us to tease apart whether gray whales target prey quantity, quality, or a mixture of both when they forage.
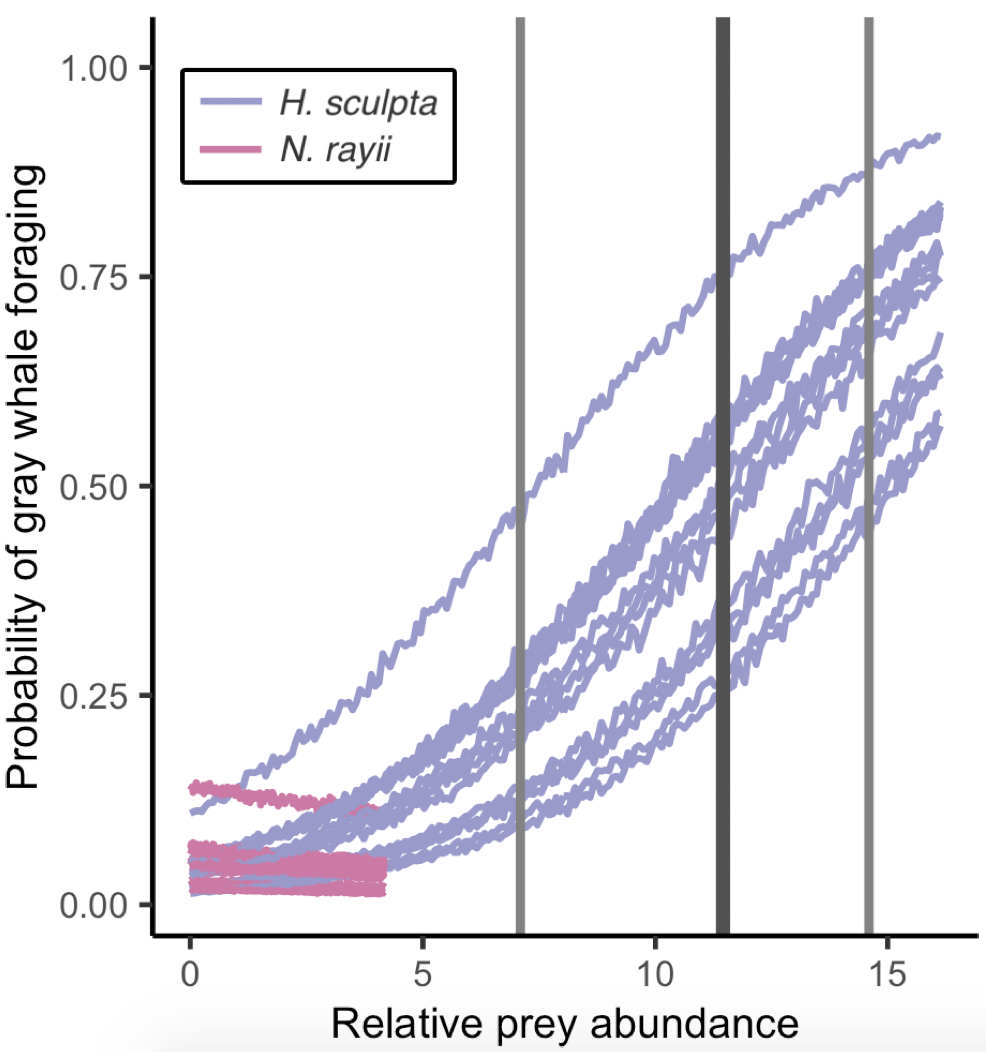
Figure 3. Figure shows the probability of gray whale foraging relative to prey abundance (color-coded by prey species). Dark grey vertical line represents the mean threshold for the H. sculpta curves (12.0); light grey vertical lines: minimum (7.2) and maximum (15.3) thresholds for the H. sculpta curves. Inflection points could not be calculated for the N. rayii curves
So, what did we find? The models proved our hypothesis wrong: foraging probability was significantly correlated with the quantity and quality of the mysid H. sculpta, which has significantly lower calories than N. rayii. This result puzzled us, until we started looking at the overall quantity of these two prey types in the study area and realized that the amount of calorically-rich N. rayii never reached a threshold where it was beneficial for gray whales to forage. But, there was a lot of H. sculpta, which likely made for an energetic gain for the whales despite not being as calorically rich as N. rayii. We determined a threshold of H. sculpta relative abundance that is required to initiate the gray whale foraging behavior, and the abundance of N. rayii never came close to this level (Figure 3). Despite not having the highest quality, H. sculpta did have the highest abundance and showed a significant positive relationship with foraging behavior, unlike the other prey items. Interestingly, whales never selected areas dominated by the low-calorie species A. tridens. These results demonstrate trade-off choices by whales for this abundant, medium-quality prey.
To our knowledge, individual baleen whale foraging decisions relative to available prey quantity and quality have not been addressed previously at this very fine-scale. Interestingly, this trade-off between prey quantity and quality has also been detected in humpback whales foraging in Antarctica at depths deeper than where the densest krill patches occur; while the whales are exploiting less dense krill patches, these krill composed of larger, gravid, higher-quality krill (Cade et al. 2022). While it is unclear how baleen whales differentiate between prey species or reproductive stages, several mechanisms have been suggested, including visual and auditory identification (Torres 2017). We assume here that gray whales, and other baleen whale species, can differentiate between prey species. Thus, our results showcase the importance of knowing the quality (such as caloric content) of prey items available to predators to understand their foraging ecology (Spitz et al. 2012).
References
Cade DE, Kahane-Rapport SR, Wallis B, Goldbogen JA, Friedlaender AS (2022) Evidence for size-selective pre- dation by Antarctic humpback whales. Front Mar Sci 9:747788
Goldbogen JA, Calambokidis J, Oleson E, Potvin J, Pyenson ND, Schorr G, Shadwick RE (2011) Mechanics, hydrody- namics and energetics of blue whale lunge feeding: effi- ciency dependence on krill density. J Exp Biol 214:131−146
Grémillet D, Pichegru L, Kuntz G, Woakes AG, Wilkinson S, Crawford RJM, Ryan PG (2008) A junk-food hypothesis for gannets feeding on fishery waste. Proc R Soc B 275: 1149−1156
Harris MP, Beare D, Toresen R, Nøttestad L, and others (2007) A major increase in snake pipefish (Entelurus aequoreus) in northern European seas since 2003: poten- tial implications for seabird breeding success. Mar Biol 151:973−983
Harris MP, Newell M, Daunt F, Speakman JR, Wanless S (2008) Snake pipefish Entelurus aequoreus are poor food for seabirds. Ibis 150:413−415
Haug T, Lindstrøm U, Nilssen KT (2002) Variations in minke whale (Balaenoptera acutorostrata) diet and body condi- tion in response to ecosystem changes in the Barents Sea. Sarsia 87:409−422
Hildebrand L, Bernard KS, Torres LG (2021) Do gray whales count calories? Comparing energetic values of gray whale prey across two different feeding grounds in the eastern North Pacific. Front Mar Sci 8:1008
Hyrenbach KD, Forney KA, Dayton PK (2000) Marine pro- tected areas and ocean basin management. Aquat Con- serv 10:437−458
Rosen DAS, Trites AW (2000) Pollock and the decline of Steller sea lions: testing the junk-food hypothesis. Can J Zool 78:1243−1250
Spitz J, Trites AW, Becquet V, Brind’Amour A, Cherel Y, Galois R, Ridoux V (2012) Cost of living dictates what whales, dolphins and porpoises eat: the importance of prey quality on predator foraging strategies. PLOS ONE 7:e50096
Torres LG, Barlow DR, Chandler TE, Burnett JD (2020) Insight into the kinematics of blue whale surface forag- ing through drone observations and prey data. PeerJ 8: e8906
Torres LG (2017) A sense of scale: foraging cetaceans’ use of scale-dependent multimodal sensory systems. Mar Mamm Sci 33:1170−1193
Trites AW, Donnelly CP (2003) The decline of Steller sea lions Eumetopias jubatus in Alaska: a review of the nutri- tional stress hypothesis. Mammal Rev 33:3−28
By Braden Adam Vigil, Oregon State University Undergraduate, GEMM Lab NSF REU Intern
Introduction
My name is Braden Vigil, and I am enjoying this summer with the company of Lisa Hildebrand and Dr. Leigh Torres as a National Science Foundation (NSF) Research Experience for Undergraduates (REU) intern. By slicing off a manageable chunk of the GRANITE project to focus on, I’ve had the chance to explore my passion for data visualization. My excitement for biological research was instilled in me by an impactful high school biology teacher (thank you Mr. Villalobos!) and was narrowed to marine biology research after a chance visit to Oregon State University’s Hatfield Marine Science Center. I’ve come all the way from Southern New Mexico to explore this passion of mine, and the REU program has been one of my first chances to get my feet wet. My advice for any students debating taking big leaps for the sake of passion is to do it – it’s scary, but I’d say there’s nothing better than living out what you want to do (and hopefully getting paid for it!). For this project, the GEMM Lab has saved me the trouble of collecting data – this summer, I’m all action.
Where Gray Whales Are and Why It Matters
Just as you might find yourself at a grocery store to buy food or at a coffee shop catching up with an old friend, so too do whales have places to go and reasons for being there. Research concerning gray whale ecology – understanding the who, what, when, where, whys – should then have a lot to do with the “where?” and “why?” That’s what my project is about: investigating where the gray whales off the Oregon coast are, and what features of the environment are related to their presence and other aspects of the population. After all, distribution is considered the foundational unit for the biogeographical understanding of a population’s location and its interactions with other species. An example of an environmental driver may be phytoplankton and – subsequently – zooplankton abundance. It’s been shown that bottom-up trophic cascades based on primary productivity directly influence predator and prey populations in both terrestrial and marine ecosystems (Sinclair and Krebs 2002; Benoit-Bird and McManus 2012). This driver specifically could then inform something as significant as population abundance of a predator, though that’s out of the scope of my project. Instead, I’m studying how these environmental drivers, specifically sea water temperature, affects the variation of the thyroid hormone (tri-iodothyronine, T3) in gray whales, which the GEMM Lab quantifies from fecal samples that they non-invasively and opportunistically collect. In terrestrial mammals, T3 is believed to be associated with thermoregulation, yet it is unclear if T3 has the same function in baleen whales who use blubber insulation to thermoregulate. To estimate blubber insulation, we use a proxy, called body area index (BAI) collected via drone footage (Burnett et al. 2018), which you can read more about in Clara’s blog. Insights into variations in T3 hormone levels as related to changes in the environment may allow researchers to better understand thermoregulatory challenges whales face in warming oceans.
This Sounds Like a Lot of Data About the Environment, Where’s it Coming From?
Not only has the GEMM Lab relieved me of the hassle that data collection and fieldwork can be, so too has the Ocean Observatories Initiative (OOI). Starting in 2014, the OOI has set up several buoys off the U.S. West Coast, each equipped with numerous sensors and data-collecting devices. These have been extracting data from the nearby environment since then, including aspects such as dissolved oxygen, pH, and most important to this study, sea temperature. These buoys run deep too! Some devices reach as low as 25 m, which is where we often expect to see whales foraging during surveys. For our interest, there is one specific buoy that is within the GRANITE project’s survey region, the Oregon Inshore Surface Mooring.
Figure 1. Locations of OOI buoys. Blue dots represent buoys, while the yellow dot represents our buoy of interest, the Oregon Inshore Surface Mooring.
Expectations
The OOI has published, and continues to publish, an unbelievable amount of data. There are many things that would be interesting to investigate, but until we know how much we can bite off versus how much we can chew, we’ve narrowed it down to a few hypotheses we’re currently investigating.
Table 1. Hypotheses and Expected Results.
A Hundred and One Data Visualizations
As fun as I find testing correlations between variables and creating satisfying looking plots, I must admit that I’m not even halfway into this project and I’ve made a LOT of plots. Plots can be an easy way to understand big datasets and observations. Since not all of the data-collecting devices on the OOI data are continuously running, I first needed to get an idea of how much data we have to work with, and how much of that data overlaps in time with our annual gray whale survey period (June 1 – October 15). Some of these preliminary plots look like Figure 2. In addition, these plots grant us an idea of how variable sea surface temperatures have been in these past few years. Marine heatwaves have occurred recently in the Pacific Ocean and off the U.S. West Coast, and it is important to know if their effects continue to linger to the present. Other, unexplained peaks might also be worth investigating.
Figure 2. Preliminary plot comparing sea surface temperature data over time, from around June 2016 to December 2021. Straight lines between December to June each year indicates no data, as we have removed these periods from our analysis.
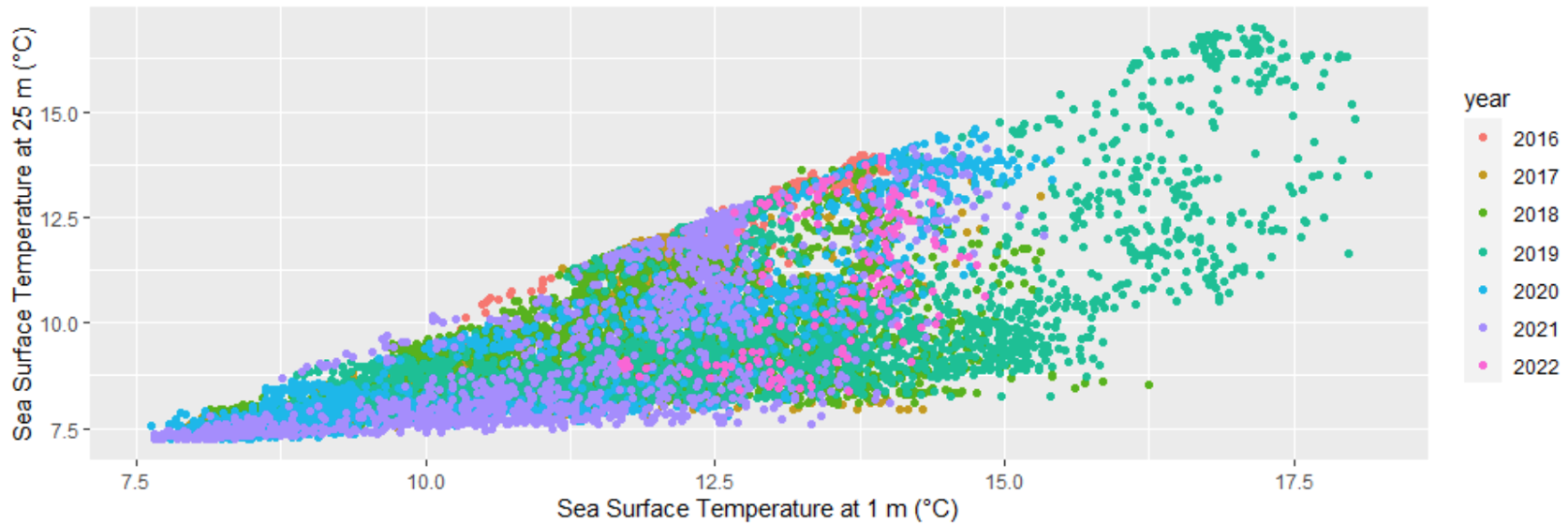
The goal here is to eventually compare the variables of sea temperature to the T3 hormone levels in gray whales foraging off the Oregon coast. Before this step, it is important to decide what depth of temperature readings are most appropriate to assess. I’ve made several correlation plots of sea temperature between varying depths of 1 m, 7 m, and 25 m. One such plot is included below (Figure 3). This plot shows variation of temperature between different depths. If there is strong variation between the depths of 1 m and 25 m, then the water column may be well stratified, meaning that gray whale response to environmental temperature may be distinct between these distances, possibly even between 1 m and 7 m.
Figure 3. Sea surface temperature at 1 m versus 25 m in degrees Celsius, with points color coded by year.
Conclusion
As previously described, this study plays part into the larger GRANITE project with the goal to understand and make predictions about the ecology and physiology of the gray whale population off of the U.S. West Coast. This study will investigate the significance of sea temperature on aspects of whale health – so far including BAI and T3 hormone level. I will be pursuing a stronger grasp on the variation of these relationships through ongoing analysis. My results should be used to clarify nodes and the correlation between them in the web of dynamics encircling the population. This project has given me great insight into how raw data can be turned into meaningful understandings and subsequent impacts. The public OOI data is a scattershot of many different measurements using many different devices constantly. The answers/solutions to the conservation of species threatened by the Anthropocene are out there, all that’s required is that we harness them.
References
Benoit-Bird, K. J., & McManus, M. A. (2012). Bottom-up regulation of a pelagic community through spatial aggregations. Biology Letters, 8(5), 813–816.https://doi.org/10.1098/rsbl.2012.0232
Burnett, J. D., & Wing, M. G. (2018). A low-cost near-infrared digital camera for fire detection and monitoring. International Journal of Remote Sensing, 39(3), 741–753.https://doi.org/10.1080/01431161.2017.1385109
Sinclair, A. R. E., & Krebs, C. J. (2002). Complex numerical responses to top–down and bottom–up processes in vertebrate populations. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 357(1425), 1221–1231.https://doi.org/10.1098/rstb.2002.1123.
We are almost halfway through June which means summer has arrived! Although, here on the Oregon coast, it does not entirely feel like it. We have been swinging between hot, sunny days and cloudy, foggy, rainy days that are reminiscent of those in spring or even winter. Despite these weather pendulums, the GEMM Lab’s GRANITE project is off to a great start in its 8th field season! The field team has already ventured out onto the Pacific Ocean in our trusty RHIB Ruby on four separate days looking for gray whales and in this blog post, I am going to share what we have seen so far.
The core GRANITE field team before the May 24th “trial run”. From left to right: Leigh Torres, KC Bierlich, Clara Bird, Lisa Hildebrand, Alejandro Fernández Ajó. Source: L. Torres.
PI Leigh, PhD candidate Clara and I headed out for a “trial run” on May 24th. While the intention for the day was to make sure all our gear was running smoothly and we still remembered how to complete the many tasks associated with our field work (boat loading and trailering, drone flying and catching, poop scooping, data download, to name a few), we could not resist surveying our entire study range given the excellent conditions. It was a day that all marine field scientists hope for – low winds (< 5 kt all day) and a 3 ft swell over a long period. Despite surveying between Waldport and Depoe Bay, we only encountered one whale, but it was a whale that put a smile on each of our faces. After “just” 252 days, we reunited with Solé, the star of our GRANITE dataset, with record numbers of fecal samples and drone flights collected. This record is due to what seems to be a strong habitat or foraging tactic preference by Solé to remain in a relatively small spatial area off the Oregon coast for most of the summer, rather than traveling great swaths of the coast in search for food. Honest truth, on May 24th we found her exactly where we expected to find her. While we did not collect a fecal sample from her on that day, we did perform a drone flight, allowing us to collect a critical early feeding season data point on body condition. We hope that Solé has a summer full of mysids on the Oregon coast and that we will be seeing her often, getting rounder each time!
Our superstar whale Solé. Her identifying features are a small white line on her left side (green box) and a white dot in front of her dorsal hump on the right side (red circle). Source: GEMM Lab. Photograph captured under NOAA/NMFS permit #21678
Just a week after this trial day, we had our official start to the field season with back-to-back days on the water. On our first day, postdoc Alejandro, Clara and I were joined by St. Andrews University Research Fellow Enrico Pirotta, who is another member of the GRANITE team. Enrico’s role in the GRANITE project is to implement our long-term, replicate dataset into a framework called Population consequences of disturbance (PCoD; you can read all about it in a previous blog). We were thrilled that Enrico was able to join us on the water to get a sense for the species and system that he has spent the last several months trying to understand and model quantitatively from a computer halfway across the world. Luckily, the whales sure showed up for Enrico, as we saw a total of seven whales, all of which were known individuals to us! Several of the whales were feeding in water about 20 m deep and surfacing quite erratically, making it hard to get photos of them at times. Our on-board fish finder suggested that there was a mid-water column prey layer that was between 5-7 m thick. Given the flat, sandy substrate the whales were in, we predicted that these layers were composed of porcelain crab larvae. Luckily, we were able to confirm our hypothesis immediately by dropping a zooplankton net to collect a sample of many porcelain crab larvae. Porcelain crab larvae have some of the lowest caloric values of the nearshore zooplankton species that gray whales likely feed on (Hildebrand et al. 2021). Yet, the density of larvae in these thick layers probably made them a very profitable meal, which is likely the reason that we saw another five whales the next day feeding on porcelain crab larvae once again.
Porcelain crab larvae. Source: GEMM Lab.A happy captain Ale! Source: GEMM Lab.Enrico (right) and myself after collecting a large fecal sample. Source: GEMM Lab.
On our most recent field work day, we only encountered Solé, suggesting that the porcelain crab swarms had dissipated (or had been excessively munched on by gray whales), and many whales went in search for food elsewhere. We have done a number of zooplankton net tows across our study area and while we did collect a good amount of mysid shrimp already, they were all relatively small. My prediction is that once these mysids grow to a more profitable size in a few days or weeks, we will start seeing more whales again.
The GRANITE team from above, waiting & watching for whales, as we will be doing for the rest of the summer! Source: GEMM Lab.
So far we have seen nine unique individuals, flown the drone over eight of them, collected fecal samples from five individuals, conducted 10 zooplankton net tows and seven GoPro drops in just four days of field work! We are certainly off to a strong start and we are excited to continue collecting rock solid GRANITE data this summer to continue our efforts to understand gray whale ecology and physiology.
Literature cited
Hildebrand L, Bernard KS, Torres LGT. 2021. Do gray whales count calories? Comparing energetic values of gray whale prey across two different feeding grounds in the Eastern North Pacific. Frontiers in Marine Science 8. doi: 10.3389/fmars.2021.683634
In September 2020, I was hired as a postdoc in the GEMM Lab and was tasked to conduct the analyses necessary for the OPAL project. This research project has the ambitious, yet essential, goal to fill a knowledge gap hindering whale conservation efforts locally: where and when do whales occur off the Oregon coast? Understanding and predicting whale distribution based on changing environmental conditions is a key strategy to assess and reduce spatial conflicts with human activities, specifically the risk of entanglement in fixed fishing gear.
Starting a new project is always a little daunting. Learning about a new region and new species, in an alien research and conservation context, is a challenge. As I have specialized in data science over the last couple of years, I have been confronted many times with the prospect of working with massive datasets collected by others, from which I was asked to tease apart the biases and the ecological patterns. In fact, I have come to love that part of my job: diving down the data rabbit hole and making my way through it by collaborating with others. Craig Hayslip, faculty research assistant in MMI, was the observer who conducted the majority of the 102 helicopter surveys that were used for this study. During the analysis stage, his help was crucial to understand the data that had been collected and get a better grasp of the field work biases that I would later have to account for in my models. Similarly, it took hours of zoom discussions with Dawn Barlow, the GEMM lab’s latest Dr, to be able to clean and process the 75 days of survey effort conducted at sea, aboard the R/V Shimada and Oceanus.
Once the data is “clean”, then comes the time for modeling. Running hundreds of models, with different statistical approaches, different environmental predictors, different parameters etc. etc. That is when you realize what a blessing it is to work with a supervisor like Leigh Torres, head of the GEMM Lab. As an early career researcher, I really appreciate working with people who help me take a step back and see the bigger picture within which the whole data wrangling work is included. It is so important to have someone help you stay focused on your goals and the ecological questions you are trying to answer, as these may easily get pushed back to the background during the data analysis process.
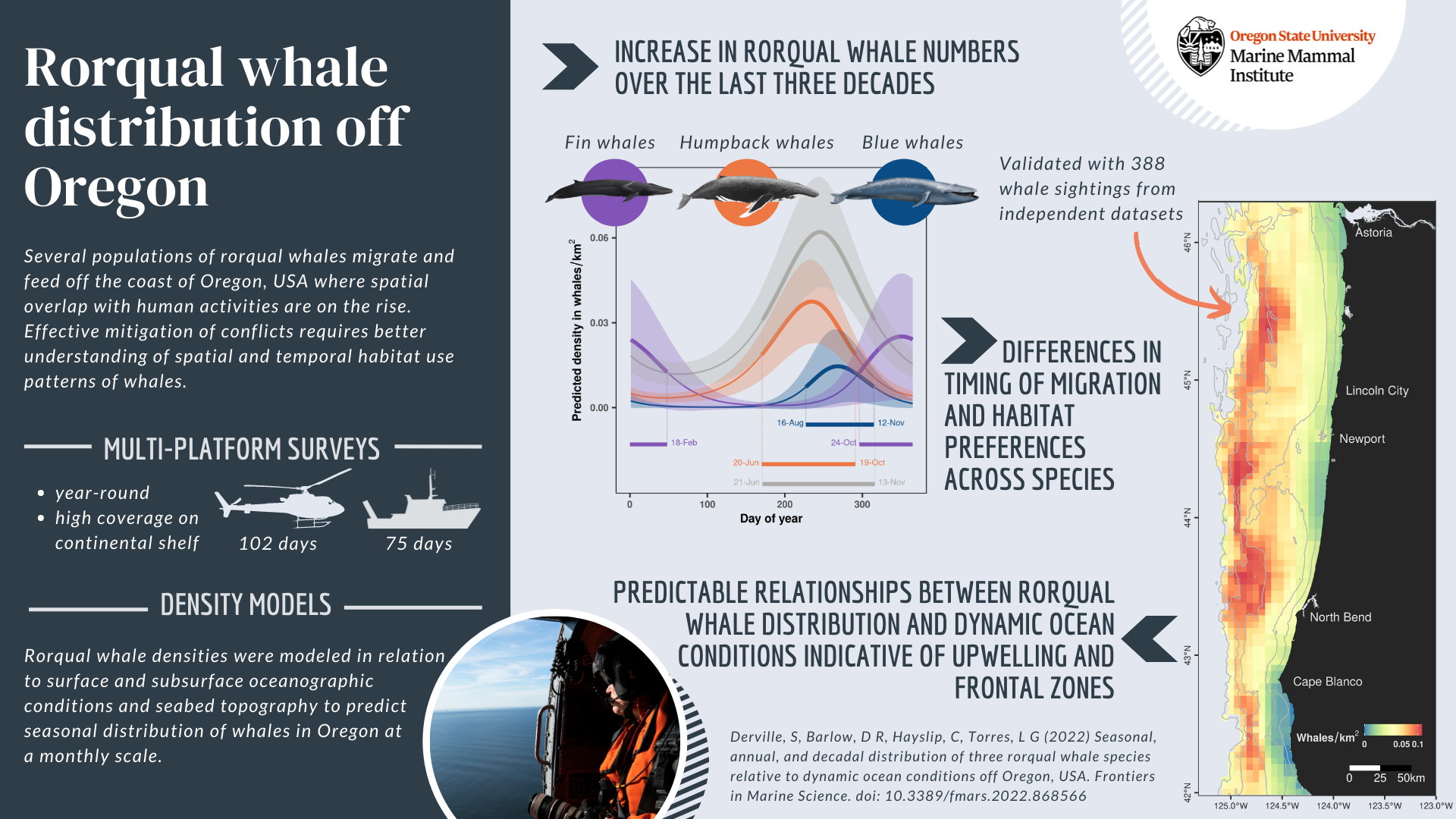
And here we are today, with the first scientific publication from the OPAL project published, a little more than three years after Leigh and Craig started collecting data onboard the United States Coast Guard helicopters off the coast of Oregon in February 2019. Entitled “Seasonal, annual, and decadal distribution of three rorqual whale species relative to dynamic ocean conditions off Oregon, USA”, our study published in Frontiers in Marine Science presents modern and fine-scale predictions of rorqual whale distribution off Oregon, as well as a description of their phenology and a comparison to whale numbers observed across three decades in the region (Figure 1). This research focuses on three rorqual species sharing some ecological and biological traits, as well as similar conservation status: humpback whales (Megaptera novaeangliae), blue whales (Balaenoptera musculus musculus), and fin whales (Balaenoptera physallus); all of which migrate and feed over the US West coast (see a previous blog to learn more about these species here).
Figure 1: Graphical abstract of our latest paper published in Frontiers in Marine Science.
We demonstrate (1) an increase in rorqual numbers over the last three decades in Oregon waters, (2) differences in timing of migration and habitat preferences between humpback, blue, and fin whales, and (3) predictable relationships of rorqual whale distribution based on dynamic ocean conditions indicative of upwellings and frontal zones. Indeed, these ocean conditions are likely to provide suitable biological conditions triggering increased prey abundance. Three seasonal models covering the months of December-March (winter model), April-July (spring) and August-November (summer-fall) were generated to predict rorqual whale densities over the Oregon continental shelf (in waters up to 1,500 m deep). As a result, maps of whale densities can be produced on a weekly basis at a resolution of 5 km, which is a scale that will facilitate targeted management of human activities in Oregon. In addition, species-specific models were also produced over the period of high occurrence in the region; that is humpback and blue whales between April and November, and fin whales between August and March.
As we outline in our concluding remarks, this work is not to be considered an end-point, but rather a stepping stone to improve ecological knowledge and produce operational outputs that can be used effectively by managers and stakeholders to prevent spatial conflict between whales and human activities. As of today, the models of fin and blue whale densities are limited by the small number of observations of these two species over the Oregon continental shelf. Yet, we hope that continued data collection via fruitful research partnerships will allow us to improve the robustness of these species-specific predictions in the future. On the other hand, the rorqual models are considered sufficiently robust to continue into the next phase of the OPAL project that aims to assess overlap between whale distribution and Dungeness crab fishing gear to estimate entanglement risk.
The curse (or perhaps the beauty?) of species distribution modeling is that it never ends. There are always new data to be added, new statistical approaches to be tested, and new predictions to be made. The OPAL models are no exception to this rule. They are meant to be improved in future years, thanks to continued helicopter and ship-based survey efforts, and to the addition of new environmental variables meant to better predict whale habitat selection. For instance, Rachel Kaplan’s PhD research specifically aims at understanding the distribution of whales in relation to krill. Her results will feed into the more general efforts to model and predict whale distribution to inform management in Oregon.
This first publication therefore paves the way for more exciting and impactful research!
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly message when we post a new blog. Just add your name and email into the subscribe box below.
Reference
Derville, S., Barlow, D. R., Hayslip, C. E., and Torres, L. G. (2022). Seasonal, Annual, and Decadal Distribution of Three Rorqual Whale Species Relative to Dynamic Ocean Conditions Off Oregon, USA. Front. Mar. Sci. 9, 1–19. doi:10.3389/fmars.2022.868566.
Acknowledgments
We gratefully acknowledge the immense contribution of the United State Coast Guard sectors North Bend and Columbia River who facilitated and piloted our helicopter surveys. We would like to also thank NOAA Northwest Fisheries Science Center for the ship time aboard the R/V Bell M. Shimada. We thank the R/V Bell M. Shimada (chief scientists J. Fisher and S. Zeman) and R/V Oceanus crews, as well as the marine mammal observers F. Sullivan, C. Bird and R. Kaplan. We give special recognition and thanks to the late Alexa Kownacki who contributed so much in the field and to our lives. We also thank T. Buell and K. Corbett (ODFW) for their partnership over the OPAL project. We thank G. Green and J. Brueggeman (Minerals Management Service), J. Adams (US Geological Survey), J. Jahncke (Point blue Conservation), S. Benson (NOAA-South West Fisheries Science Center), and L. Ballance (Oregon State University) for sharing validation data. We thank J. Calambokidis (Cascadia Research Collective) for sharing validation data and for logistical support of the project. We thank A. Virgili for sharing advice and custom codes to produce detection functions.
When I was younger, I aspired to be a marine mammal biologist. I thought it was purely about knowing as much about marine mammal species as possible. However, over time and with experience in this field, I have realized that in order to understand a species, you need to have a holistic understanding of its prey, habitat, and environment. When I first applied to be advised by Leigh in the GEMM Lab, I had no idea how much of my time I would spend looking at tiny zooplankton under a microscope, thinking about the different benefits of different habitat types, or reading about oceanographic processes. But these things have been incredibly vital to my research to date and as a result, I now refer to myself as a marine ecologist. This holistic understanding that I am gaining will only grow throughout my PhD as I am broadly looking at the habitat use, site fidelity, and population dynamics of the Pacific Coast Feeding Group (PCFG) of gray whales for my thesis research.
The PCFG display many foraging tactics and occupy several habitat types along the Oregon coast while they spend their summer feeding seasons here (Torres et al. 2018). Here, I will focus on one of these habitats: kelp. When you hear the word kelp, you probably conjure an image of long, thick stalks that reach from the ocean floor to the surface, with billowing fronds waving around (Figure 1a). However, this type is only one of three basic morphologies (Filbee-Dexter & Scheibling 2014) and it is called canopy kelp, which often forms extensive forests. The other two morphologies are stipitate and prostrate kelps. The former forms midwater stands (Figure 1b) while the latter forms low-lying kelp beds (Figure 1c). All three of these morphologies exist on the Oregon coast and create a mosaic of understory and canopy kelp patches that dot our coastline.
Figure 1. Examples of the three different kelp morphologies. a: bull kelp (Nereocystis luetkeana) is a type of canopy kelp and the dominant kelp on the Oregon coast (Source: Oregon Coast Aquarium); b: sea palm (Postelsia palmaeformis) is a type of stipitate kelp that forms mid-water stands (Source: Oregon Conservation Strategy); c: sea cabbage (Saccharina sessilis) is a type of prostrate kelp that is stipeless and forms low-lying kelp beds (Source: Central Coast Biodiversity).
One of the most magnificent things about kelp is that it is not just a species itself, but it provides critical habitat, refuge, and food resources to a myriad of other species due to its high rates of primary production (Dayton 1985). Kelp is often referred to as a foundation species due to all of these critical services it provides. In Oregon, many species of rockfish, which are important commercial and recreational fisheries, use kelp as habitat throughout their life cycle, including as nursery grounds. Lingcod, another widely fished species, forages amongst kelp. A large number of macroinvertebrates can be found in Oregon kelp forests, including anemones, limpets, snails, sea urchins, sea stars, and abalone, to name a fraction of them.
Kelps grow best in cold, nutrient-rich waters (Tegner et al. 1996) and their growth and distribution patterns are highly naturally variable on both temporal and spatial scales (Krumhansl et al. 2016). However, warm water, low nutrient or light conditions, intensive grazing by herbivores, and severe storm activity can lead to the erosion and defoliation of kelp beds (Krumhansl et al. 2016). While these events can occur naturally in cyclical patterns, the frequency of several of these events has increased in recent years, as a result of climate change and anthropogenic impacts. For example, Dawn’s blog discussed increasing marine heatwaves that represent an influx of warm water for a prolonged period of time. In fact, kelps can be useful sentinels of change as they tend to be highly responsive to changes in environmental conditions (e.g., Rogers-Bennet & Catton 2019) and their nearshore, coastal location directly exposes them to human activities, such as pollution, harvesting, and fishing (Bennett et al. 2016).
Due to its foundational role, changes or impacts to kelp can reverberate throughout the ecosystem and negatively affect many other species. As mentioned previously, kelp is naturally highly variable, and like many other ecological processes, undergoes boom and bust cycles. For over four decades, dense, productive kelp forests have been shown to transition to sea urchin barrens, and back again, in natural cycles (Sala et al. 1998;Pinnegar et al. 2000;Steneck et al. 2002; Figure 2). These transitions are called phase shifts. In a healthy, balanced kelp forest, sea urchins typically passively feed on detrital plant matter, such as broken off pieces of kelp fronds that fall to the seafloor. A phase shift occurs when the grazing intensity of sea urchins increases, resulting in them actively feeding on kelp stalks and fronds to a point where the kelp in an area can become greatly reduced, creating an urchin barren. Sea urchin grazing intensity can change for a number of reasons, including reduction in sea urchin predators (e.g., sea otters, sunflower sea stars) or poor kelp recruitment events (e.g., due to warm water temperature). Regardless of the reason, the phases tend to transition back and forth over time. However, there is concern that sea urchin barrens may become an alternative stable state of the subtidal ecosystem from which kelp in an area cannot recover (Filbee-Dexter & Scheibling 2014).
Figure 2. Screenshots from GoPro videos from 2016 (left) and 2018 (right) at the same kayak sampling station in Port Orford showing the difference between a dense kelp forest and what appears to be an urchin barren. (Source: GEMM Lab).
For example, in 2014, bull kelp canopy cover in northern California was reduced by >90% and has not shown signs of recovery since (Rogers-Bennet & Catton 2019; Figure 3). This massive decline was attributed to two major events: 1) the onset of sea star wasting disease (SSWD) in 2013 and 2) the “warm blob” of 2014-2016. SSWD affected over 20 sea star species along the coast from Mexico to Alaska, with the predatory sunflower sea star, which consumes purple sea urchins, most affected, including population declines of 80-100% along the coast (Harvell et al. 2019). Following this SSWD outbreak, the “warm blob”, which was an extreme marine heatwave in the Pacific Ocean, caused ocean temperatures to spike. These two events allowed purple sea urchin populations to grow unchecked by their predators, and created nutrient-poor and warm water conditions, which limited kelp growth and productivity. Intense grazing on bull kelp by growing urchin populations resulted in the >90% reduction in bull kelp canopy cover and has left behind widespread urchin barrens instead (Rogers-Bennet & Catton 2019). Consequently, there have been ecological and economic impacts on the ecosystem and communities in northern California. Without bull kelp, red abalone and red sea urchin populations starved, leading to a subsequent loss of the recreational red abalone (estimated value of $44 million/year) and commercial red urchin fisheries in northern California (Rogers-Bennet & Catton 2019).
Figure 3. Surface kelp canopy area pre- and post-impact from sites in Sonoma and Mendocino counties, northern California from aerial surveys (2008, 2014-2016). Figure and figure caption taken from Rogers-Bennett & Catton (2019).
As I mentioned earlier, while phase shifts between kelp forests and urchin barrens are common cycles, the intensity of the events described above in northern California are an example of sea urchin barrens potentially becoming a stable state of the subtidal ecosystem (Filbee-Dexter & Scheibling 2014). Given that marine heatwaves are only expected to increase in intensity and frequency in the future (Frölicher et al. 2018), the events documented in northern California may not be an isolated incidence.
Considering that parts of the Oregon coast, particularly the southern portion, are very similar to northern California biogeographically, and that it was not exempt from the “warm blob”, similar changes in kelp forests may be occurring along our coast. There are many individuals and groups that are actively working on this issue to examine potential impacts to kelp and the species that depend on the services it provides. For more information, check out the Oregon Kelp Alliance.
Figure 4. A gray whale surfaces in a large kelp bed during a foraging bout along the Oregon coast. (Source: GEMM Lab).
So, what does all of this information have to do with gray whales? Given their affinity for kelp habitats (Figure 4) and their zooplankton prey that aggregates there, changes to kelp ecosystems may affect gray whale health and ecology. This aspect of the complex kelp trophic web has not been examined to date; thus one of my PhD chapters focuses on the response of gray whales to changing kelp ecosystems along the southern Oregon coast. To do this, I am examining 6 years of data collected during the TOPAZ/JASPER project in Port Orford, to look at the relationships between kelp health, sea urchin density, zooplankton abundance, and gray whale foraging effort over space and time. Documenting impacts of changing kelp forests on gray whales is important to assist management efforts as healthy and abundant kelp seems critical in providing ample food opportunities for these iconic Pacific Northwest marine predators.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly alert when we make a new post! Just add your name into the subscribe box on the left panel.
References
Bennett S, et al. The ‘Great Southern Reef’: Social, ecological and economic value of Australia’s neglected kelp forests. Marine and Freshwater Research 67:47-56.
Dayton PK (1985) Ecology of kelp communities. Annual Review of Ecology and Systematics 16:215-245.
Filbee-Dexter K, Scheibling RE (2014) Sea uechin barrens as alternative stable states of collapsed kelp ecosystems. Marine Ecology Progress Series 495:1-25.
Frölicher TL, Fischer EM, Gruber N (2018) Marine heatwaves under global warming. Nature 560:360-364.
Harvell CD, et al. (2019) Disease epidemic and a marine heat wave are associated with the continental-scale collapse of a pivotal predator (Pycnopodia helianthoides). Science Advances 5(1) doi:10.1126/sciadv.aau7042.
Krumhansl KA, et al. (2016) Global patterns of kelp forest change over the past half-century. Proceedings of the National Academy of Sciences of the United States of America 113(48):13785-13790.
Pinnegar JK, et al. (2000) Trophic cascades in benthic marine ecosystems: lessons for fisheries and protected-area management. Environmental Conservation 27:179-200.
Rogers-Bennett L, Catton CA (2019) Marine heat wave and multiple stressors tip bull kelp forest to sea urchin barrens. Scientific Reports 9:15050.
Sala E, Boudouresque CF, Harmelin-Vivien M (1998) Fishing, trophic cascades and the structure of algal assemblages; evaluation of an old but untested paradigm. Oikos 82:425-439.
Steneck RS, et al. (2002) Kelp forest ecosystems: biodiversity, stability, resilience and future. Environmental Conservation 29:436-459.
Tegner MJ, Dayton PK, Edwards PB, Riser KL (1996) Is there evidence for the long-term climatic change in southern California kelp forests? California Cooperative Oceanic Fisheries Investigations Report 37:111-126.
Torres LG, Nieukirk SL, Lemos L, Chandler TE (2018) Drone up! Quantifying whale behavior from a new perspective improves observational capacity. Frontiers in Marine Science doi:10.3389/fmars.2019.00319.
By Allison Dawn, GEMM Lab Master’s student, OSU Department of Fisheries, Wildlife, and Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
During my second term of graduate school, I have been preparing to write my research proposal. The last two months have been an inspiring process of deep literature dives and brainstorming sessions with my mentors. As I discussed in my last blog, I am interested in questions related to pattern and scale (fine vs. mesoscale) in the context of the Pacific Coast Feeding Group (PCFG) of gray whales, their zooplankton prey, and local environmental variables.
My work currently involves exploring which scales of pattern are important in these trophic relationships and whether the dominant scale of a pattern changes over time or space. I have researched which analysis tools would be most appropriate to analyze ecological time series data, like the impressive long-term dataset the GEMM lab has collected in Port Orford as part of the TOPAZ project, where we have monitored the abundance of whales and zooplankton, as well as environmental variables since 2016.
A useful analytical tool that I have come across in my recent coursework and literature review is called wavelet analysis. Importantly, wavelet analysis can handle non-stationarity and edge detection in time series data. Non-stationarity is when a dataset’s mean and/or variance can change over time or space, and edge detection is the identification of the change location (in time or space). For example, it is not just the cycles or “ups and downs” of zooplankton abundance I am interested in, but when in time or where in space these cycles of “ups and downs” might change in relation to what their previous values, or distances between values, were. Simply stated, non-stationarity is when what once was normal is no longer normal. Wavelet analysis has been applied across a broad range of fields, such as environmental engineering (Salas et al. 2020), climate science (Slater et al. 2021), and bio-acoustics (Buchan et al. 2021). It can be applied to any time series dataset that might violate the traditional statistical assumption of stationarity.
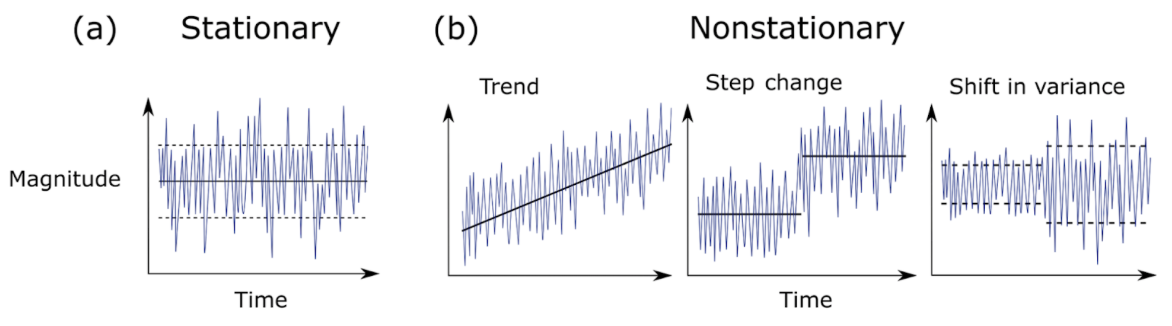
In a recent review of climate science methodology, Slater et al. (2021) outlined the possible behavior of time series data. Using theoretical plots, the authors show that data can a) have the same mean and variance over time, or b) have non-stationarity that can be broken into three major groups – trend, step change, or shifts in variance. Figure 1 further demonstrates the difference between stationary vs. non-stationary data in relation to a given variable of interest over time.
Figure 1. Plots showing the possible magnitude of a given variable across a time series: a) Stationary behavior, b) Non-stationary trend, step-change, and a shift in variance. [Taken from Slater et. al(2021)].
Traditional correlation statistics assumes stationarity, but it has been shown that ecological time series are often non-stationary at certain scales (Cazelles & Hales, 2006). In fact, ecological data rarely meets the requirements of a controlled experiment that traditional statistics require. This non-stationarity of ecological data means that while widely-used methods like generalized linear models and analyses of variances (ANOVAs) can be helpful to assess correlation, they are not always sufficient on their own to describe the complex natural phenomena ecologists seek to explain. Non-stationarity occurs frequently in ecological time series, so it is appropriate to consider analysis tools that will allow us to detect edges to further investigate the cause.
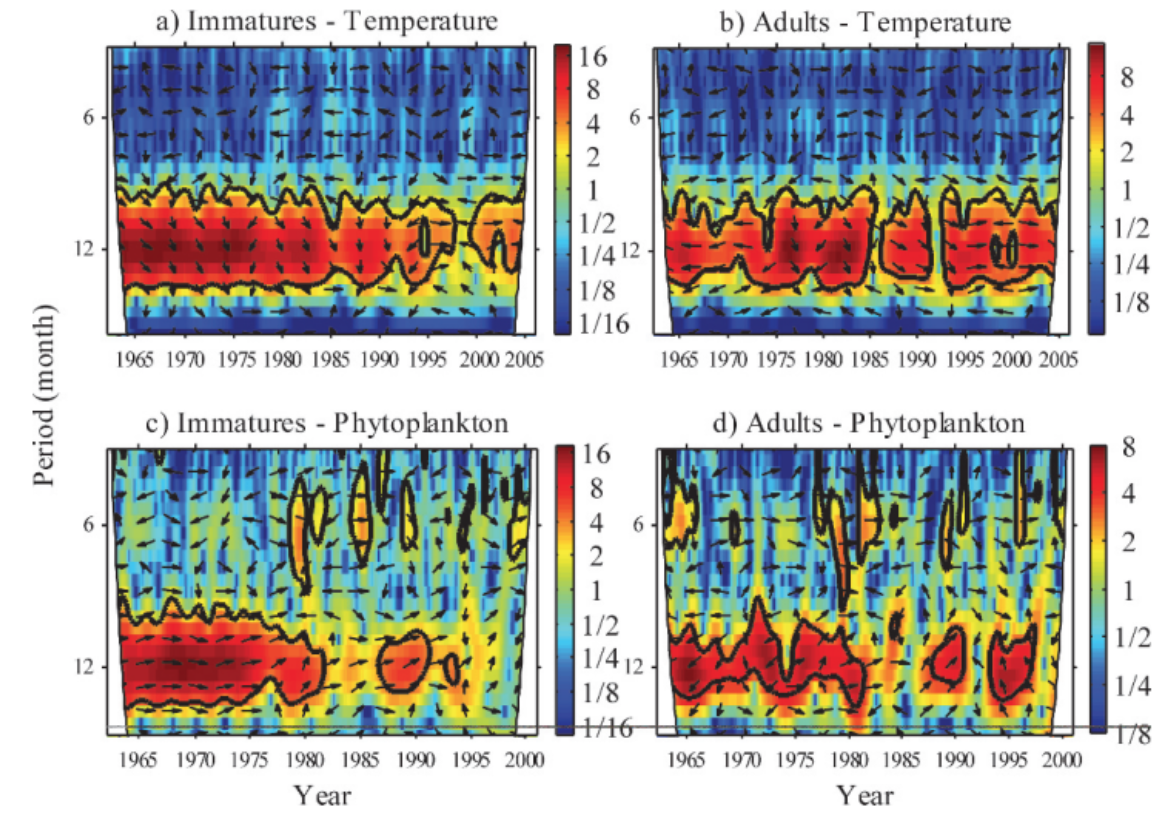
Wavelet analysis can also be conducted across a time series of multiple response variables to assess if these variables share high common power (correlation). When data is combined in this way it is called a cross-wavelet analysis. An interesting paper used cross-wavelet analysis to assess the seasonal response of zooplankton life history in relation to climate warming (Winder et. al 2009). Results from their cross-wavelet analysis showed that warming temperatures over the past two decades increased the voltinism (number of broods per year) of copepods. The authors show that where once annual recruitment followed a fairly stationary pattern, climate warming has contributed to a much more stochastic pattern of zooplankton abundance. From these results, the authors contribute to the hypothesis that climate change has had a temporal impact on zooplankton population dynamics, and recruitment has increasingly drifted out of phase from the original annual cycles.
Figure 2. Cross-wavelet spectrum for immature and adult Leptodiaptomus ashlandi for 1965 through either 2000 or 2005. Plots show a) immatures and temperature, b) adults and temperature, c) immatures and phytoplankton, and d) adults and phytoplankton. Arrows indicate phase between combined time series. 0 degrees is in-phase and 180 degrees is anti-phase. Black contour lines show “cone of influence” or the 95% significance level, every value within the cone is considered significant. Left axis shows the temporal period, and the color legend shows wavelet frequency power, with low frequencies in blue and high frequencies in red. Plots show strong covariation of high common power at the 12-month period until the 1980s. This pattern is especially evident in plot c) and d). [Taken from (Winder et. al 2009)].
While wavelet and cross-wavelet analyses should not be the only tool used to explore data, due to its limitations with significance testing, it is still worth implementing to gain a better understanding of how time series variables relate to each other over multiple spatial and/or temporal scales. It is often helpful to combine multiple methods of analysis to get a larger sense of patterns in the data, especially in spatio-temporal research.
When conducting research within the context of climate change, where the concentration of CO2 in ppm in the atmosphere is a non-stationary time series itself (Figure 3), it is important to consider how our datasets might be impacted by climate change and wavelet analysis can help identify the scales of change.
When considering our ecological time series of data in Port Orford, we want to evaluate how changing ocean conditions may be related to data trends. For example, has the annual mean or variance of zooplankton abundance changed over time, and where has that change occurred in time or space? These changes might have occurred at different scales and might be invisible at other scales. I am eager to see if wavelet analysis can detect these sorts of changes in the abundance of zooplankton across our time series of data, particularly during the seasons of intense heat waves or upwelling.
Did you enjoy this blog? Want to learn more about marine life, research and conservation? Subscribe to our blog and get a weekly email when we make a new post! Just add your name into the subscribe box on the left panel.
References
Buchan, S. J., Pérez-Santos, I., Narváez, D., Castro, L., Stafford, K. M., Baumgartner, M. F., … & Neira, S. (2021). Intraseasonal variation in southeast Pacific blue whale acoustic presence, zooplankton backscatter, and oceanographic variables on a feeding ground in Northern Chilean Patagonia. Progress in Oceanography, 199, 102709.
Cazelles, B., & Hales, S. (2006). Infectious diseases, climate influences, and nonstationarity. PLoS Medicine, 3(8), e328.
Salas, J. D., Anderson, M. L., Papalexiou, S. M., & Frances, F. (2020). PMP and climate variability and change: a review. Journal of Hydrologic Engineering, 25(12), 03120002.
Slater, L. J., Anderson, B., Buechel, M., Dadson, S., Han, S., Harrigan, S., … & Wilby, R. L. (2021). Nonstationary weather and water extremes: a review of methods for their detection, attribution, and management. Hydrology and Earth System Sciences, 25(7), 3897-3935.
Winder, M., Schindler, D. E., Essington, T. E., & Litt, A. H. (2009). Disrupted seasonal clockwork in the population dynamics of a freshwater copepod by climate warming. Limnology and Oceanography, 54(6part2), 2493-2505.
If you are an avid reader of our blog, you probably know quite a bit about gray whales, specifically the Pacific Coast Feeding Group (PCFG) of gray whales. Of the just over 50 GEMM Lab blogs written in 2021, 43% of them were about PCFG gray whales (or at least mentioned gray whales in some way). I guess this statistic is not too surprising when you consider that six of the 10 GEMM Lab members conduct gray whale-related research. You might think that we would have reached our annual limit of online gray whale content with that many blogs featuring these gentle giants, but you would in fact be wrong…
At the end of 2021, we launched a brand new website all about gray whales called IndividuWhale! It features stories of some of the Oregon coast’s most iconic gray whales, as well as information about how we study them, stressors they experience in our waters, and even a game to test your gray whale identification skills. IndividuWhale is a true labor of love that took over a year to create and that we are extremely proud to share with you today. Before I tell you more about the website, I want to take a moment to give a huge shout out to Erik Urdahl who was instrumental in getting this website off the ground and making it as interactive and beautiful as it is – hurrah Erik!
Equal‘s right side with visible boat propeller scars. Source: GEMM Lab.
Like us humans, gray whales have individual personalities and stories. They experience life-altering events, go through periods of stress, must provide for their offspring, and can behave differently to one another. Since Leigh & co. have been conducting in-depth research about PCFG gray whales in Oregon waters since 2016, we have been able to document several fascinating stories and events that these individuals have experienced. Take Equal, for example, a male whale that is at least 7 years old. The GEMM Lab observed Equal on consecutive days in June 2018, where on the first day he looked healthy and normal, but on the second day had fresh boat propeller-like scars on his back. Not only did we document these scars in photographs, but we were also able to collect a fecal sample from Equal the day we observed him with these scars. After analyzing his fecal sample for stress hormones, we discovered that Equal had very high stress levels compared to previous samples collected – unsurprising seeing as he had been hit by a boat! While this event was certainly sad for Equal (although don’t worry – we have seen him many times again in the years after this event looking healthy & normal once again), it was a very fortuitous occurrence for us since we were able to “validate” our stress hormone data relative to the value from Equal when he was clearly stressed out. Find out more about Equal as well as seven other gray whales here!
You might be wondering, how we knew that the whale with the boat propeller scar was Equal and how we recognize him again years after the incident. Gray whales have unique pigmentation patterns on their bodies and flukes that allow us to re-identify individuals between years and distinguish them from one another. Additionally, scars, such as those that Equal now carries on his back, can also be useful in telling whales apart. Therefore, we take photographs of every whale we see to match markings and identify whales. This process is called photo ID. Some individuals can have very distinctive markings, such as Roller Skate who has two big white dots on her right side, while others can look more inconspicuous, like Clouds. Therefore, conducting photo ID requires a lot of attention to detail and perseverance. To learn more about the different features we use to identify individuals, check out the “Studying Whales With Photographs” page. Do you think you have what it takes to tell individuals apart? Then try your luck at our photo ID game after!
Test your photo ID skills in our whale match game!
Unfortunately, these whales do not live in a pristine environment, as is evidenced by Equal’s story. Their main objective during the summer when in Oregon waters is to gain weight (energy stores) by consuming large amounts of prey, which is made more difficult by a number of stressors, including potential fishery entanglements, ocean noise, vessel traffic, and habitat changes. We describe these four stressors on the IndividuWhale website since we are trying to assess the impacts of them on gray whales through our research, however they are certainly not the only stressors that these whales experience. Little is known about the level at which these stressors may have a negative impact on the whales, and how whales react when they experience some of these stressors in concert. The answers to these questions are difficult to tease apart but the GRANITE project is aiming to do so through a framework called Population Consequences of Multiple Stressors (read more about it here). This approach requires a lot of data on a lot of individuals in a population and as you can see from the IndividuWhale website, we are slowly starting to get there! 2022 will certainly bring many more gray whale-themed blogs to this website, and you can share in our journey of learning about the lives of PCFG gray whales by exploring the IndividuWhale website (https://www.individuwhale.com).
Another year has come and gone, and the GEMM Lab has expanded in many facets! Every year it gets just a little bit harder to succinctly summarize all of the research, outreach, and successes that the GEMMs accomplish but it is an absolute honor and thrill to be a member of this lab. So, please enjoy the 6th edition of a GEMM Lab Year in the Life!
Our lab has almost doubled in size since I wrote the 2020 edition of this blog! This year we welcomed the arrival of two postdocs (Drs. Alejandro [Ale] Fernández Ajó and KC Bierlich) and two Master’s students (Allison Dawn and Miranda Mayhall). Ale and KC joined us as freshly minted Drs., as Ale defended his doctoral thesis from Northern Arizona University in April, while KC graduated in May from Duke University. Both of them immediately jumped into GRANITE fieldwork, scooping gray whale poop and flying drones (more below). Allison also dove headfirst into gray whale fieldwork as she co-led the TOPAZ and JASPER projects with me (Lisa) after defending her undergraduate thesis and graduating from the University of North Carolina with highest honors in the spring. Miranda, a U.S. Army Intelligence veteran, also joins us from the East Coast as she moved from Virginia to Oregon with her 10-year-old daughter, Mia, and two dogs, Angus and Mr. Gibbs. Unlike our other new arrivals, Miranda’s research does not relate to gray whales as she is part of the GEMM Lab’s newest research project…
There are exciting developments in the research project realm of the GEMM Lab every year. This year’s new project, HALO (Holistic Assessment of Living marine resources off Oregon), is particularly exciting as it is a joint project with the Cornell Lab, with GEMM Lab PI Leigh collaborating with Dr. Holger Klinck to better understand cetacean distributions off Oregon. HALO will involve monthly survey cruises aboard MMI’s R/V Pacific Storm along the Newport Hydrographic line (65 nm to 5 nm off Newport), where three Rockhopper hydrophones have been deployed and are passively monitoring cetacean acoustics. The HALO team, which includes GEMM students Miranda and PhD candidates Dawn Barlow and Rachel Kaplan, has already had two successful cruises this year! Check out the HALO website to stay tuned for updates throughout 2022. In addition to starting new research projects in our Oregon backyard, the GEMM Lab has also ventured further north, to the more frigid waters of Kodiak, Alaska. Postdocs KC and Ale went on a scouting mission to Kodiak Island to see whether the multidisciplinary methods we use in the GRANITE project to study PCFG gray whales in Oregon, can also be applied to other gray whales in other study areas. The reconnaissance trip was a huge success with KC and Ale making vital connections with potential collaborators and managing to collect some pilot data (drone flights, prey samples, and one fecal sample!). Both of these new ventures are funded by sales and renewals of the special Oregon gray whale license plate, which benefits MMI. We gratefully thank all the gray whale license plate holders, who made this research possible, and encourage any Oregonians that don’t have a whale on their tale yet, to do so in 2022!
These new research ventures certainly do not mean that we neglected our already established field research projects – in fact, most of them have flourished and thrived this year! Rachel and Dawn returned as marine mammal observers to the R/V Shimada for the May stint of the Northern California Current research cruise. They observed Dall’s porpoise, Northern right whale & Pacific white-sided dolphins, as well as killer, humpback, & fin whales. These sightings will add to the growing OPAL (Overlap Predictions About Large whales) dataset that both Rachel and postdoc Solène Derville are analyzing to better understand whale distribution patterns in Oregon waters. Speaking of OPAL, MMI Faculty Research Assistant Craig Hayslip and Leigh continued to take to the skies in U.S. Coast Guard helicopters to obtain monthly cetacean distribution data, which is also being used in the OPAL project to identify the co-occurrence between whales and fishing effort in Oregon to reduce entanglement risk. Both of our gray whale projects, GRANITE (Gray whale Response to Ambient Noise Informed by Technology & Ecology) and TOPAZ (Theodolite Overlooking Predators & Zooplankton)/JASPER (Journey for Aspiring Scientists Pursuing Ecological Research) had another year of successful field seasons. The GRANITE team, which includes Leigh, Todd Chandler, Ale, KC, PhD student Clara Bird, and myself, headed out in search of gray whales earlier than usual this year to document the potential effects of a National Science Foundation (NSF) funded seismic survey, which was conducted off the Oregon coast, on gray whales in the area. By the end of October, we had conducted 80 drone flights, collected 48 and 66 fecal and prey samples (respectively), and seen 36 individual whales during 201 sightings. Down south in Port Orford, the TOPAZ/JASPER project experienced a passing of the torch as I stepped down from the team lead position (which I held since 2018) and handed the project reins over to Allison. We co-led another fantastic field season this year. While whale sightings were much lower than in previous years (read some musings here), the project continued to be successful at making real impacts on young people’s lives as we once again engaged a local Pacific High School student (Damian Amerman-Smith) and two OSU undergraduates (Nadia Leal & Jasen White) in the field work. While our annual reach may be small in terms of numbers, the impact we have is huge, with many of the high school interns (including this year’s) deciding to go to college and/or to study biology directly as a result of our project.
TOPAZ/JASPER certainly is not the only project in our lab that engages students in ecological research. This year, we collectively oversaw and mentored 13 students. The OBSIDIAN (Observing Blue whale Spatial ecology to Investigate Distribution In Aotearoa New Zealand) project was assisted by three interns (Grace Hancock, Mateo Estrada Jorge, and Mattea Holt Colberg) overseen by Dawn and Leigh. Grace worked on maintaining the New Zealand blue whale photo-ID catalogue and won best student poster at our department’s annual student conference (RAFWE) for this work. Mattea, a 2020 TOPAZ/JASPER team member, switched study species and assisted Dawn in validating blue whale calls and songs. Mateo was a NSF Research Experience Undergraduate (REU) who conducted an analysis on blue whales and earthquakes. Clara also supervised a REU student with Leigh: Marc Donnelly, who created a habitat map for the GRANITE project. Rachel mentored Amanda Kent, an Undergraduate Research, Scholarship, & the Arts (URSA) Engage student, who helped her conduct a literature review about two Oregon krill species that are primary prey of whales. Over the summer, we had two student workers (Noah Goodwin-Rice & Julia Parker) join us in our efforts to better understand gray whale prey. Noah assisted us by sorting and identifying gray whale prey samples collected this summer and Julia wrapped up the microplastics analysis of gray whale prey and fecal samples. In the fall, both Clara and Allison supervised students (Kathryne Macallan & Jasen White, respectively) taking the Coastal & Estuarine Research Management class in our department who produced independent research projects during the term. Kathryne investigated the relationship between body length and blow intervals of gray whales during different behavioral states, while Jasen dove into the relationship between zooplankton abundance and environmental covariates.
The sharing of our research and expertise was not limited to mentoring students. Despite most conferences and seminars still occurring virtually this year due to the pandemic, the GEMM Lab presented numerous talks including at the State of the Coast (Rachel, Dawn, Leigh, & myself), International Biologging Symposium (Solène), HMSC Research Seminar (Ale & Solène, KC), and the Northwest Student Society of Marine Mammalogy chapter conference (Clara, Dawn, & myself), to name a few. Furthermore, Clara and I were guest lecturers once again for Dr. Renee Albertson’s marine mammal classes in our department, and Solène gained her first teaching experience by creating and leading a data visualization workshop (called “Pimp my figure!”) for RAFWE in May, which she reiterated at the University of New Caledonia in October.
Another huge accomplishment comes from the southern hemisphere as the hard work and time that Leigh and Dawn dedicated to OBSIDIAN and the results generated contributed to the denial of a seabed mining permit to extract iron sands in the South Taranaki Bight. This milestone has been years in the making, starting in 2013 when Leigh published her hypothesis that an unrecognized blue whale foraging ground existed in New Zealand. Since then, Leigh and Dawn have been building a tower of knowledge about these resident New Zealand blue whales block-by-block. They first confirmed Leigh’s hypothesis by presenting a bounty of evidence in support of this resident population, then assessed the skin condition of these whales, modeled the functional relationships between oceanography, krill and the distribution of blue whales, discovered temporal and spatial lags between wind, coastal upwelling, and blue whale occurrence, and most recently, developed dynamic models to forecast blue whale distribution three weeks into the future. We are extremely proud of the direct applications that the OBSIDIAN research outputs have had on the management and conservation of these New Zealand blue whales – hurrah to Leigh & Dawn!!
Other hurrahs this year include that Rachel passed her College of Earth, Ocean, & Atmospheric Sciences qualifying exam, now making her a PhD Candidate. Clara also reached a graduate milestone this year as she not only formed her PhD committee but also successfully defended her research review in the spring. Additionally, Clara became a certified drone pilot right before the start of the GRANITE field season and joined Todd and KC as pilots this summer. The lab and its members also received numerous grants and awards. There are too many to name for this blog, but we are very grateful for all of them! I do want to highlight two here: Dawn was awarded the Bob Moch memorial endowment award that recognizes service to the Hatfield Marine Science Center (HMSC) and broader Oregon coast community. I cannot think of anyone more deserving of this award than Dawn who truly does so much to serve and better the HMSC and Oregon coast communities! Clara was awarded a prestigious ARCS (Achievement Rewards for College Scientists) scholarship which provides awards to academically outstanding students to further their scientific knowledge.
We have once again been prolific writers, contributing 24 total peer-reviewed publications to 17 different scientific journals. If you are in the mood for some holiday reading, you will find the full list of publications at the end of this post.
And YOU, our awesome, supportive readers, have once again been supportive viewers, with a whopping 27,135 views of our blog this year!!! Thank you for joining us on our 2021 journey! We hope you have enjoyed the tales that we have told and the knowledge we have (hopefully) conveyed. We wish you all restful, happy, and most importantly, healthy holidays and hope you will join us again in 2022!
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get weekly updates and more! Just add your name into the subscribe box on the left panel.
Publications
Andréfouët, S., Derville, S., Buttin, J., Dirberg, G., Wabnitz, C.C.C., Garrigue, C., & Payri, C. E. 2021. Nation-wide hierarchical and spatially-explicit framework to characterize seagrass meadows in New Caledonia, and its potential application to the Indo-Pacific. Marine Pollution Bulletin 173:113036. https://doi.org/10.1016/j.marpolbul.2021.113036.
Barlow, D.R., & Torres, L.G. 2021. Planning ahead: Dynamic models forecast blue whale distribution with applications for spatial management. Journal of Applied Ecology. (Link)
Barlow, D.R., Klinck, H., Ponirakis, D., Garvey, C., & Torres, L.G. (2021). Temporal and spatial lags between wind, coastal upwelling, and blue whale occurrence. Scientific Reports 11(1):1-10. (Link)
Beal, M., … Torres, L.G., et al. 2021. Global political responsibility for the conservation of albatrosses and large petrels. Science Advances 7(10):eabd7225.
Bierlich, K.C., Schick, R.S., Hewitt, J., Dale, J., Goldbogen, J.A., Friedlaender, A.S., & Johnston D.J. 2021. A Bayesian approach for predicting photogrammetric uncertainty in morphometric measurements derived from UAS. Marine Ecology Progress Series. DOI: https://doi.org/10.3354/meps13814
Bierlich, K.C., Hewitt, J., Bird, C.N., Schick R.S., Friedlaender, A.S., Torres, L.G., Dale, J., Goldbogen, J.A., Read, A., Calambokidis J., & Johnston, D.W. 2021. Comparing uncertainty associated with 1-, 2-, and 3D aerial photogrammetry-based body condition measurements of baleen whales. Frontiers in Marine Science 8:749943. doi: 10.3389/fmars.2021.749943
Bonneville, C.D., Derville, S., Luksenburg, J.A., Oremus, M., Garrigue, C. 2021. Social structure, habitat use and injuries of Indo-Pacific Bottlenose Dolphins (Tursiops aduncus) reveal isolated, coastal, and threatened communities in the South Pacific. Frontiers in Marine Science 8:1–14. https://doi.org/10.3389/fmars.2021.606975.
D’Agostino, V.C., Fernandez, A.A.A., Degrati M., Krock, B., Hunt, K.E., Uhart, M.M., & Buck, C.L. 2021. Potential endocrine correlation with exposure to domoic acid in Southern Right Whale (Eubalaena australis) at the Península Valdés breeding ground. Oecologia 1-14.
Dillon, D., Fernandez, A.A.A., Hunt, K.E., & Buck, C.L. 2021. Investigation of keratinase digestion to improve steroid hormone extraction from diverse keratinous tissues. General and Comparative Endocrinology 309:113795.
Fernandez, A.A.A., Hunt, K.H., Sironi, M., Uhart, M., Rowntree, V., Giese, A.C., Maron, C.F., DiMartino, M., Dillon, D., & Buck, C.L. 2021. Retrospective analysis of the lifetime endocrine response of southern right whales calves to gull wounding and harassment: a baleen hormone approach. Integrative and Comparative Biology 61.
Fernandez, A.A.A., Hunt, K.E., Dillon, D., Uhart, M., Sironi, M., Rowntree, V., & Buck, C.L. 2021. Optimizing hormone extraction protocols for whale baleen: tackling questions of solvent: sample ratio and variation. General and Comparative Endocrinology 113828.
Garrigue, C., & Derville, S. 2021. Behavioral responses of humpback whales to biopsy sampling on a breeding ground : the influence of age-class , reproductive status , social context , and repeated sampling. Marine Mammal Science 1–16. https://doi.org/10.1111/mms.12848.
Gough, W.T., Smith, H.J., Savoca, M.S., Czapanskiy M.F., Fish, F.E., Potvin, J., Bierlich, K.C., Cade, D.E., Di Clemente, J., Kennedy, J., Segre, P., Stanworth, A., Weir, C., & Goldbogen, J.A. 2021. Scaling of oscillatory kinematics and Froude efficiency in baleen whales. Journal of Experimental Biology224(13):jeb237586. DOI: https://doi.org/10.1242/jeb.237586
Hildebrand, L., Bernard, K.S., & Torres, L.G. 2021. Go gray whales count calories? Comparing energetic values of gray whale prey across two different feeding grounds in the eastern North Pacific. Frontiers in Marine Science, https://doi.org/10.3389/fmars.2021.683634
Jones, D.C., Ceia, F.R., Murphy, E., Delord, K., Furness, R.W., Verdy, A., Mazloff, M., Phillips, R.A., Sagar, P.M., Sallée, J-B., Schreiber, B., Thompson, D.R., Torres, L.G., Underwood, P.J., Weimerskirch, H., & Xavier J.C. 2021. Untangling local and remote influences in two major petrel habitats in the oligotrophic Southern Ocean. Global Change Biology 27(22):5773-5785.
Kone, D.V., Tinker, M.T., & Torres, L.G. 2021. Informing sea otter reintroduction through habitat and human interaction assessment. Endangered Species Research 55:159-176.
Lemos, L.S., Olsen, A., Smith, A., Burnett, J.D., Chandler, T.E., Larson, S., Hunt, K.E., & Torres, L.G. 2021. Stressed and slim or relaxed and chubby? A simultaneous assessment of gray whale body condition and hormone variability. Marine Mammal Science.
Lemos, L.S., Haxel, J.H., Olsen, A., Burnett, J.D., Smith, A., Chandler, T.E., Nieukirk, S.L., Larson, S.E., Hunt, K.E., & Torres, L.G. 2021. Sounds of stress: assessment of relationships between ambient noise, vessel traffic, and gray whale stress hormone. Scientific Reports. DOI:10.21203/rs.3.rs-923450/v1
Maron, C.F., Lábaque, M.C., Beltramino L., DiMartino, M., Alzugaray, L., Ricciardi, M., Fernandez, A.A.A., Adler, F.R., Seger, J., Sironi, M., Rowntree, V.J., & Uhart, M.M. 2021. Patterns of blubber fat deposition and evaluation of body condition in growing southern right whale calves (Eubalaena australis). Marine Mammal Science. DOI: 10/1111/mms.12818.
Orben, R.A., Adams, J., Hester, M., Shaffer, S.A., Suryan, R.M., Deguchi, T., Ozaki, K., Sato, F., Young, L.C., Clatterbuck, C., Conners, M.G., Kroodsma, D.A., & Torres, L.G. 2021. Across borders: External factors and prior behavior influence North Pacific albatross associations with vessel traffic. Journal of Applied Ecology.
Savoca, M.S. Czapanskiy, M.F., Kahane-Rapport, S.R., Gough, W.T., Falhbusch, J.A., Bierlich, K.C., Segre, P.S., Di Clemente, J., Penry G.S., Wiley, D.N., Calambokids, J., Nowacek, D.P., Johnston, D.W., Pyenson, N.D., Friedlaender, A.S., Hazen, E.L., & Goldbogen, J.A. 2021. Baleen whale prey consumption based on high-resolution foraging measurements. Nature 599:85–90. https://doi.org/10.1038/s41586-021-03991-5
Stephenson, F., Hewitt, J.E., Torres, L.G., Mouton, T.L., Brough, T., Goetz, K.T., Lundquist, C.J., MacDiarmid, A.B., Ellis, J., & Constantine, R. 2021. Cetacean conservation planning in a global diversity hotspot: dealing with uncertainty and data deficiencies. Ecosphere 12(7):e03633.
Thompson, D.R., Goetz, K.T., Sagar, P.M., Torres, L.G., Kroeger, C.E., Sztukowski, LA., Orben, R.A., Hoskins, A.J., & Phillips, R.A. 2021. The year-round distribution and habitat preferences of Campbell albatross (Thalassarche impavida). Aquatic Conservation: Marine and Freshwater Ecosystems 31(10):2967-2978.