I successfully defended my PhD 55 days ago and I have been grappling with a lot of complex emotions since then. I feel pride in having accomplished such a milestone, sadness to have left such an incredible lab and workplace, excitement to see what the future holds, and, admittedly, also relief to have a bit of time off. I have also been reflecting about all the things I have learned during my seven-year journey of being a graduate student. Naturally there is all the ecological, quantitative and communication knowledge and skills I learned that have shaped me into the scientist I am today. However, there are so many lessons that I have learned that are less tangible but equally as important in building a successful career in academia, I think.
Feeling stupid is kind of the point
Graduate school is riddled with unknowns. As we embark on these journeys there is much to be learned and it can be exhausting not knowing a lot of the time. During many stints of my graduate school journey, I felt stupid, with varying magnitudes of stupidity. Whether I felt stupid because I had to Google “how to save one object in R environment” for the thousandth time or because I simply could not grasp why a Bayesian posterior distribution was so much better than a frequentist p-value result. If you’re struggling with feeling stupid in academia, know that it is actually an important part of the process – sitting in the discomfort of not knowing something is what drives us to get answers. Of course, in graduate school, it isn’t that we just don’t know the answer to a scientific question (for example, in my case, how do different individual gray whales use space during a foraging season?), but we also don’t know how to get the answers to that question right away. Before I could even begin to tackle that question for one of my PhD chapters, I had to learn about how to quantify survey effort from survey conditions, the principles of Bayesian statistics, theoretical spatial ecology, coding in Python, capture-recapture models, and so much more. Each of those pursuits involved weeks, if not months, of feeling stupid. But feeling stupid throughout the pursuits was kind of the point. There’s an incredibly excellent (and short) article on this topic by Martin Schwartz that I cannot recommend highly enough.
Asking questions isn’t a weakness, it’s a strength
Somewhat related to the importance of feeling stupid, is realizing that it’s ok, neigh even necessary, to ask for help. Learning to be an independent problem-solver is of course a huge part of being a graduate student. But it’s also important to learn that at some points, continuing to bang your head against the keyboard because you can’t figure something out, is not the most productive use of your time. I have been so lucky to have been surrounded by so many knowledgeable, generous and kind people throughout my graduate career, ranging from fellow graduate students to lab mates, to project colleagues, and my committee members and advisor. I have benefitted so much from countless Zoom meetings or whiteboarding sessions where people have shared their wisdom with me in helping me solve a problem. I know this quote is probably a cliché but I think it holds so much truth: “Ask a question and be a fool for five minutes, don’t ask a question and be a fool for life.”. More often than not, a question that you ask is one that someone else probably also had in mind but was too shy or embarrassed to ask. So, be the risk taker and ask that question!
Learning takes time
The months leading up to a defense are usually a massive crunch. There are analyses that are still getting figured out, chapters that need to be written, citations that need to be integrated, and a big beefy Word document that needs to be formatted according to very specific thesis guidelines. By the end of a degree, a graduate student has learned so much and honed so many skills that tasks are happening much more efficiently than they did the first time around. A few weeks before my defense, I had a conversation with Marine Mammal Institute research associate Selene Fregosi where I lamented about not having achieved enough in the first three years of my PhD seeing as I was able to complete two data analyses and chapters in the final six months of my PhD. It left me wondering, what did I spend all those years doing at the start of my PhD?! Selene chuckled in response and she said, “Lisa, you were learning!”. Selene helped me realize that it was those years spent in the slow learning process that allowed me to be fast yet effective in the final months before my defense, enabling the completion of two chapters in a much shorter amount of time than my first chapters took. That flip in perspective was a much kinder way of looking at the years gone by. Dovetailed with this lesson is that everything takes way longer than you think it will, especially if it involves learning, so budget for learning time!
Adaptability is key
I consider myself a Type A personality. I like being organized, I like having a plan, and I like knowing what is around every corner. Unfortunately, a graduate student trajectory (and life!) rarely has a defined, clear, unchanging path. I recently looked back at a document titled ‘PhD Chapter Ideas’ that I made in June 2020 while I was in the final six months of my Master’s degree and starting to brainstorm with Leigh about what my PhD dissertation could entail. Of the six ideas listed in that document, only two of them sort of align with the chapters that ended up in my final dissertation five years later. My dissertation morphed quite a bit over the years and this change is inevitable. So many things can go wrong during scientific endeavors: equipment can malfunction and fail, data may not be available in time or impossible to acquire, proposed collaborations may not take place, weather, funding hiccups, computational limitations, to name a few. While it is important to be persistent and to find solutions to problems that arise, sometimes the solution to a problem may be pivoting and embracing change.
Thinking ahead
As I begin the next, somewhat daunting, journey of entering the job market, I am reminded that graduate school has shaped me in ways I did not always recognize at the time. I take comfort in knowing that graduate school has prepared me in ways that extend far beyond technical training. Alongside tools and knowledge, I leave with resilience, perspective, and a collection of hard-earned lessons that will continue to guide me as I navigate what comes next.
Baleen whales must navigate a seemingly featureless world to locate the resources they need to survive. The task of finding prey to feed on in the vast seascapes relies on the use of several sensory modalities that operate at different scales (Torres 2017; Figure 1). For example, baleen whale vision is believed to be rather limited, with the ability to see objects about 10-100 meters away. Yet, baleen whale somatosensory perception of oceanographic stimuli is thought to be on the order of 100-1000s kilometers. This diversity in sensory ability has led scientists to believe that whales, in fact all animals, perceive cues and make decisions at several scales. As ecologists, we endeavor to understand why and when animals are found (or not found) in certain locations as this knowledge allows us to better manage and conserve animal populations. With this information we can aim to minimize potential anthropogenic disturbance and protect important resource areas, such as foraging or nursing grounds. In order to accomplish this goal, we ourselves must conduct studies and test hypotheses at several scales (Levin 1992; Hobbs 2003). As someone who tackles spatial foraging ecology questions, I am particularly interested in understanding whale behavior and movement in the context of feeding. Since accurately measuring predator and prey distribution at the same scales can be challenging, we often resort to environmental variables to serve as proxies for prey, whereby we look for correlations between environmental variables and whales to understand and predict the distribution of our population.
Figure 1. Schematic of hypothetical interchange of sensory modalities used by baleen whales to locate prey at variable scales. X-axis represents log distance to prey from micro (left) to macro (right). Y-axis represents the relative use of each sensory modality between 0 (no contribution) to 10 (highest contribution). Each line and color represent a different sensory modality. Figure taken and caption adapted from Torres 2017.
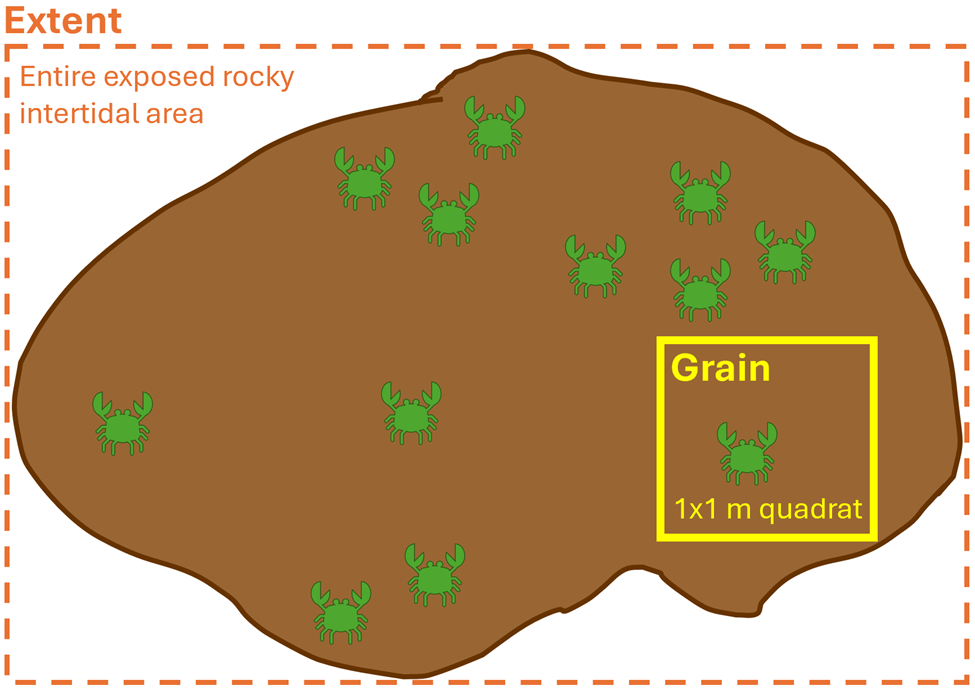
What do I mean when I use the word ‘scale’? The term scale is typically explained by two components: grain and extent (Wiens 1989). The grain is the finest resolution measured; in other words, how detailed we are measuring. The extent is the overall coverage of what we are measuring. These components can be applied to both spatial scale and temporal scale. For example, spatially, if we were using a 1×1 meter sampling quadrat to count the number of crabs on a rocky shore, then our grain would be the 1 m2 quadrat and the extent would be the entire exposed rocky intertidal area that we are surveying (Figure 2). Temporally, if we placed a temperature logger at the mouth of Yaquina Bay that took a temperature recording every minute for two years, then our grain would be one minute and the extent would be two years. So, when designing a study, it is imperative for us to decide on the spatiotemporal scales of the ecological questions we are asking and the hypotheses we are testing, as it will inform what data we need to collect. When making this decision, it is important to think about the scale at which the ecological process happens, as opposed to the scale at which we can observe the process (Levin 1992). In other words, we need to think from the perspective of our study species, as opposed to from our own human perspective. Making informed and ecologically reasonable decisions regarding the choice of scale relies on having prior knowledge of an animal’s biology, such as knowing that baleen whales might see a prey patch that is 50 meters away, but it may also somatosensorily perceive an oceanic front where zooplankton prey aggregate from 500 kilometers away.
Figure 2. Schematic of spatial scale where the extent (depicted by dashed orange box) is the entire exposed rocky intertidal area being surveyed and the grain (solid yellow box) is the 1×1 m quadrat being used to count crabs.
There is a wealth of studies that have explored space use patterns of wildlife relative to environmental variables to better understand foraging behavior. I want to share a couple from the marine mammal realm with you that I find particularly fascinating. In their 2018 study, González García and colleagues used opportunistic sightings of blue whales around the Azorean islands of Portugal and modeled their distribution patterns relative to physiographic and oceanographic variables summarized at different spatial (fine [1-10 km] and meso [10-100 km]) and temporal (daily, weekly, monthly) scales. The two variables that were most correlated with blue whale occurrence was distance from the coast and eddy kinetic energy (a measure of mesoscale variability of ocean dynamics). Both of these variables were interestingly found to be scale invariant, meaning that no matter which spatial and temporal scale was investigated, the relationship between blue whales and these two variables stayed the same; blue whale occurrence increased with increasing distance from the coast and was maximal at an eddy kinetic energy value of 0.007 cm2/s2 (Figure 3).
Figure 3. Functional response curves between presence of Azorean blue whales and distance to the coast (panel 1 on left) and eddy kinetic energy (panel 2 on right). The top row of each panel represents the low spatial scale and the bottom row represents the high spatial scale. Each column represents a different temporal scale (from left to right: daily, weekly, monthly). Note that the general shape of the relationship remains similar across all spatiotemporal scales and that the peak of the curves tend to occur at the same values for distance to coast and eddy kinetic energy across all scales. Figures taken from González García et al. 2018.
However, not all studies find scale invariant relationships. For example, Cotté and co-authors (2009) found that habitat use of Mediterranean fin whales was very much scale dependent. At a large scale (700-1,000 km and annual), fin whales were more densely aggregated during the summer in the Western Mediterranean where there was consistently colder water than in the winter. However, at a meso scale (20-100 km and weekly-monthly), fin whale densities were highest in areas where there were steep changes in temperature, as opposed to consistently cold temperatures. The authors explain that these differences in fin whale density and temperature at different scales are likely due to whale movement being driven by annually persistent prey abundance at the large scale, but at the meso scale, where prey aggregations are less predictable, the fin whales’ distribution becomes more driven by areas of physical ocean mixing.
As I investigate the environmental drivers of individual gray whale space use using our 8-year GRANITE (Gray whale Response to Ambient Noise Informed by Technology and Ecology) dataset, these studies (and many more) are at the top of my mind to interpret the patterns we are detecting. Our goal is to quantify and describe what environmental conditions (1) lead to a higher probability of a gray whale being seen in our central Oregon coast study area (~70 km) at a daily scale, and (2) influence space use patterns (activity range, residency, activity center) of different individual whales at annual scales. Our results show both consistency and variation in the environmental drivers of gray whales across these scales, leading me to deeply consider how gray whales make decisions at different points in their lives, based on information gained through various senses, to maximize their chances of capturing food. Previous work from the GEMM Lab on the relationships between gray whales and prey, at both fine (read more here) and large (read more here) scales have guided my work by providing specific hypotheses regarding environmental variables and lag times for me to test. Investigating the environmental drivers of animal space use and behavior is exciting work as it reveals that no single environmental variable determines animal distribution, but rather that multiple processes are happening concomitantly that animals respond to at different scales continually. It is only by studying animal space use patterns across spatiotemporal scales that we can begin to understand their complex decision-making patterns.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog below and get a monthly message when we post a new blog. Just add your name and email into the subscribe box below.
References
Cotté, C., Guinet, C., Taupier-Letage, I., Mate, B., & Petiau, E. (2009). Scale-dependent habitat use by a large free-ranging predator, the Mediterranean fin whale. Deep Sea Research Part I: Oceanographic Research Papers, 56(5), 801-811.
González García, L., Pierce, G. J., Autret, E., & Torres-Palenzuela, J. M. (2018). Multi-scale habitat preference analyses for Azorean blue whales. PLoS One, 13(9), e0201786.
Hobbs, N. T. (2003). Challenges and opportunities in integrating ecological knowledge across scales. Forest Ecology and Management, 181(1-2), 223-238.
Levin, S. A. (1992). The problem of pattern and scale in ecology: the Robert H. MacArthur award lecture. Ecology, 73(6), 1943-1967.
Torres, L. G. (2017). A sense of scale: Foraging cetaceans’ use of scale‐dependent multimodal sensory systems. Marine Mammal Science, 33(4), 1170-1193.
Wiens, J. A. (1989). Spatial scaling in ecology. Functional ecology, 3(4), 385-397.
By Hali Peterson, rising freshman, Western Oregon University
Hello, my name is Hali Peterson and I am a rising freshman in college. Last summer (2023) I was given the opportunity to be a paid high school intern for the OSU Marine Mammal Institute’s very own GEMM Lab (Geospatial Ecology of Marine Megafauna Laboratory) based at the Hatfield Marine Science Center in Newport, Oregon. My time working in the GEMM Lab has been supported by the Oregon Coast STEM Hub. I started my internship in June 2023 and I was one of the two GEMM Lab summer interns. However, my internship did not end when summer did, as I continued to work throughout the school year and even into this summer.
Figure 1: Leaving work late and accompanied with a beautiful view of the Newport bridge over Yaquina Bay.
June 29, 2023 to September 20, 2024 (1 year, 2 months, and 21 days if anyone is curious) – what did I do and what did I learn during this time…
Initially, I was tasked with helping the GRANITE project (Gray whale Response to Ambient Noise Informed by Technology and Ecology) by processing drone footage of Pacific Coast Feeding Group (PCFG) gray whales and identifying their zooplankton prey. I started off my internship under the mentorship of KC Bierlich and Lisa Hildebrand and I dove into looking at zooplankton underneath a microscope and watching whales in drone footage, both gathered by the GEMM Lab field team.
KC taught me how to process drone footage, measure whales and calibration boards, test an artificial intelligence model, as well as write a protocol of the drone processing methods that I had worked on. These tasks were a big responsibility as the measurements need to be accurate and precise so that they can be used to effectively assess the body condition of gray whales, which provides crucial insights into population health.
Figure 2: My favorite drone video of moms and calves meeting up for a playdate!
Under Lisa’s mentorship I learned how to identify and process zooplankton prey samples, process underwater GoPro videos, as well as identify and analyze kelp patches from satellite images. Within these tasks, I honed my expertise in zooplankton and habitat analysis and the results of my work will contribute to a deeper understanding of gray whale feeding habits along the Oregon coast.
Figure 3: My favorite zooplankton to see, a juvenile crab larva.
As my main mentors, KC and Lisa taught me so much about the world of science and research. All of these detail-oriented and multi-layered tasks helped me improve some of the skills I already had before I started the internship as well as gift me with skills I didn’t previously possess. For example, I learned how to collaborate and work with a team, pay attention to detail, double and even triple check everything for quality work, problem solve, and learn to ask questions.
However, as my time in the GEMM Lab extended beyond the summer of 2023, so did my tasks. Later on I received another mentor, Clara Bird. Under Clara I learned how to identify whales from drone footage recorded in Baja, Mexico (an area that is specifically known as the breeding lagoons where the gray whales go in the winter), as well as use the Newport, Oregon drone footage and CATS (Customized Animal Tracking Solution) tag data to measure inhalation duration and bubble blast occurrences. These experiences furthered my knowledge and yet again I learned something new, a common theme throughout my time in the GEMM Lab.
Just a few months ago, the GEMM Lab hired Laura Flores Hernandez as a new high school student summer intern, and under the guidance of both Lisa Hildebrand and Leigh Torres, I was given the opportunity to develop my own mentoring skills. I used the skills I had obtained over the past year to teach someone else how to do the tasks I once was new to. I taught Laura how to identify zooplankton, process drone footage, and measure calibration boards. Stepping into that mentor role helped me reflect on my own learning and experiences. I had to go back and figure out how I did things, where I struggled, and how I overcame those struggles. Not an easy task but one I was glad to be presented with.
Figure 4: Matthew Vaughan (chief scientist on the trip) and me (right) looking at a box core sample.
During my time here I was also invited to join a STEM (Science, Technology, Engineering, Mathematics) cruise led by Oregon Sea Grant with fellow high school students. On this science cruise I got to help look at box core samples (a tool used to collect large amounts of sediment off of the ocean floor). Equipped with my previous knowledge on zooplankton identification, I was able to help the chief scientist on the trip to explain to other high school students what we were seeing in the samples. This trip helped me grow my teamwork and identification skills, as well as experience what it is like to collect data while on a moving ship.
Figure 5: Sea Kayaking through the fjord with the Girls on Icy Fjords team of 2024.
Another amazing opportunity I was selected for was to join the 2024 Girls on Icy Fjords team. This program, in association with OSU, was designed to empower young women in STEM in the backcountry of Alaska. With a team of 3 amazing instructors and 8 girls (all from different parts of the United States of America) we camped in the backcountry for 8 days, learning about glaciers and fjords, surviving in the backcountry, sea kayaking, and working as a team. I would highly recommend any young woman interested in science, art, or just an amazing experience to check out Inspiring Girls Expeditions.
Bonus Image: This is Jeff the Moyebi Shrimp and I love him.
All in all this will be a job that I will not soon forget; interning in the GEMM Lab has been both a learning opportunity as well as a challenge. My internship wasn’t without its challenges, from a computer that seemed determined to shut down whenever I made progress, to endless hours spent staring at a green screen, waiting to count a fish that might eventually swim by. Though the job had its ups and downs, I am so glad I was given this opportunity and was kept on in the lab for as long as I was. In just a few weeks, I will start my Bachelors of Aquarium Science at Western Oregon University and I’m both excited and nervous. I know that without a doubt the skills I learned during this internship will come in handy as I continue my education and pursue a career in the future.
Thank you to all my mentors, anyone who answered one of the many questions I had, and to the friends I made along the way!
In case you aren’t already aware, I want to remind you of a website called IndividuWhale we created about Pacific Coast Feeding Group (PCFG) gray whales we study as part of our GRANITE project. IndividuWhale features stories of some of the Oregon coast’s most iconic gray whales, as well as information about how we study them, stressors they experience in our waters, and even a game to test your gray whale identification skills. We also provide details about where to best spot gray whales along our coast and the different behaviors you might see gray whales displaying at different times of the year. Since launching the website in late 2021, we have made small tweaks and updates along the way, but now, after about 2.5 years, the time has come for a major content update as we are introducing you to three new individuals and their stories! Head over to IndividuWhale.com to check out the updates or continue reading for a preview of the content…
Lunita
Even though “Lunita” is only two years old (as of 2024), they (sex currently unknown!) have quickly become a star of our dataset and hearts. We documented Lunita as a calf with their mother “Luna” (hence the name Lunita, which means little Luna/moon) in 2022. We observed the mom-calf pair in our study area for almost two weeks during which it seemed like Lunita was a very attentive calf, always staying close to Luna and appearing to benthic feed alongside their mom. As is often the case when we document mom–calf pairs, we wonder whether we will see the calf again and how it will fair in an environment increasingly impacted by human activities. Much to our delight, we were reunited with Lunita later in the same summer when we saw them feeding independently, indicating that they had successfully weaned. We were even more delighted when we were reunited with Lunita again many times during the summer of 2023 as Lunita spent almost the entire feeding season along the central Oregon coast. This is yet another example, much like “Cheetah” and “Pacman,” of successful internal recruitment of calves born to PCFG females into the PCFG sub-population.
Lunita’s high site fidelity to our study area in 2023 meant that she was an excellent candidate for the suction-cup tagging we have been conducting in the last few years. During suction-cup tagging, we attach a device (or tag) via suction cups to a whale’s back. The tag contains a number of different sensors, including an accelerometer (to measure speed), a gyroscope (to measure direction), and a magnetometer (to measure magnetic field), as well as a high-definition video camera and hydrophone (or underwater microphone). These tags typically stay on for a maximum of 24 hours before they pop off the whale leaving no harm to the whale. Upon retrieval, we can recreate the whale’s dive path and see the environment and conditions that the whale experienced over several hours. We sometimes refer to tagging as giving the gray whales some temporary jewelry because the tags are a very flashy, bright orange color. From the video from Lunita’s tag shows how they soared through kelp forests feeding on mysids for many, many hours. Check out their profile here: https://www.individuwhale.com/whales/lunita/
Burned
There are many ways to assess the health of a whale. In our lab, we calculate body condition from drone images to determine how fat or skinny a whale is, examine different hormones from their poop, and assess growth rates via length measurements from drone images. Another health assessment metric that we explore in the lab is the skin and scarring on the individuals that we see in our central Oregon study area. By conducting a skin and scarring analysis, we can identify scarring patterns and lesions that may indicate interactions with human activities and track the progression of skin diseases that will help us understand the prevalence and impacts of pathogens on whales. One skin condition that we are particularly interested in tracking appears as a thick white or gray layer that can mask a gray whale’s natural pigmentation. An example of a whale that has experienced this skin condition is “Burned.”
Burned is a female who is at least 9 years old (as of 2024), as she was first documented in the PCFG range in 2015. We saw Burned for the first time in 2016. At the time, we noticed small, isolated, gray patches of the skin condition on both sides of Burned’s body. Throughout the years as we have continued to resight Burned, we noticed the skin condition spreading progressively across her body. We saw the skin condition at its maximum extent in 2022 when, at first glance, Burned was hardly recognizable. Luckily, we can identify gray whales using more than just their pigmentation patterns (learn more on our whale identification page). Interestingly, when we saw Burned in June 2024, it appeared that the skin condition completely disappeared! Burned is just one example of whales with this skin condition, leaving us with many questions about its origin and impact on the whales: What causes the skin condition (viral, fungal, bacterial?); How it is transmitted (via air or contact?); Is it harmful to the whale (weakened immune system?). Our research is aimed at addressing these questions to make this skin condition a little less mysterious. Check out her profile here: https://www.individuwhale.com/whales/burned/
Heart
“Heart,” who is also known as “Ginger,” is a very well known and popular whale in the Depoe Bay region. Heart is a female who is particularly famous for being a “tall fluker,” meaning that when she dives, she arches her tail fluke high in the air before it glides elegantly into the water. Heart was first documented as a calf in 2010, which means that she is 14 years old (as of 2024). At 14 years of age, we would expect for Heart to have had at least one, if not more, calves by now, as it is believed that gray whales reach sexual maturity at age 8 or 9. However, Heart has never been documented with a calf. Why?
While we cannot know for sure, we have a theory that it might be linked to her body length. Recent work in our lab has explored how growth of PCFG whales has changed over time. Using measurements of whales from our drone data, we investigated how the asymptotic length (i.e. the final length reached once an individual stops growing) for the PCFG whales has changed since the 1980s. Shockingly, we found that starting in the year 2000 the asymptotic length of PCFG whales has declined at an average rate of 0.05–0.12 meters per year. Over time, this means that a whale born in 2020 is expected to reach an adult body length that is 13% shorter than a gray whale born prior to 2000. In Heart’s case specifically, when we last measured her length at 13 years old, she was 10.65 meters long. If she had been born prior to 2000, then she would be 12.04 meters long by now at the age of 13. That’s a whole 1.5 meters (or almost 5 feet) shorter!
You might be wondering how Heart’s length links back to her ability to have a calf. It takes a lot of energy to be pregnant and support the fetus, so by being smaller, Heart may not be able to store and allocate enough energy towards reproduction. Many of the whales we commonly see are shorter than expected based on their age (including “Zorro”), so we are monitoring the number and frequency of calves in the PCFG to see how this decline in length may impact the population. Check our her profile here: https://www.individuwhale.com/whales/heart/
Be sure to head over to IndividuWhale.com to explore all of the whale profiles and lots of other information that we have provided there about PCFG gray whales and how we study them here in Oregon waters!
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly message when we post a new blog. Just add your name and email into the subscribe box below.
As many of our avid readers already know, the Pacific Coast Feeding Group (PCFG) of gray whales employs a wide range of foraging tactics to feed on a number of different prey items in various benthic substrate types (Torres et al. 2018). One example foraging tactic is when PCFG whales, particularly when they are in the Oregon portion of their feeding range, forage on mysid shrimp in and near kelp beds on rocky reefs. We have countless drone video clips of whales weaving their large bodies through kelp and many photographs of whales coming to the surface to breath completely covered in kelp, looking more like a sea monster than a whale (Figure 1). So, when former intern Dylan Gregory made an astute observation during the 2018 TOPAZ/JASPER field season in Port Orford about a GoPro video the field team collected that showed many urchins voraciously feeding on an unhealthy-looking kelp stalk (Figure 2a), it made us wonder if and how changes to kelp forests may impact gray whales.
Fig 1. Gray whale surfacing in a large kelp patch. Photograph captured under NOAA/NMFS research permit #16111. Source: GEMM Lab.
Kelp forests are widely used as a marine example of trophic cascades. Trophic cascades are trigged by the addition/removal of a top predator to/from a system, which causes changes further down the trophic chain. Sea urchins are common inhabitants of kelp forests and in a balanced, healthy system, urchin populations are regulated by predators as they behave cryptically by hiding in crevices in the reef and individual urchins feed passively on drift kelp that breaks off from larger plants. When we think about who controls urchins in kelp forests, we probably think of sea otters first. However, sea otters have been absent from Oregon waters for over a century (Kone et al. 2021), so who controls urchins here? The answer is the sunflower sea star (Figure 2b). Sunflower sea stars are large predators with a maximum arm span of up to 1 m! Unfortunately, a disease epidemic that started in 2013 known as sea star wasting disease caused 80-100% population decline of sunflower sea stars along the coastline between Mexico and Alaska (Harvell et al. 2019). Shortly thereafter, a record-breaking marine heatwave caused warm, nutrient-poor water conditions to persist in the northeast Pacific Ocean from 2014 to 2016 (Jacox et al. 2018). These co-occurring stressors caused unprecedented and long-lasting decline of a previously robust kelp forest in northern California (Rogers-Bennett & Catton 2019), where sea otters are also absent. Given the biogeographical similarity between southern Oregon and northern California and the observation made by Dylan in 2018, we decided to undertake an analysis of the eight years of data collected during the TOPAZ/JASPER project in Port Orford starting in 2016, to investigate the trends of four trophic levels (purple sea urchins, bull kelp, zooplankton, and gray whales) across space and time. The results of our study were published last week in Scientific Reports and I am excited to be able to share them with you today.
Fig 2a. Sea urchins actively feeding on kelp stalks. Source: GEMM Lab.Fig 2b. Diver holding a sunflower sea star near Port Orford, OR. Source: Scott Groth.
Every day during the TOPAZ/JASPER field season, two teams head out to collect data. One team is responsible for tracking gray whales from shore using a theodolite, while the other team heads out to sea on a tandem research kayak to collect prey data (Figure 3). The kayak team samples prey in multiple ways, including dropping a GoPro camera at each sampling station. When the project was first developed, the original goal of these GoPro videos was to measure the relative abundance of prey. Since the sampling stations occur on or near reefs that are shallow with dense surface kelp, traditional methods to assess prey density, such as using a boat with an echosounder, are not suitable options. Instead, GEMM Lab PI Leigh Torres, together with the first Master’s student on this project Florence Sullivan, developed a method to score still images extracted from the GoPro videos to estimate relative zooplankton abundance. However, after we saw those images of urchins feeding on kelp in 2018, we decided to develop another protocol that allowed us to use these GoPro videos to also characterize sea urchin coverage and kelp condition. Once we had occurrence values for all four species, we were able to dig into the spatiotemporal trends.
Figure 3. Map of Port Orford, USA study area showing the 10 kayak sampling stations (white circles) within the two study sites (Tichenor Cove and Mill Rocks). The white triangle represents the cliff top location where theodolite tracking of whales was conducted. Figure and caption taken from Hildebrand et al. 2024.
When we examined the trends for each of the four study species across years, we found that purple sea urchin coverage in both of our study sites within Port Orford increased dramatically within our study period (Figure 4). In 2016, the majority of our sampled stations contained no visible urchins. However, by 2020, we detected urchins at every sampling station. For kelp, we saw the reverse trend; in 2016 all sampling stations contained kelp that was healthy or mostly healthy. But by 2019, there were many stations that contained kelp in poor health or where kelp was absent entirely. Zooplankton and gray whales experienced similar temporal trends as the kelp, with their occurrence metrics (abundance and foraging time, respectively) having higher values at the start of our study period and declining steadily during the eight years. While the rise in urchin coverage across our study area occurred concurrently with the decrease in kelp condition, zooplankton abundance, and gray whale foraging, we wanted to explicitly test how these species are related to one another based on prior ecological knowledge.
Figure 4. Temporal trends of purple sea urchin coverage, bull kelp condition, relative zooplankton abundance, and gray whale foraging time by year across the eight-year study period (2016–2023), from the generalized additive models. The colored ribbons represent approximate 95% confidence intervals. Line types represent the two study sites, Mill Rocks (MR; solid) and Tichenor Cove (TC; dashed). All curves are statistically significant (P < 0.05). Figure and caption taken from Hildebrand et al. 2024.
To test whether urchin coverage had an effect on kelp condition, we hypothesized that increased urchin coverage would be correlated with reduced kelp condition based on the decades of research that has established a negative relationship between the two when a trophic cascade occurs in kelp forest systems. Next, we wanted to test whether kelp condition had an effect on zooplankton abundance and hypothesized that increased kelp condition would be correlated with increased zooplankton abundance. We based this hypothesis on several pieces of prior knowledge, particularly as they pertain to mysid shrimp: (1) high productivity within kelp beds provides food for mysids, including kelp zoospores (VanMeter & Edwards 2013), (2) current velocities are one third slower inside kelp beds compared to outside (Jackson & Winant 1983), which might support the retention of mysids within kelp beds since they are not strong swimmers, and (3) the kelp canopy may serve as potential protection for mysids from predators (Coyer 1984). Finally, we wanted to test whether both kelp condition and zooplankton abundance have an effect on gray whales and we hypothesized that increased values for both would be correlated with increased gray whale foraging time. While the reasoning behind our hypothesized correlation between zooplankton prey and gray whales is obvious (whales eat zooplankton), the reasoning behind the kelp-whale connection may not be. We speculated that since kelp habitat may aggregate or retain zooplankton prey, gray whales may use kelp as an environmental cue to find prey patches.
When we tested our hypotheses through generalized additive models, we found that increased urchin coverage was significantly correlated with decreased kelp condition in both study sites, providing evidence that a shift from a kelp forest to an urchin barren may have occurred in the Port Orford area. Additionally, increased kelp condition was correlated with increased zooplankton abundance, supporting our hypothesis that kelp forests are an important habitat and resource for nearshore zooplankton prey. Interestingly, this relationship was bell-shaped in one of our two study sites, suggesting that there are other factors besides healthy bull kelp that influence zooplankton abundance, which likely include upwelling dynamics, habitat structure, and local oceanographic characteristics. For the whale model, we found that increased kelp condition was significantly correlated with increased gray whale foraging time, which may corroborate our hypothesis that gray whales use kelp as an environmental cue to locate prey. Zooplankton abundance was significantly correlated with gray whale foraging time in one of our two sites. Once again, this relationship was bell-shaped, which suggests other factors influence gray whale foraging time, including prey quality (Hildebrand et al. 2022) and density.
Figure 5. Effects derived from trophic path generalized additive models of purple sea urchin coverage on kelp condition (A), kelp condition on relative zooplankton abundance (B), and kelp condition and relative zooplankton abundance on gray whale foraging time (C). The colored ribbons represent approximate 95% confidence intervals. Line types represent the two study sites, Mill Rocks (MR; solid) and Tichenor Cove (TC; dashed). Curves with asterisks indicate statistically significant (P < 0.05) relationships. Figure and caption taken from Hildebrand et al. 2024.
Our results highlight the potential larger impacts of reduced gray whale foraging time as a result of these trophic dynamics may cause at the individual and population level. If an area that was once a reliable source of food (like Port Orford) is no longer favorable, then whales likely search for other areas in which to feed. However, if the areas affected by these dynamics are widespread, then individuals may spend more time searching for, and less time consuming, prey, which could have energetic consequences. While our study took place in a relatively small spatial area, the trophic dynamics we documented in our system may be representative of patterns across the PCFG range, given ecological and topographic similarities in habitat use patterns. In fact, in the years with the lowest kelp, zooplankton, and whale occurrence (2020 and 2021) in Port Orford, the GRANITE field team also noted low whale numbers and minimal surface kelp extent in the central Oregon field site off of Newport. However, ecosystems are resilient. We are hopeful that the dynamics we documented in Port Orford are just short-term changes and that the system will return to its former balanced state with less urchins, more healthy bull kelp, zooplankton, and lots of feeding gray whales.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly message when we post a new blog. Just add your name and email into the subscribe box below.
References
Coyer, J. A. (1984). The invertebrate assemblage associated with the giant kelp, Macrocystis pyrifera, at Santa Catalina Island, California: a general description with emphasis on amphipods, copepods, mysids, and shrimps. Fishery Bulletin, 82(1), 55-66.
Harvell, C. D., Montecino-Latorre, D., Caldwell, J. M., Burt, J. M., Bosley, K., Keller, A., … & Gaydos, J. K. (2019). Disease epidemic and a marine heat wave are associated with the continental-scale collapse of a pivotal predator (Pycnopodia helianthoides). Science advances, 5(1), eaau7042.
Hildebrand, L., Sullivan, F. A., Orben, R. A., Derville, S., & Torres, L. G. (2022). Trade-offs in prey quantity and quality in gray whale foraging. Marine Ecology Progress Series, 695, 189-201.
Jackson, G. A., & Winant, C. D. (1983). Effect of a kelp forest on coastal currents. Continental Shelf Research, 2(1), 75-80.
Jacox, M. G., Alexander, M. A., Mantua, N. J., Scott, J. D., Hervieux, G., Webb, R. S., & Werner, F. E. (2018). Forcing of multi-year extreme ocean temperatures that impacted California Current living marine resources in 2016. Bull. Amer. Meteor. Soc, 99(1).
Kone, D. V., Tinker, M. T., & Torres, L. G. (2021). Informing sea otter reintroduction through habitat and human interaction assessment. Endangered Species Research, 44, 159-176.
Rogers-Bennett, L., & Catton, C. A. (2019). Marine heat wave and multiple stressors tip bull kelp forest to sea urchin barrens. Scientific reports, 9(1), 15050.
Torres, L. G., Nieukirk, S. L., Lemos, L., & Chandler, T. E. (2018). Drone up! Quantifying whale behavior from a new perspective improves observational capacity. Frontiers in Marine Science, 5, 319.
VanMeter, K., & Edwards, M. S. (2013). The effects of mysid grazing on kelp zoospore survival and settlement. Journal of Phycology, 49(5), 896-901.
Understanding how baleen whales are affected by human activity is a central goal for many research projects in the GEMM Lab. The overarching goal of the GRANITE (Gray whale Response to Ambient Noise Informed by Technology and Ecology) project is to quantify baleen whale physiological response to different stressors (e.g., boat presence and noise) and model the subsequent impacts of these stressors on the population. We will achieve this goal by implementing our long-term, replicate dataset of Pacific Coast Feeding Group (PCFG) gray whales into a framework called population consequences of disturbance (PCoD). I will not go into the details of PCoD in this blog (but I wrote a post a few years ago that you can revisit). Instead, I will explain the approach I am taking to assess where and when individual whales spend time in our study area, which will form an essential component of PCoD and be one of the chapters of my PhD dissertation.
Individuals in a population are unlikely to be exposed to a stressor in a uniform way because they make decisions differently based on intrinsic (e.g., sex, age, reproductive status) and extrinsic (e.g., environment, prey, predators) factors (Erlinge & Sandell 1986). For example, a foraging female gray whale who is still nursing a calf will need to consider factors that are different to ones that an adult single male might need to consider when choosing a location to feed. These differences in decision-making exist across the whole population, which makes it important to understand where individuals are spending time and how they overlap with stressors in space and time before trying to quantify the impacts of stressors on the population as a whole (Pirotta et al. 2018). I am currently working on an analysis that will determine an individual’s exposure to a number of stressors based on their space use patterns.
We can monitor space use patterns of individuals in a population through time using spatial capture-recapture techniques. As the name implies, a spatial capture-recapture technique involves capturing an individual in a marked location during a sampling period, releasing it back into the population, and then (hopefully) re-capturing it during another sampling period in the future, at either the same or a different location. With enough repeat sampling events, the method should build spatial capture histories of individuals through time to better understand an individual’s space use patterns (Borchers & Efford 2008). While the use of the word capture implies that the animal is being physically caught, this is not necessarily the case. Individuals can be “captured” in a number of non-invasive ways, including by being photographed, which is how we “capture” individual PCFG gray whales. These capture-recapture methods were first pioneered in terrestrial systems, where camera traps (i.e., cameras that take photos or videos when a motion sensor is triggered) are set up in a systematic grid across a study area (Figure 1; Royle et al. 2009, Gray 2018). Placing the cameras in a grid system ensures that there is an equal distribution of cameras throughout the study area, which means that an animal theoretically has a uniform chance of being captured. However, because we know that individuals within a population make space use decisions differently, we assume that individuals will distribute themselves differently across a landscape, which will manifest as individuals having different centers of their spatial activity. The probability of capturing an individual is highest when a camera trap is at that individual’s activity center, and the cameras furthest away from the individual’s activity center will have the lowest probability of capturing that individual (Efford 2004). By using this principle of probability, the data generated from spatial capture-recapture field methods can be modelled to estimate the activity centers and ranges for all individuals in a population. The overlap of an individual’s activity center and range can then be compared to the spatiotemporal distribution of stressors that an individual may be exposed to, allowing us to determine whether and how an individual has been exposed to each stressor.
Figure 1. Example of camera trap grid in a study area. Figure taken from Gray (2018).
While capture-recapture methods were first developed in terrestrial systems, they have been adapted for application to marine populations, which is what I am doing for our GRANITE dataset of PCFG gray whales. Together with a team of committee members and GRANITE collaborators, I am developing a Bayesian spatial capture-recapture model to estimate individual space use patterns. In order to mimic the camera trap grid system, we have divided our central Oregon coast study area into latitudinal bins that are approximately 1 km long. Unfortunately, we do not have motion sensor activated cameras that automatically take photographs of gray whales in each of these latitudinal bins. Instead, we have eight years of boat-based survey effort with whale encounters where we collect photographs of many individual whales. However, as you now know, being able to calculate the probability of detection is important for estimating an individual’s activity center and range. Therefore, we calculated our spatial survey effort per latitudinal bin in each study year to account for our probability of detecting whales (i.e., the area of ocean in km2 that we surveyed). Next, we tallied up the number of times we observed every individual PCFG whale in each of those latitudinal bins per year, thus creating individual spatial capture histories for the population. Finally, using just those two data sets (the individual whale capture histories and our survey effort), we can build models to test a number of different hypotheses about individual gray whale space use patterns. There are many hypotheses that I want to test (and therefore many models that I need to run), with increasing complexity, but I will explain one here.
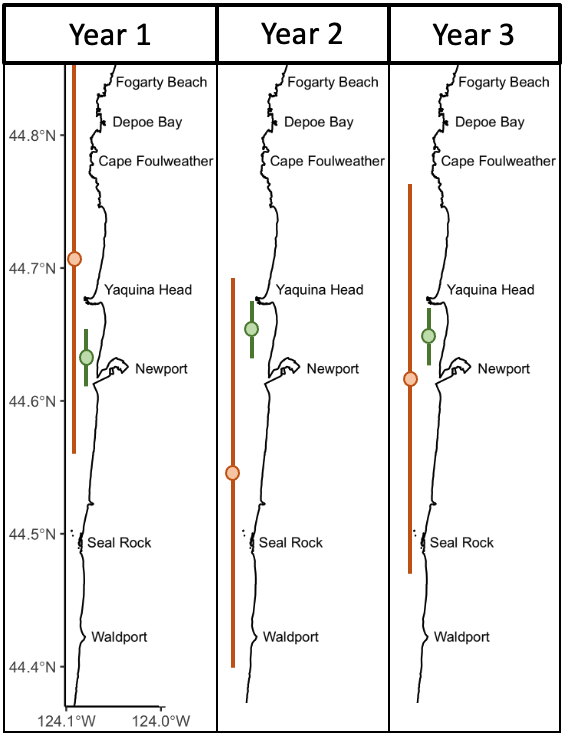
Over eight years of field work for the GRANITE project, consisting of over 40,000 km2 of ocean surveyed with 2,169 sightings of gray whales, our observations lead us to hypothesize that there are two broad space use strategies that whales use to optimize how they find enough prey to meet their energetic needs. For the moment, we are calling these strategies ‘home-body’ and ‘roamer’. As the name implies, a home-body is an individual that stays in a relatively small area and searches for food in this area consistently through time. A roamer, on the other hand, is an individual that travels and searches over a greater spatial area to find good pockets of food and does not generally tend to stay in just one place. In other words, we except a home-body to have a consistent activity center through time and a small activity range, while a roamer will have a much larger activity range and its activity center may vary more throughout the years (Figure 2).
Figure 2. Schematic representing one of the hypotheses we will be testing with our Bayesian spatial capture-recapture models. The schematic shows the activity centers (the circles) and activity ranges (vertical lines attached to the circles) of two individuals (green and orange) across three years in our central Oregon study area. The green individual represents our hypothesized idea of a home-body, whereas the orange individuals represents our idea of a roamer.
While this hypothesis sounds straightforward, there are a lot of decisions that I need to make in the Bayesian modeling process that can ultimately impact the results. For example, do all home-bodies in a population have the same size activity range or can the size vary between different home-bodies? If it can vary, by how much can it vary? These same questions apply for the roamers too. I have a long list of questions just like these, which means a lot of decision-making on my part, and that long list of hypotheses I previously mentioned. Luckily, I have a fantastic team made up of Leigh, committee members, and GRANITE collaborators that are guiding me through this process. In just a few more months, I hope to reveal how PCFG individuals distribute themselves in space and time throughout our central Oregon study area, and hence describe their exposure to different stressors. Stay tuned!
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly message when we post a new blog. Just add your name and email into the subscribe box below.
References
Borchers DL, Efford MG (2008) Spatially explicit maximum likelihood methods for capture-recapture studies. Biometrics 64:377-385.
Efford M (2004) Density estimation in live-trapping studies. Oikos 106:598-610.
Erlinge S, Sandell M (1986) Seasonal changes in the social organization of male stoats, Mustela erminea: An effect of shifts between two decisive resources. Oikos 47:57-62.
Gray TNE (2018) Monitoring tropical forest ungulates using camera-trap data. Journal of Zoology 305:173-179.
Pirotta E, Booth CG, Costa DP, Fleishman E, Kraus SD, and others (2018) Understanding the population consequences of disturbance. Ecology and Evolution 8(19):9934–9946.Royle J, Nichols J, Karanth KU, Gopalaswamy AM (2009) A hierarchical model for estimating density in camera-trap studies. Journal of Applied Ecology 46:118-127.
Another year has come and gone, and the GEMM Lab has expanded and accomplished in many facets! Every year it gets just a little bit harder to succinctly summarize all of the research, outreach, and successes that the GEMMs accomplish and this year we are trying something a little different for our Year in the Life tradition. Rather than summarizing what the GEMM Lab has been up to thematically, we have decided to let everyone tell you their 2023 recap in their own words. So, snuggle up with your favorite holiday drink and enjoy our 8th edition of the Year in the Life!
Leigh
As the captain at the helm of the GEMM Lab ship, Leigh plays a major role in all of our accomplishments and celebrates them right along with us. She returned to Oregon after a very well-deserved, refreshing and reflective sabbatical in New Zealand, where she spent three months traveling the north and south islands with her family. One highlight of her trip was when Leigh went paddleboarding in Tarakena Bay and was surrounded by common dolphins the whole time, swimming around her and making eye contact! Almost immediately after her sabbatical, Leigh headed to the lagoons in Baja with Clara, which kick-started a busy year of field work as Leigh oversaw six different projects that involved field work throughout the year. In early summer, Leigh hosted her graduate advisor Dr. Andy Read as part of the OSU Hatfield Marine Science Center’s Lavern Weber Visiting Scientist program. Andy spent a jam-packed 10 days in Oregon, which included many meetings with Leigh and each GEMM project team as well as a fantastic first day on the water for the GRANITE project where Andy was introduced to the beloved PCFG gray whales! Another huge accomplishment for 2023 was Leigh’s successful funding application to the National Science Foundation for the SAPPHIRE project with Dawn and KC (more below), which will see the team head off for more blue whale research in New Zealand in January 2024!
Ale
For postdoctoral scholar Alejandro A. Fernández Ajó, a big highlight of 2023 was the 61 fecal samples from 25 individual gray whales collected by the GRANITE team, with most samples originating from known whales that regularly visit the Oregon coast. This presents a unique opportunity to study changes and track these individual whales across seasons and years, allowing us to observe variations in their reproductive health, body condition, and responses to stressors such as vessel noise and entanglements. Currently, Alejandro is back at his graduate institute, Northern Arizona University (NAU), conducting lab work to analyze these fecal samples. Monitoring endocrine biomarkers (hormones) enables us to understand how Pacific Coast Feeding Group (PCFG) whales respond to stressors, providing insights into different aspects of the PCFG gray whale’s biology and physiology.
In addition, Alejandro led research this year that assessed diagnostic tools for non-invasive pregnancy diagnosis, and proposed a methodological approach for identifying pregnancies in gray whales. He also taught as a guest lecturer in the grad-level course ‘Conservation Physiology’ at NAU and started mentoring Camila Muñoz Moreda, a PhD student in Argentina, investigating stressors impacting Southern Right Whales’ health in Patagonia. Alejandro was also invited and awarded a travel grant to participate in a workshop to be held in Kruger, South Africa, where a group of 20 leading experts will gather to discuss research approaches and resources that are needed for future comparative physiology research in a changing world.
Allison
Master’s student Allison Dawn started the year off by taking two challenging SCUBA courses, first honing skills like underwater navigation and completing a 100 ft dive in the Hood Canal as part of Advanced Diving. Her favorite memory was seeing a Giant Pacific Octopus with dive buddy and fellow MMI marine scientist Kyra Bankhead! Next, Allison passed her Rescue Dive training where she learned best practices for effective rescue & emergency response while working in the water. During this time, she also completed her Master’s thesis, titled “Intermittent upwelling impacts zooplankton and their gray whale predators at multiple scales” which she successfully defended this past June. Afterwards, Allison led another successful field season in Port Orford for the 9th consecutive year of the TOPAZ/JASPER projects, where she mentored two high school students, one undergraduate SCUBA diver, and one NSF REU undergrad. Read their individual blogs and all about the exciting season here!
Clara
2023 started off with a big data processing milestone for Clara – she finished annotating all seven years of drone footage for her PhD! She started working on this in her first year, so to finally have her completed dataset was momentous – and meant that she could get to work on analysis and writing. While nothing is yet published, her first chapter is under review and the second and third are both underway. She also presented the results of her first chapter, focused on individual specialization in PCFG gray whale foraging behaviors, at the Animal Behavior Society conference in July. In addition, Clara’s former REU intern, Celest Sorrentino, was in attendance, and Clara enjoyed mentoring Celest through her first scientific conference. Actually, Celest came back for the whole summer as a research assistant, processing data from Clara and Leigh’s trip to Baja California Sur, Mexico in March (read about Celest’s summer here).
Clara also taught her photogrammetry lab for Renee Albertson’s marine megafauna course for the fourth year in a row and gave outreach talks for the American Cetacean Society Oregon Chapter and the Cape Perpetua Land Sea Symposium. And an update wouldn’t be complete without mentioning field work! Clara participated in the GRANITE project’s eighth field season (her fourth). An absolute highlight was her trip to Baja with Leigh where she collected incredible drone footage and crossed paths with a known whale to the GEMM lab, Pacman! As she works through the final year of her PhD, Clara is excited to continue exploring this incredible behavior data set and learning more about these whales!
Dawn
Through the EMERALD project, postdoctoral scholar Dawn Barlow has been busy examining habitat use, distribution, and abundance of gray whales and harbor porpoises in the Northern California Current over three decades. This project has documented long-term hotspots in gray whale habitat, illustrated regional differences in overlap between harbor porpoises and different fish species, and explored the importance of upwelling dynamics for these nearshore cetaceans. Dawn presented findings at the Effects of Climate Change on the World’s Ocean (ECCWO) conference in Bergen, Norway, which was a fruitful opportunity to connect with researchers from around the world and across disciplines.
More exciting news came in spring 2023, when a GEMM Lab team was granted funding from the National Science Foundation for the project “Marine Predator and Prey Response to Climate Change: Synthesis of Acoustics, Physiology, Prey, and Habitat in a Rapidly Changing Environment (SAPPHIRE)”. Through the SAPPHIRE project, we will examine how changing ocean conditions affect the availability and quality of krill, and thus impact blue whale behavior, health, and reproduction. Dawn and the team are busily preparing to head to Aotearoa New Zealand to find krill and blue whales for our first field season in January!
Throughout 2023, Dawn also had the opportunity to conduct fieldwork here in our Oregon backyard aboard the R/V Pacific Storm for the HALO project, in the skies aboard USCG helicopters for the OPAL project, and as chief scientist of a research cruise for the MOSAIC project. She also had the pleasure of working with undergraduate student Mariam Alsaid to document the occurrence patterns of the little-studied sei whale in Oregon waters. The fifth and final chapter of her PhD was published in early 2023, wrapping up a decade of research on New Zealand blue whales through the OBSIDIAN project. In December, a collaborative study led by Dawn was published comparing blue whale morphology and oceanography of foraging grounds in California, New Zealand, and Chile. As 2023 comes to a close with various projects nearing completion, in full swing, and just beginning, Dawn looks forward to what 2024 will bring!
Kate
For Master’s student Kate Colson, a highlight of 2023 was teaching an introductory science class to first year undergraduate students at University of British Columbia (UBC). After shaping these young minds, she headed south and moved to Newport to be a part of the GRANITE field team and reunite with the PCFG gray whales! Kate spent the summer working to process the season’s CATS tag deployments, and successfully defended her Master’s thesis in August. After spending the fall turning her thesis chapters into manuscripts, she submitted her first scientific paper, and will be ready to submit her second early in the new year! And, after another season of beautiful Oregon beach walks, Kate finally found a trophy agate from the Oregon coast (see photo). Kate recently moved back to the east coast and has started a new research assistant job working with Dtag data, further developing the tag analysis skills she learned in her master’s program.
KC
This year was productive on many levels for postdoctoral scholar KC Bierlich! He published seven research papers, with an additional three currently in review, and was fortunate enough to receive several funding awards from a) the Marine Mammal Commission, supporting GRANITE fieldwork for the next two years, b) the National Science Foundation, funding the GEMM Lab’s new SAPPHIRE project, and c) the Office of Naval Research. This last grant will support KC launching MMI’s Center of Drone Excellence (CODEX), which focuses on developing analytical tools for processing and analyzing drone imagery, including user-friendly hardware and software. Some early highlights from CODEX includes major updates to the photogrammetry software MorphoMetriX and CollatriX, and the development of LidarBoX, a 3D printed altimeter hardware system that can attach to several types of commercial drones and help improve the accuracy of altitude measurements.
KC mentored seven students this year (two high school, two masters, and three undergraduates), and was awarded the “Excellence in Undergraduate Research Mentoring by a Postdoc” by OSU. He had a busy summer with another great GRANITE field season, and partnered with the Innovation Lab (iLab) at HMSC to develop a system for dropping tags onto whales using drones. The team successfully tested this tagging system on a stand-up paddle board, and next will employ the tags while studying Pygmy blue whales in January and February for SAPPHIRE. And most importantly, KC became a dad; Caroline Marie Bierlich was born in early September. KC and Colette have been absolutely overjoyed with their new role as parents!
Lisa
A big milestone was reached by Lisa in the first quarter of 2023 when she successfully passed her written and oral exams, allowing her to advance to PhD candidacy! Her committee members gave Lisa lots of food for thought in the many scientific papers and book chapters assigned to her during her study period, ranging in topic from Bayesian ecological modeling to baleen whale energetics to Pacific Ocean oceanographic dynamics to foundational foraging theory, all of which will help Lisa as she now works to accomplish her proposed PhD research in the next couple of years. Lisa was once again part of the GRANITE field team this summer, providing her the opportunity to spend over 130 magical hours with the beloved (and by now very well known to the team) PCFG gray whales! Together with KC, Lisa greatly enjoyed mentoring two high school interns, Hali Peterson and Isaac Cancino, during the summer as they assisted her with zooplankton identification and sorting. Hali, who lives and goes to school close to Newport, has continued working with Lisa for the GEMM Lab into the fall, helping with a number of tasks. Lisa was involved in five publications this year, of which she is probably most proud of the paper published in Current, the Journal of Marine Education, where together with Leigh and Tracy Crews (the Associate Director for Education at the Oregon Sea Grant’s STEM Hub), she laid out the roadmap, successes, and hurdles associated with JASPER, the STEM component of the paired TOPAZ/JASPER projects in Port Orford, Oregon. The project has graduated a total of 31 students and Lisa is immensely proud to have been part of this project that will forever remain near and dear to her heart.
Marissa
PhD student Marissa Garcia’s memories of the year take her back to early mornings driving down the Pacific Coast Highway, the Pacific Ocean as the backdrop to her daily commute to the Hatfield Marine Science Center. For Marissa, the highlight of 2023 was the extended stay — or “PhD sabbatical” — she carved out of the routine summer fieldwork for the HALO Project. Following a July crash course in modeling with Dawn, Soléne, and Leigh, Marissa implemented an oceanographic analysis to share at the Acoustics 23 conference in Sydney, Australia in December. Another highlight from her year was co-organizing the oral session “All Ears In: Advancing Ecology and Conservation with Bioacoustics” at the Ecological Society of America conference over the summer. Earlier in the year, Marissa was selected as an NSF GRFP Fellow as well as an Animal Bioacoustics representative for the Acoustical Society of America’s Student Council. Marissa is proud of the skillsets she gained this year: wrangling large acoustic data sets, running click detectors, wading through oceanographic variables, and setting her sights on species distribution modeling. This upcoming year, she looks forward to challenging herself to grow even more!
Nat
Although new PhD student Natalie Chazal is ending this year at Oregon State University, she actually started 2023 at North Carolina State University, where she defended her Master’s thesis in the spring. Over the summer, Nat submitted both of her thesis chapters for publication, and then moved to Oregon, spending a couple weeks in Newport where she got a taste of fieldwork in the GEMM Lab. During the fall term, Nat took Bayesian statistics with MMI professor Josh Stewart, where she dug into zooplankton tow data from the past 3 years of GRANITE work. She also took a few orientation courses that helped her understand the resources available at OSU and how to best prepare for the journey ahead. In between all of her classwork, TA grading, and research, she has explored the Pacific Northwest with hikes to Mount St. Helens and Mount Hood, birding on Mary’s Peak and Yaquina Head Outstanding Natural Area, and visiting waterfalls near Portland and Silver Falls State Park.
Rachel
2023 was a far-ranging year for PhD student Rachel Kaplan! Skipping out on the beautiful Oregon summer, she instead spent a six-month winter field season at Palmer Station, the smallest of the U.S. research bases in Antarctica. Working with her CEOAS co-advisor Kim Bernard and undergraduate student Abby Tomita, Rachel loved studying Antarctic krill through at-sea fieldwork and long-term experiments, with plenty of crafting and skiing through the long polar night. Now, she is thrilled to be back with her Oregon krill and labmates. Rachel is happy to be closing out the year with the acceptance of her first PhD chapter for publication, and is excited for all that 2024 will hold!
Solène
After almost three years of working remotely as a postdoctoral scholar, Solène Derville finally made it to Oregon! She spent a year in Newport, mainly working on the OPAL and SLATE projects that address the issue of whale entanglements off the coast of Oregon. Solène contributed to several GEMM Lab milestones this year, including finalizing the first phase of OPAL with a publication of a study investigating how the exposure of rorqual whales to Dungeness crab fishing gear varies in time and space (Derville et al. 2023 in Biological Conservation) and publishing an isotope-based analysis of southern right whale feeding ground distribution over the whole Southern Ocean (Derville et al. 2023 in PNAS). Being in Newport in person offered a lot more opportunities to participate in fieldwork (April STEM cruises, September NCC cruise, small-boat rorqual whale biopsy and photo-ID work) and academic life (co-teaching a graduate course on the Spatial Ecology of Marine Megafauna with Leigh and Dawn). She also got to explore the marvels of Oregon’s amazing outdoors… from climbing at Smith Rock, or skiing in the cascades, to hanging out with blue whales… all in the good company of GEMM Lab friends!
Dear reader…
Thank you, dear reader, for taking the time to review the year with us! You have once again been awesome, supportive viewers of our blog, with a whopping 25,893 views of our blog this year!! We wish you all restful, happy, and most importantly, healthy holidays, and hope you will join us again in 2024!
The GEMM Lab with their white elephant gifts during our annual holiday party
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly message when we post a new blog. Just add your name and email into the subscribe box below.
Baleen whales face a multitude of threats on a daily basis. The exposure to some of these threats can be assessed visually. For example, the presence of propeller scars on a whale are indicative that the individual was struck by a boat. However, there are some threats that are not easily detected from visual assessments. One of these threats is the ingestion of microparticles (MPs), which include microplastics and other anthropogenic debris. While MP research has entered its second decade and documentation of MPs in the marine environment is common, we still lack empirical information on the rates of MP ingestion by baleen whales and their prey. Hence, one of the objectives of the Coastal Oregon Zooplankton Investigation (COZI; read more about it in a previous blog), which GEMM Lab PI Leigh Torres led, was to determine to what extent Pacific Coast Feeding Group (PCFG) gray whales and their nearshore zooplankton prey are impacted by MPs. The results of this work were recently published in the journal Frontiers in Marine Science and I am going to summarize them for you here today.
A number of studies have documented MP ingestion in baleen whales, including in humpback (Besseling et al., 2015), fin (Fossi et al., 2012, 2014, 2016, 2017), Bryde’s, and sei whales (Zantis et al., 2022). The effects of ingesting MPs on baleen whales are theorized to include blockage of internal organs, mechanical damage of the digestive tract, false feeling of satiation (full from eating), and potentially leaching of toxicants depending on the length of the digestive period (Donohue et al., 2019; Hudak & Sette 2019; Zhu et al., 2019; Novillo et al., 2020). Despite the fact that MPs have been documented in a number of baleen whale species, there is still a lack of knowledge regarding MP ingestion rates by baleen whales from empirical data, although modeled estimates have been derived for a few species (Zantis et al., 2022; Kahane-Rapport et al., 2022). Basically, we know whales eat MPs because it has been detected in their stomachs, but we do not know how much MPs they consume. The COZI team therefore aimed to quantify baleen whale MP consumption rates from empirically counted MP loads in zooplankton prey and to look at MP exposure of baleen whales from “zoop to poop” (Figure 1).
Figure 1 Schematic depicting our “zoop to poop” approach. Taken from Torres et al., 2023.
In order to accomplish this aim, we used “zoop” and “poop” samples collected between 2017 to 2019 during the GEMM Lab’s long-term GRANITE (Gray whale Response to Ambient Noise Informed by Technology and Ecology) project. We analyzed MP loads in three prey zooplankton species found in nearshore Oregon waters (the amphipod Atylus tridens and the mysid shrimp Holmesimysis sculpta and Neomysis rayii), all of which are known PCFG gray whale prey (Hildebrand et al., 2021), as well as five fecal samples collected from four unique individual gray whales. While the field collection of these samples was led by the GEMM Lab, the processing and MP analysis was led by Dr. Susanne Brander and conducted by a number of undergraduate student workers. MP analysis is no easy feat as it involves many, many meticulous and time-intensive steps in order to get from a sample of gray whale prey or poop to a known number of MPs that the sample contained. The process involves (1) sorting and identifying the prey into the different species; (2) rinsing the individuals to ensure no external MPs are counted; (3) digesting the sample in potassium hydroxide (KOH) for 24-72 hours; (4) sieving and filtering the digested samples; (5) picking out suspected MPs from the filters and measuring them; (6) analyzing the suspected MPs to confirm chemical composition. On top of all of these steps, anyone working with the samples has to try and minimize potential MP contamination, which is not easy since MPs are practically everywhere, such as synthetic fibers from our clothes or microplastics that are floating around in the air.
Figure 2 Microparticle (MP) loads and morphotypes by zooplankton species. (A) the number of MPs per 1 gram per species, with the dotted line representing the average MP level in controls. (B) the proportion of MP morphotypes found in each zooplankton species. (C) the proportion of Fourier transform infrared (FTIR) spectroscopy categories of MPs found in each zooplankton species. The sample size for each sample is denoted above all columns.Taken fromTorres et al., 2023.
After many long years of lab work (COVID lab restrictions included), we are excited (and a little daunted) to share the results of this collaborative project with you. We detected MPs in all 26 zooplankton prey samples that we analyzed and found that the number of MPs in the three species were pretty similar, with an average of 4 MPs per gram of zooplankton (Figure 2). Over 50% of the 418 suspected MPs that we identified in the zooplankton samples were fibers. We also detected MPs in all five gray whale fecal samples that we analyzed. While we also detected fibers among the 37 suspected MPs pulled from the fecal samples, we found a higher proportion of larger MPs such as fragments and pellets in the “poop” samples, than we did in the “zoop” samples (Figure 3). We also tested some seawater samples as controls to see how the composition of MPs in seawater compared to that of zooplankton and gray whale feces. We found that seawater was dominated by fibers, similar to the zooplankton prey. This finding suggests that the larger MPs (e.g., fragments, pellets) that we found in gray whale feces must be coming from somewhere other than their prey and the ambient seawater. This led us to hypothesize that gray whales are likely exposed to MPs through two pathways, via (1) trophic transfer from their zooplankton prey and (2) indiscriminate consumption of ambient MPs in the benthos while foraging benthically (Figure 1).
Figure 3 Microparticle (MP) loads and morphotypes found in each of the five gray whale fecal samples analyzed. (A)the number of MPs per gram of fecal sample, with the dotted line representing the average MP level in controls. (B) the proportion of MP morphotypes found in each fecal sample. (C) the proportion of Fourier transform infrared (FTIR) spectroscopy categories of MPs found in each fecal sample. The sample size for each sample is denoted above all columns. Taken from Torres et al., 2023.
Next we wanted to estimate the daily ingestion rates of MPs by gray whales. For this estimation, we used our known values of zooplankton MP ingestion (from our analyzed samples) and extrapolated them using daily energetic needs of gray whales (i.e., how many calories does the whale need each day). The only published values of daily gray whale caloric needs are for pregnant and lactating females (Villegas-Amtmann et al., 2015, 2017), which is why we were only able to estimate daily MP ingestion rates for these two demographic groups. The numbers we calculated were rather staggering (and led us to double-, triple-, and quadruple-check our math) as we estimate that if a pregnant gray whale only ate the mysid N. rayii in a day, she would consume 9.55 million MP per day. We made these estimates for all three prey species that we analyzed as well as a “composite preyscape” (an average of the three prey species) and you can see all of those results in Table 1.
Table 1 Estimates of the number of microparticles (MPs) that a pregnant and lactating female gray whale consumes per day generated through extrapolation of results from this study (Microparticles per individual zooplankton; first row) to their daily energetic needs by zooplankton prey species from Hildebrand et al., 2021. Taken from Torres et al., 2023.
These results are frightening. They still are to me even though I have spent months with this knowledge after having done a lot of the data analysis for this project. I think it is particularly frightening to think about the fact that MPs are not the only anthropogenic threat that gray whales (and really any organism in the ocean) are exposed to. The good news is that you can do something to help reduce this threat in the oceans. Below are just a few suggestions of what you can do to reduce MP pollution to the environment:
A major source of pollution in the ocean comes from microfibers through our laundry (as you saw in our results). You can help stop this pathway by simply using a Cora Ball or installing a filter (such as this one) in your washing machine that captures microfleece & polyester fibers.
Minimize your use of single-use plastics. There are so many ways to do so including reuseable water bottles, travel mugs for coffee or tea, fabric totes as shopping bags, carry a set of utensils for takeout food, beeswax wraps instead of plastic wrap or sandwich bags.
Use public transport when possible as another huge source of microplastics comes from tire treads! This solution also helps reduce your carbon footprint.
References
Besseling E., Foekema E. M., Van Franeker J. A., Leopold M. F., Kühn S., Bravo Rebolledo E. L., et al. (2015). Microplastic in a macro filter feeder: humpback whale Megaptera novaeangliae. Mar. pollut. Bull. 95, 248–252. doi: 10.1016/j.marpolbul.2015.04.007
Donohue M. J., Masura J., Gelatt T., Ream R., Baker J. D., Faulhaber K., et al. (2019). Evaluating exposure of northern fur seals, callorhinus ursinus, to microplastic pollution through fecal analysis. Mar. pollut. Bull. 138, 213–221. doi: 10.1016/j.marpolbul.2018.11.036
Fossi M. C., Panti C., Guerranti C., Coppola D., Giannetti M., Marsili L., et al. (2012). Are baleen whales exposed to the threat of microplastics? a case study of the Mediterranean fin whale (Balaenoptera physalus). Mar. pollut. Bull. 64, 2374–2379. doi: 10.1016/j.marpolbul.2012.08.013
Fossi M. C., Coppola D., Baini M., Giannetti M., Guerranti C., Marsili L., et al. (2014). Large Filter feeding marine organisms as indicators of microplastic in the pelagic environment: the case studies of the Mediterranean basking shark (Cetorhinus maximus) and fin whale (Balaenoptera physalus). Mar. Environ. Res. 100, 17–24. doi: 10.1016/j.marenvres.2014.02.002
Fossi M. C., Marsili L., Baini M., Giannetti M., Coppola D., Guerranti C., et al. (2016). Fin whales and microplastics: the Mediterranean Sea and the Sea of cortez scenarios. Environ. pollut. 209, 68–78. doi: 10.1016/j.envpol.2015.11.022
Fossi M. C., Romeo T., Baini M., Panti C., Marsili L., Campani T., et al. (2017). Plastic debris occurrence, convergence areas and fin whales feeding ground in the Mediterranean marine protected area pelagos sanctuary: a modeling approach. Front. Mar. Sci. 4. doi: 10.3389/fmars.2017.00167
Hildebrand L., Bernard K. S., Torres L. G. (2021). Do gray whales count calories? comparing energetic values of gray whale prey across two different feeding grounds in the eastern north pacific. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.683634
Hudak C. A., Sette L. (2019). Opportunistic detection of anthropogenic micro debris in harbor seal (Phoca vitulina vitulina) and gray seal (Halichoerus grypus atlantica) fecal samples from haul-outs in southeastern Massachusetts, USA. Mar. pollut. Bull. 145, 390–395. doi: 10.1016/j.marpolbul.2019.06.020
Kahane-Rapport S. R., Czapanskiy M. F., Fahlbusch J. A., Friednlaender A. S., Calambokidis J., Hazen E. L., et al. (2022). Field measurements reveal exposure risk to microplastic ingestion by filter-feeding megafauna. Nat. Commun. 13, 6327. doi: 10.1038/s41467-022-33334-5
Novillo O., Raga J. A., Tomás J. (2020). Evaluating the presence of microplastics in striped dolphins (Stenella coeruleoalba) stranded in the Western Mediterranean Sea. Mar. pollut. Bull. 160, 111557. doi: 10.1016/j.marpolbul.2020.111557
Torres, L. G., Brander, S. M., Parker, J. I., Bloom, E. M., Norman, R., Van Brocklin, J. E., Lasdin, K.S., Hildebrand, L. (2023) Zoop to poop: assessment of microparticle loads in gray whale zooplankton prey and fecal matter reveal high daily consumption rates. Front. Mar. Sci. https://doi.org/10.3389/fmars.2023.1201078
Villegas-Amtmann S., Schwarz L. K., Sumich J. L., Costa D. P. (2015). A bioenergetics model to evaluate demographic consequences of disturbance in marine mammals applied to gray whales. Ecosphere 6, 1–19. doi: 10.1890/ES15-00146.1
Villegas-Amtmann S., Schwarz L. K., Gailey G., Sychenko O., Costa D. P. (2017). East Or west: the energetic cost of being a gray whale and the consequence of losing energy to disturbance. Endangered Species Res.34, 167–183. doi: 10.3354/esr00843
Zantis L. J., Bosker T., Lawler F., Nelms S. E., O’Rorke R., Constantine R., et al. (2022). Assessing microplastic exposure of large marine filter-feeders. Sci. Total Environ. 818, 151815. doi: 10.1016/j.scitotenv.2021.151815
Zhu J., Yu X., Zhang Q., Li Y., Tan S., Li D., et al. (2019). Cetaceans and microplastics: first report of microplastic ingestion by a coastal delphinid, Sousa chinensis. Sci. Total Environ. 659, 649–654. doi: 10.1016/j.scitotenv.2018.12.389
When I wrote my first blog a year ago, I was just starting to dig into the field work and data analysis I had loftily proposed for my graduate degree. Now I am writing to you as a new Master of Science! A little more than a month ago, I successfully defended my thesis research where I used the data from minimally invasive high-resolution accelerometry suction cup tags deployed by the GEMM Lab to estimate the relative energetic cost of different foraging behaviors (see my first blog) of Pacific Coast Feeding Group (PCFG) gray whales that forage during summer months off the coast of Oregon. I learned a lot of new skills through this research project and am excited to share some of my odyssey with you.
To start, I want to highlight the technology that made this work possible: the high-resolution accelerometry suction cup tags. These suction cup tags not only record fine scale data about the whale’s movement and behavior from inertial sensors like accelerometers, magnetometers and gyroscopes, the tags also incorporate video and audio data to record what the whale is seeing and hearing in its environment. I used the data from these tags to achieve two research objectives relating to 1) describing foraging behavior of PCFG gray whales and 2) estimating the relative energetic cost of these behaviors.
Through the hard work of the GEMM Lab field team and collaborators John Calambokidis and Dr. Dave Cade, 10 of these high-resolution accelerometry tags were deployed to collect approximately 91.5 hours of data from PCFG gray whales. Excitingly, two of these tags were deployed on a known mother-daughter pair on the same day. The mother and her 8-year-old daughter were even observed foraging together while tagged later in the day and recorded each other while feeding (Figure 1)!
My first research objective was to quantitatively describe the foraging behaviors of PCFG gray whales. These quantitative descriptions exist for other baleen whale foraging behaviors, such as the lunge feeding behavior of rorqual whales (e.g., humpback, fin, and blue whales) where large mouthfuls of prey are engulfed, or the ram filtration feeding of bowhead and right whales where water is filtered for prey as the whale swims along with its mouth open. High-resolution accelerometry tag data has found that a strong acceleration signal is useful for detecting lunges (Goldbogen et al., 2013) and continuous fluking paired with low swim speeds signal the occurrence of ram filtration feeding (Simon et al., 2009). However, gray whales are the only baleen whales to use suction feeding behavior where the whale rolls to one side and sucks up water to filter for prey. Beyond describing side preferences when performing suction feeding (Woodward & Winn, 2006), this unique foraging behavior of gray whales lacks quantitative descriptions. My thesis works to add quantitative descriptions of gray whale suction feeding to the existing descriptions of baleen whale foraging behavior using high-resolution tag data. Based on previous drone focal follows that qualitatively describe gray whale foraging behaviors (Torres et al., 2018), I hypothesized that body position variables would be important for quantitatively describing PCFG gray whale foraging tactics using tag data. I anticipated that the signals of gray whale suction feeding behavior would be different from the signals of lunge and ram filtration feeding performed by other species of baleen whales (Figure 2).
Figure 1. Snapshots from the video footage of high-resolution accelerometry suction cup tags deployed on a mother-daughter pair of whales foraging together. The top photo is from the tag deployed on the daughter filming the mother, and the bottom photo is from the tag deployed on the mother filming the daughter.
Figure 2. Quantitative descriptions of foraging behaviors for different baleen whale feeding mechanisms. The left panel indicates the strong acceleration signal used to detect lunges of rorquals (e.g., humpback, fin, and blue whales). The acceleration figure is adapted from Izadi et al., (2022). The middle panel shows the continuous fluking and low swim speed signal for ram filtration feeding of bowheads (Simon et al., 2009). The right panel indicates the hypothesized importance of body position variables when quantitatively describing the unique suction feeding behavior of gray whales (my thesis).
My second objective was to estimate the relative energetic cost of PCFG gray whale foraging behaviors. Previous research has estimated the energetic cost of gray whale broad state behaviors (e.g., transit, search, and forage) using respiration rates (Sumich, 1986). However, respiration rates are difficult to use when estimating the energetic cost of fine scale behaviors, like the precise foraging tactics performed within a dive. Therefore, I calculated biologging-derived proxies of energy expenditure from the tag data such as stroke rate (i.e., the frequency of the whale’s fluke beats) and Overall Dynamic Body Acceleration (ODBA; i.e., the total body movement of the whale) to estimate the relative energetic cost of three different PCFG gray whale foraging tactics: benthic dig, headstand, and side swim. To put these energy expenditure proxies into a human example, if you were walking, your stroke rate would be the frequency of your steps and your ODBA would be all the acceleration of your body including your legs moving with each step, your swinging arms, and turning head. Stroke rate and ODBA are common proxies of energy expenditure that are easily calculated from biologging tag data and have been linked to metabolic rate in many species, including bottlenose dolphins (Allen et al., 2022) and fur seals (Jeanniard-Du-Dot et al., 2016).
When comparing these two proxies of energy expenditure between PCFG gray whale foraging tactics, I expected that the benthic dig foraging behavior would be the least energetically costly compared to the other PCFG foraging tactics. Benthic digs occur when the whale is rolled onto its side and plows through the sediment to suction up prey while leaving feeding pits on the seafloor. Benthic digs are assumed to be the primary foraging tactic of the majority of the gray whale population (Nerini, 1984) that feed in the Arctic and sub-Arctic region where gray whales must dive deeper to reach their prey in the bottom sediments, making it likely that a lower energetic cost of foraging motivates the higher use of this foraging tactic. Excitingly, preliminary results indicate that these three gray whale foraging tactics have different energy expenditures, which can potentially help explain patterns of behavior choice and tactic usage between different groups of gray whales. My thesis research is a foundational step toward better understanding gray whale foraging energetics by providing a means to assess prey requirements and inform management policies to reduce threats facing the PCFG gray whales. For example, previous work has shown that vessel disturbance reduces the searching time (Sullivan & Torres, 2018) and increases the stress levels (Lemos et al., 2022) of PCFG gray whales. My work builds on this by demonstrating an elevated energetic cost of some foraging tactics that could be used to support increasing the distance requirements between vessels and feeding PCFG whales as a way to reduce the energetic impact of vessel disturbance (Figure 3). Additionally, differences in energetic cost of foraging behaviors may help inform potential risks posed by changes in prey quality and quantity for gray whales using different foraging tactics.
Figure 3. Snapshot from the video footage of a high-resolution accelerometry suction cup tag deployed on a PCFG gray whale showing the high number of vessels present during a surfacing following a foraging dive. The energetic cost of foraging behaviors found in my thesis might suggest that increasing distance between vessels and forging whales could reduce the energetic impacts of vessel disturbance.
Overall, I am so grateful for my Master’s experience. I had the opportunity to work with amazing scientists that taught me many valuable skills and lessons that I will take with me as I move onto the next phase of my career. Throughout my degree I’ve had a lot of “firsts” and I am excited to embark on another as I prepare my first manuscripts for publication!
References:
Allen, A. S., Read, A. J., Shorter, K. A., Gabaldon, J., Blawas, A. M., Rocho-Levine, J., & Fahlman, A. (2022). Dynamic body acceleration as a proxy to predict the cost of locomotion in bottlenose dolphins. Journal of Experimental Biology, 225(4). https://doi.org/10.1242/jeb.243121
Goldbogen, J. A., Friedlaender, A. S., Calambokidis, J., McKenna, M. F., Simon, M., & Nowacek, D. P. (2013). Integrative approaches to the study of baleen whale diving behavior, feeding performance, and foraging ecology. BioScience, 63(2), 90–100. https://doi.org/10.1525/bio.2013.63.2.5
Izadi, S., Aguilar de Soto, N., Constantine, R., & Johnson, M. (2022). Feeding tactics of resident Bryde’s whales in New Zealand. Marine Mammal Science, 1–14. https://doi.org/10.1111/mms.12918
Jeanniard-Du-Dot, T., Trites, A. W., Arnould, J. P. Y., Speakman, J. R., & Guinet, C. (2016). Flipper strokes can predict energy expenditure and locomotion costs in free-ranging northern and Antarctic fur seals. Scientific Reports, 6. https://doi.org/10.1038/srep33912
Lemos, L. S., Haxel, J. H., Olsen, A., Burnett, J. D., Smith, A., Chandler, T. E., Nieukirk, S. L., Larson, S. E., Hunt, K. E., & Torres, L. G. (2022). Effects of vessel traffic and ocean noise on gray whale stress hormones. Scientific Reports, 12(1). https://doi.org/10.1038/s41598-022-14510-5
Nerini, M. (1984). A review of gray whale feeding ecology. In M. Lou Jones, S. L. Swartz, & S. Leatherwood (Eds.), The gray whale: Eschrichtius robustus (pp. 423–450). Academic Press. https://doi.org/10.1016/B978-0-08-092372-7.50024-8
Simon, M., Johnson, M., Tyack, P., & Madsen, P. T. (2009). Behaviour and kinematics of continuous ram filtration in bowhead whales (Balaena mysticetus). Proceedings of the Royal Society B: Biological Sciences, 276(1674), 3819–3828. https://doi.org/10.1098/rspb.2009.1135
Sullivan, F. A., & Torres, L. G. (2018). Assessment of vessel disturbance to gray whales to inform sustainable ecotourism. Journal of Wildlife Management, 82(5), 896–905. https://doi.org/10.1002/jwmg.21462
Sumich, J. L. (1986). Latitudinal distribution, calf growth and metabolism, and reproductive energetics of gray whales, Eschrichtius robustus. Oregon State University.
Torres, L. G., Nieukirk, S. L., Lemos, L., & Chandler, T. E. (2018). Drone up! Quantifying whale behavior from a new perspective improves observational capacity. Frontiers in Marine Science, 5. https://doi.org/10.3389/fmars.2018.00319
Woodward, B. L., & Winn, J. P. (2006). Apparent lateralized behavior in gray whales feeding off the Central British Columbia coast. Marine Mammal Science, 22(1), 64–73. https://doi.org/10.1111/j.1748-7692.2006.00006.x
The winds are consistently (and sometimes aggressively) blowing from the north here on the Oregon coast, which can only mean one thing – summer has arrived! Since mid-May, the GRANITE (Gray whale Response to Ambient Noise Informed by Technology and Ecology) team has been looking for good weather windows to survey for gray whales and we have managed to get five great field work days already. In today’s blog post, I am going to share what (and who) we have seen so far.
On our first day of the field season, PI Leigh Torres, postdoc KC Bierlich and myself, were joined by a special guest: Dr. Andy Read. Andy is the director of the Duke University Marine Lab, where he also runs his own lab, which focuses on conservation biology and ecology of marine vertebrates. Andy was visiting the Hatfield Marine Science Center as part of the Lavern Weber Visiting Scientist program and was hosted here by Leigh. For those of you that do not know, Andy was Leigh’s graduate school advisor at Duke where she completed her Master’s and doctoral degrees. It felt very special to have Andy on board our RHIB Ruby for the day and to introduce him to some friends of ours. The first whale we encountered that day was “Pacman”. While we are always excited to re-sight an individual that we know, this sighting was especially mind-blowing given the fact that Leigh had “just” seen Pacman approximately two months earlier in Guerrero Negro, one of the gray whale breeding lagoons in Mexico (read this blog about Leigh and Clara’s pilot project there). Aside from Pacman, we saw five other individuals, all of which we had seen during last year’s field season.
The first day of field work for the 2023 GRANITE field season! From left to right: Leigh Torres, Lisa Hildebrand, Andy Read, and KC Bierlich. Source: L. Torres.
Since that first day on the water, we have conducted field work on four additional days and so far, we have only encountered known individuals in our catalog. This fact is exciting because it highlights the strong site fidelity that Pacific Coast Feeding Group (PCFG) gray whales have to areas within their feeding range. In fact, I am examining the residency and space use of each individual whale we have observed in our GRANITE study for one of my PhD chapters to better understand the level of fidelity individuals have to the central Oregon coast. Furthermore, this site fidelity underpins the unique, replicate data set on individual gray whale health and ecology that the GRANITE project has been able to progressively build over the years. So far during this field season in 2023, we have seen 13 unique individuals, flown the drone over 10 of them and collected four fecal samples from two, which represent critical data points from early on in the feeding season.
Our sightings this year have not only highlighted the high site fidelity of whales to our study area but have also demonstrated the potential for internal recruitment of calves born to “PCFG mothers” into the PCFG. Recruitment to a population can occur in two ways: externally (individuals immigrate into a population from another population) or internally (calves born to females that are part of the population return to, or stay, within their mothers’ population). Three of the whales we have seen so far this year are documented calves from females that are known to consistently use the PCFG range, including our central Oregon coast study area. In fact, we documented one of these calves, “Lunita”, just last year with her mother (see Clara’s recap of the 2022 field season blog for more about Lunita). The average calf survival estimate between 1997-2017 for the PCFG was 0.55 (Calambokidis et al. 2019), though it varied annually and widely (range: 0.34-0.94). Considering that there have been years with calf survival estimates as low as ~30%, it is therefore all the more exciting when we re-sight a documented calf, alive and well!
“Lunita”, an example of successful internal recruitment
We have also been collecting data on the habitat and prey in our study system by deploying our paired GoPro/RBR sensor system. We use the GoPro to monitor the benthic substrate type and relative prey densities in areas where whales are feeding. The RBR sensor collects high-frequency, in-situ dissolved oxygen and temperature data, enabling us to relate environmental metrics to relative prey measurements. Furthermore, we also collect zooplankton samples with a net to assess prey community and quality. On our five field work days this year, we have predominantly collected mysid shrimp, including gravid (a.k.a. pregnant) individuals, however we have also caught some Dungeness and porcelain crab larvae. The GEMM Lab is also continuing our collaboration with Dr. Susanne Brander’s lab at OSU and her PhD student Lauren Kashiwabara, who plan on conducting microplastic lab experiments on wild-caught mysid shrimp. Their plan is to investigate the growth rates of mysid shrimp under different temperature, dissolved oxygen, and microplastic load conditions. However, before they can begin their experiments, they need to successfully culture the mysids in the lab, which is why we collect samples for them to use as their ‘starter culture’. Stay tuned to hear more about this project as it develops!
So, all in all, it has been an incredibly successful start to our field season, marked by the return of many familiar flukes and flanks! We are excited to continue collecting rock solid GRANITE data this summer to increase our efforts to understand gray whale ecology and physiology.
References
Calambokidis, J., Laake, J., and Perez, A. (2019). Updated analyses of abundance and population structure of seasonal gray whales in the Pacific Northwest, 1996-2017. IWC, SC/A17/GW/05 for the Workshop on the Status of North Pacific Gray Whales. La Jolla: IWC.