We need energy to function and survive. For animals in the wild who may have limited food availability, knowing how they spend their energy is a critical question for many scientists because it fundamentally informs how we understand their decisions about where they go and what they do. The entire field of foraging theory is founded on the concept that animals optimize their ratio of energy in and energy out so that they have enough energy to survive, reproduce (pass on their genes), watch out for threats, if need be, and rest. And, if we understand an animal’s ‘typical’ energy budget, we can then try to predict how disturbance or environmental change will affect their actual energy budgets as a consequence of that change. But how do we measure energy expenditure?
The most commonly measured energy currency is oxygen. Since our cells use oxygen to produce energy (this is why we need oxygen to live), we can measure oxygen consumption as a metric of energy expenditure. The more oxygen we consume, the more energy we’re expending. In ideal lab settings, oxygen consumption can be accurately measured by placing the subject in a chamber where the oxygen flow can be controlled (Speakman, 1999). However, you can probably see how that approach is problematic for measuring oxygen consumption in most large free-living animals, especially cetaceans. It isn’t exactly feasible to put a whale in a box.
Image 1. A great tit in a metabolic chamber. Figure 1 from Broggi et al., 2009
Fortunately, a tool called a spirometer was developed to measure oxygen consumption in restrained cetaceans. A spirometer is a device that can be placed over the blowhole(s) of an individual to accurately measure the amount of air that is exhaled and inhaled (Figure 1). For trained cetaceans in captivity (e.g., dolphins), spirometers can be used to quantify how respiration changes after the animal performs certain behaviors (Fahlman et al., 2019). The breathing patterns of diving mammals are particularly interesting because they cannot breathe during most of their exercise (energy expenditure) as they are underwater. So, their breathing patterns after a dive tell us a lot about how much energy they spent during that dive. For example, Fahlman et al. (2019) used spirometer data from dolphins in captivity to study how their breathing patterns changed while recovering from dives of different durations. Interestingly, they found that after longer dives, dolphins took larger breaths (i.e., inhaled more air) while recovering but did not change the number of breaths. This finding is particularly relevant to the work we are conducting in the GEMM lab, where we utilize breathing patterns to quantify the energy expenditure of cetaceans in the wild, where spirometers cannot be used.
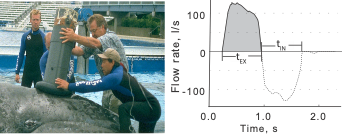
Figure 1. Figure 1 from Sumich et al. (2023). Left: a spirometer being held over the blow holes of JJ, a gray whale calf at sea world in 1997; one of the rare times that a large baleen whale was in captivity and available for these measurements. Right: example of a plot created using the data from a spirometer over JJ’s blow holes. The duration of a “blow” (exhale followed by immediate inhale) is on the x-axis, the flow rate (in liters per second) is on the y-axis. The positive curve during the exhale shows that the whale strongly exhales a lot of air very quickly, then the negative curve shows the whale inhaling a lot of air very quickly.
In a previous blog, I described how inter-breath intervals (the time between consecutive blows) are useful for estimating energy expenditure in free-living cetaceans. Essentially, a shorter interval indicates that the whale was just engaged in an energetically demanding activity. When you’re recovering from a sprint, you breathe faster (i.e., with shorter inter-breath intervals), than when you’re recovering from a walk. However, a big assumption in using inter-breath intervals as a proxy for energy expenditure is that every breath is equal. But as Fahlman et al. emphasize in their 2016 paper, every blow is not equal (Fahlman et al., 2016). In addition to varying the time between breaths, an animal can vary the intensity of each breath (e.g., Fahlman et al., 2019), the duration of each breath (Sumich et al., 2023), the number of breaths, and even the expansion of their nostrils (Nazario et al., 2022; check out this blog for more).
Altogether, this means that it’s important to measure every breath and that no one metric tells the complete story. This also means my research question focused on comparing the energetic costs of different tactics is more complicated than I originally thought. If we go back to the first blog I wrote on this topic, I was planning ons only using inter-breath intervals to estimate energy expenditure. Fast forward four years, with all my new knowledge gained on respiration variability, I’ve modified my plan and now I’m working to first understand how all these different metrics of breathing relate to each other. Then, I’ll compare how breathing varies between different foraging tactics, which is an important follow up to my questions around individual specialization of foraging tactics. If different whales are using different foraging behaviors, does that mean they’re spending different amounts of energy? If so, are certain behaviors more advantageous than others? Of course, these answers are incomplete without understanding the prey the whales are eating, but that’s something that PhD student Nat Chazal is working to understand (check out her recent blog here). For now, I’m working on bringing integrating all the measures of breathing, then we will start putting the story together and finding some answers to our pressing questions.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly alert when we make a new post! Just add your name into the subscribe box below!
References
Broggi, J., Hohtola, E., Koivula, K., Orell, M., & Nilsson, J. (2009). Long‐term repeatability of winter basal metabolic rate and mass in a wild passerine. Functional Ecology, 23(4), 768–773. https://doi.org/10.1111/j.1365-2435.2009.01561.x
Fahlman, A., Brodsky, M., Miedler, S., Dennison, S., Ivančić, M., Levine, G., Rocho-Levine, J., Manley, M., Rocabert, J., & Borque-Espinosa, A. (2019). Ventilation and gas exchange before and after voluntary static surface breath-holds in clinically healthy bottlenose dolphins, Tursiops truncatus. Journal of Experimental Biology, 222(5), jeb192211. https://doi.org/10.1242/jeb.192211
Fahlman, A., van der Hoop, J., Moore, M. J., Levine, G., Rocho-Levine, J., & Brodsky, M. (2016). Estimating energetics in cetaceans from respiratory frequency: Why we need to understand physiology. Biology Open,5(4), 436–442. https://doi.org/10.1242/bio.017251
Nazario, E. C., Cade, D. E., Bierlich, K. C., Czapanskiy, M. F., Goldbogen, J. A., Kahane-Rapport, S. R., Hoop, J. M. van der, Luis, M. T. S., & Friedlaender, A. S. (2022). Baleen whale inhalation variability revealed using animal-borne video tags. PeerJ, 10, e13724. https://doi.org/10.7717/peerj.13724
Speakman, J. R. (1999). The Cost of Living: Field Metabolic Rates of Small Mammals. In A. H. Fitter & D. G. Raffaelli (Eds.), Advances in Ecological Research (Vol. 30, pp. 177–297). Academic Press. https://doi.org/10.1016/S0065-2504(08)60019-7
Sumich, J. L., Albertson, R., Torres, L. G., Bird, C. N., Bierlich, K. C., & Harris, C. (2023). Using audio and UAS-based video for estimating tidal lung volumes of resting and active adult gray whales (Eschrichtius robustus). Marine Mammal Science, 1(8). https://doi.org/10.1111/mms.13081
By Alejandro Fernandez Ajo, PhD student at the Department of Biology, Northern Arizona University, Visiting scientist in the GEMM Lab working on the gray whale physiology and ecology project
Two years ago, in August 2018, I came to Newport and visited the Hatfield Marine Science Center for the first time with an NSF/RCN-founded laboratory exchange with the GEMM Lab and met Dr. Leigh Torres. My goals during this exchange where to learn about non-invasive fieldwork techniques for studying free-range whales while interacting, exchanging ideas, and networking with the GEMM Lab members; also, to discuss some projects and thoughts for future collaborations with Dr. Torres. During those two weeks in Newport, I had the opportunity to help with field work on the project “Evaluation of gray whale ecology and physiology in response to variable ambient ocean noise conditions”, which aims to evaluate the hormonal variability and health of the gray whales that forage along the Oregon coast in the context of multiple stressors. I would return during the summers of 2019 and 2020 as a visiting scientist and research assistant to work on this project. This year the experience has been a bit different in terms of interactions with the HMSC community due to COVID-19; however, we were able to successfully start the field season in time and now we are wrapping up our second month of surveys with many new and interesting data gathered, and many new, unforgettable memories to be treasured. Working with these animals is incredibly fascinating because there are so many things we don´t know about them, and the questions can become both overwhelming and exciting.
An essential part of this project, and arguably any research project done with cetaceans, is the identification of individuals. Hence, considerable effort is expended each year attempting to photograph every gray whale possible within our study region and to identify each whale we encounter. The GEMM Lab maintains a catalog of the gray whales that visit the Oregon coast, a sub-population known as the Pacific Coast Feeding Group (PCFG). This catalog currently consists of 173 individuals. which we frequently compare with a larger catalog of gray whales that includes 2060 individuals observed since 1977 (Cascadia Research Collective). These methods allow us to know who is who among the whales we encounter each day at sea.
The different species of cetaceans can be individually identified by markings on their bodies, very much like fingerprints in humans. Some features on these animals are unique and conserved through life. For example, Southern and Northern right whales are identified by the callosity patters in their heads (Picture 1), while humpback whales are mostly identified individually by the shape and the patterns of black and white pigmentation on the underside of their fluke (Picture 2). Gray whales have very mottled skin coloration, so we use a combination of markings and features to identify individuals: pigmentation patterns, scars, shape and pigmentation of their fluke, and sometimes the shape of their knuckles, which are a series of “humps” that gray whales have instead of a dorsal fin on their back. It might sound very difficult to do, and it can be a tedious task, however as you train your eye it becomes easier, and features that at first seemed undistinguishable become recognizable and unique (Picture 3). As a reward, it is such a joy to find a match and recognize old friends when they arrive from their long journeys in the vast ocean each year to the Oregon coast.
The callosity pattern of a southern right whale.Photo by Alejandro Fernández Ajó – Instituto de Conservación de Ballenas.Three different humpback whales with notably different pigmentation patterns on their flukes. Photographs captured under NOAA/NMFS permit #21678.The gray whale “Knife” we have observed frequently this summer off the Oregon coast. Can you see how the right pigmentation pattern identified looks like a knife? Photograph captured under NOAA/NMFS permit #21678.
As a result of our photo-identification efforts and the high site-fidelity of the whales we study, the large majority of the gray whales we observe here in Oregon are known individually. For many whales, we also have detailed sightings records that can span years and decades, that document calving history, lactation, appearance of scars indicative of injury or entanglement, minimum age, sex, habitat-use patterns, behaviors, etc. Holding such detailed information of individual whales provides incredible contributions to our understanding of the basic patterns in life history of whales, such as reproduction rates, calving intervals, age of first reproduction, etc. Moreover, when these life history events are linked with physiological sample collection large steps can be made in the development and validation of physiological methods. Many endocrine assays currently in use for whales are based on non-traditional sample types including feces, respiratory vapor, and baleen, which have been validated using the catalogs of well-known individuals to verify that measured hormones reflect patterns expected for various physiological states. For example, we can compare endocrine data of confirmed pregnant females, known mature males, and known-injured whales to learn how whale physiological responses are different during different life history events (e.g., Burgess et al. 2017, 2018, Corkeron et al. 2017, Hunt et al. 2006, 2016, 2018, Lysiak et al. 2018, Rolland et al. 2005).
Here in Oregon we are learning from the lives of the gray whales we study, and here I want to share with you two of their stories, one happy and one not-so-happy.
Let´s start with the not-so-happy story so we can end with some brighter news. On June 24 this year, we encountered a whale near Cape Foulweather, which is a very tricky area to work as there are many rocks and shallow water that make the sea conditions very choppy even with low swell. We started documenting the sighting as usual, taking pictures of the left side, the right side and ideally also the fluke of the whale. As we approached this whale, we started noting that something was wrong with its fluke. With the challenging sea conditions, it was not easy to approach the whale and the whale was not exposing its fluke when diving. When we put our drone up to collect photogrammetry and behavior data we gained a much better perspective. This whale has a bad injury on it fluke (Picture 4.C). On the boat we started making conjectures about the cause of this terrible injury that had basically amputate most of its left fluke lobe. Once back on shore, we sorted out the photos and compared the field images captured during the day with the photo-ID catalog and we made a match. This whale is known in our catalog as “ROLLER SKATE”, is a female, and was first sighted in 2015, so she is at least 5 years old today.
The story unfolded when we reviewed Roller Skate’s sighting history. Interestingly we observed this same whale in the same location last September 2019. Unfortunately, it was a very brief encounter but enough for photo documentation of the whale and an interesting observation. Here I quote the field notes that Dr. Torres wrote from this sighting: “September 6th, 2019. Sighting 9: Scattered whales feeding and/or traveling across area to north of Cape Foulweather. One whale had recently chopped fluke; tried to re-find to get better photos but could not (looking at photos now, this whale is clearly entangled in line!). Ceiling too low for UAS [drone flight].” (Picture 4.B).
Progressive photos of Roller Skate’s fluke. In 2017, her fluke was healthy (A). In 2019, we observed her fluke with a line entanglement and “chewed up” (B). In 2020, we have observed the resulting wound and healing process (C). Photographs captured under NOAA/NMFS permit #21678.
Roller skate’s story is an example of how essential is to keep an ID catalog. After a close-up examination of the 2019 picture, we can clearly see a rope entanglement (Picture 4.B). Photos from previous years show how beautiful and healthy her fluke was before this event (Picture 4.A). This event is heart breaking to witness, but this whale could be considered lucky because she was able to shed the gear and survive this entanglement, at least in the short term. Additionally, we can learn from Roller Skate’s misfortune to help us understand what the consequences of such an injury (stressor) may be on the physiology of a whale. We have been eager to collect a fecal sample from Roller Skate to analyze how her hormone levels compare to non-injured whales. Fortunately, we got lucky a couple weeks ago and collected this sample, so now we need to get in the lab and analyze the samples. But more questions remain: Will this injury impact her ability to reproduce? If so, for how long? And at a larger scale, what are the population consequences of such events? If we can understand the magnitude of lethal and sublethal human caused impacts on individual whales and their populations from events such as entanglements, we can develop better methods to mitigate and limit such hazards for whales in their environments.
As I promised, there is also some good news to share. A very well-known PCFG whale, almost a celebrity I dear to say, is “Scarback”, or as we like to call her “Scarlett”. Scarlett is a female known since 1996, making her at least 24 years old, and she also has a very bad injury of unknown origin. Scarlett has a terrible scar on her back that is theorized to have been caused by an explosive harpoon, or maybe a bad ship-strike (Picture 5), but we really do not know. However, we do know she survived this injury and this year she brought a new calf into the population (Picture 6). This is the second calf we have documented from Scarlett, with her previous calf sighted during the 2016 field season and we call it “Brown”. Scarlett is an example of how resilient these amazing giants can be; however, it is likely that while she was recovering from this injury, she was unable to reproduce. How many calves from Scarlett did the PCFG population “lose” due to such a tragedy? We can´t know, but we are learning, and her story will also help us understand whale physiology as we will analyze her fecal hormones and body condition during pregnancy, lactation, and resting phases.
The scar on Scarlett’s back. This significant wound has an unknown origin.Photograph captured under NOAA/NMFS permit #21678.
Scarlett is a survivor. We need to recognize that we are sharing the ocean with different forms of life. We need to acknowledge their existence and understand how our use of the oceans is affecting them, and, more importantly, work toward improving their conditions. I hope that with our research we highlight and communicate how amazing are these animals, and how important are they for marine ecosystems. And ultimately, I hope our work helps minimize the impacts that affect other forms of ocean life that coexist with us, both above and below the surface.
The well-known gray whale “Scarlett” surfaces under her calf while it checks out the GEMM Lab field team. Photo by Alejandro Fernandez Ajo taken under NOAA/NMFS permit #21678.
References:
Burgess, E., Hunt, K. E., Kraus, S. D. and Rolland, R. M. (2016). Get the most out of blow hormones: validation of sampling materials, field storage and extraction techniques for whale respiratory vapor samples. Conservation Physiology, 4, cow024.
Burgess, E. A., Hunt, K. E., Kraus, S. D. and Rolland, R. M. (2018). Quantifying hormones in exhaled breath for physiological assessment of large whales at sea. Scientific Reports, 8, 10031.
Corkeron, P. J., Rolland, R. M., Hunt, K. E. and Kraus, S. D. (2017). A right whale PooTree: Fecal hormones and classification trees identify reproductive states in North Atlantic right whales (Eubalaena glacialis). Conservation Physiology, 5, cox006. DOI: 10.1093/conphys/cox006.
Hunt, K., Lysiak, N., Moore, M. and Rolland, R. (2017). Multi-year longitudinal profiles of cortisol and corticosterone recovered from baleen of North Atlantic right whales (Eubalaena glacialis). General and Comparative Endocrinology, 254, 50-59. DOI: 10.1016/j.ygcen.2017.09.009.
Hunt, K., Lysiak, N. S. J., Matthews, C. J. D., et al. (2018). Multi-year patterns in testosterone, cortisol and corticosterone in baleen from adult males of three whale species. Conservation Physiology, 6, coy049. DOI: 10.1093/conphys/coy049.
Hunt, K. E., Rolland, R. M., Kraus, S. D. and Wasser, S. K. (2006). Analysis of fecal glucocorticoids in the North Atlantic Right Whale (Eubalaena glacialis). General and Comparative Endocrinology, 148, 260-272.
Lysiak, N., Trumble, S., Knowlton, A. and Moore, M. (2018). Characterizing the duration and severity of fishing gear entanglement on a North Atlantic right whale (Eubalaena glacialis) using stable isotopes, steroid and thyroid hormones in baleen. Frontiers in Marine Science. DOI: 10.3389/fmars.2018.00168.
Rolland, R. M., Hunt, K. E., Kraus, S. D. and Wasser, S. K. (2005). Assessing reproductive status of right whales (Eubalaena glacialis) using fecal hormone metabolites. General and Comparative Endocrinology, 142, 308-317.
By: Alexa Kownacki, Ph.D. Student, OSU Department of Fisheries and Wildlife, Geospatial Ecology of Marine Megafauna Lab
While sitting on my porch and watching the bees pollinate the blooming spring flowers, I intermittently pause to scratch the hives along my shoulders and chest. In the middle of my many Zoom calls, I mute myself and stop my video because a wave of pollen hits my face and I immediately have to sneeze. With this, I’m reminded: Welcome to prime allergy season in the Northern Hemisphere. As I was scratching my chronic idiopathic urticaria (hives caused by an overactive immune system), I asked myself “Do dolphins get hives?” I had no idea. I know most terrestrial mammals can and do—just yesterday, one of the horses in the nearby pasture was suffering from a flare of hives. But, what about aquatic and marine mammals?
Springtime flowers blooming on the Central California Coast 2017. (Image Source: A. Kownacki)
As with most research on marine mammal health, knowledge is scare and is frequently limited to studies conducted on captive and stranded animals. Additionally, most of the current theories on allergic reactions in marine mammals are based on studies from terrestrial wildlife and humans. Because nearly all research on histamine pathways centers on terrestrial animals, I wanted to see what information exists the presence of skin allergies in marine mammals.
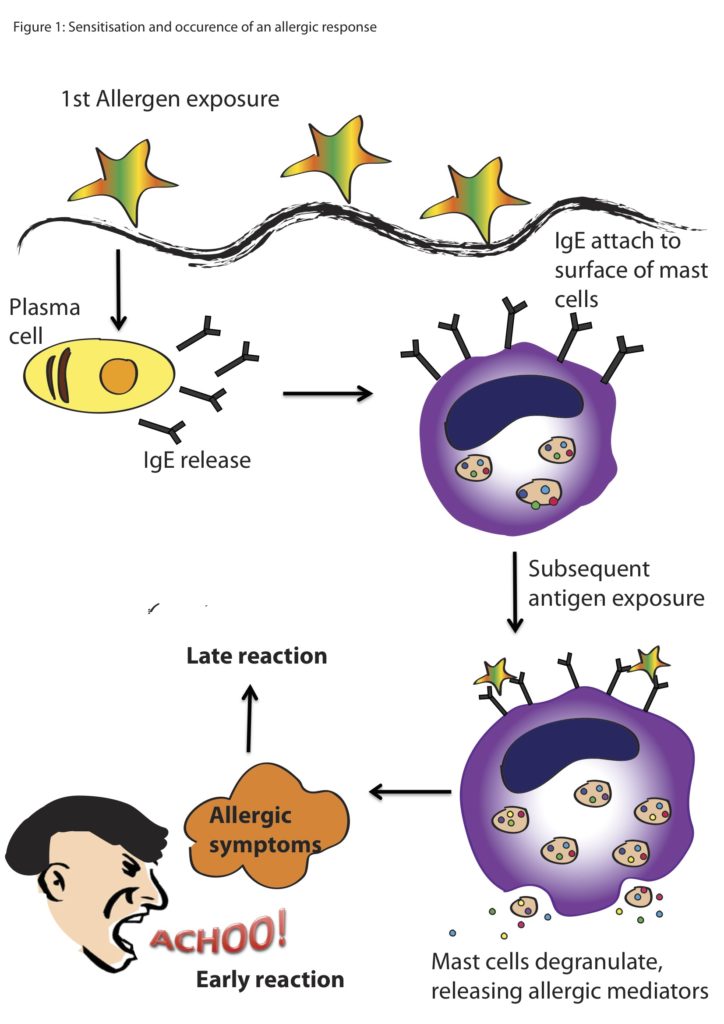
Allergic reactions trigger a cascade within the body, beginning with the introduction of a foreign body, which for many people is pollen. The allergen binds to antibodies that are produced to fight potentially harmful substances. Once this allergen binds to different types of cells, including mast cells, chemicals like histamines are released. Histamines cause the production of mucus and constriction of blood vessels, and thus are the reason your eyes water, your nose runs, or you start coughing.
Basic cartoon of an allergic reaction from exposure to the allergen to the reaction from the animal. (Image Source: Scientific Malaysian)
As you probably can tell just by looking at a marine mammal, they have thicker skin and fewer mucus membranes that humans, due to the fact that they live in the water. However, mast cells or mast cell-like cells have been described in most vertebrate lineages including mammals, birds, reptiles, amphibians, and bony fishes (Hellman et al. 2017, Reite and Evenson 2006). Mast cell-like cells have also been described in an early ancestor of the vertebrates, the tunicate, or sea squirt (Wong et al. 2014). Therefore, allergic-reaction cascades that may present as hives, red and itchy eyes or nose in humans, also exist in marine mammals, but perhaps cause different or less visible symptoms.
Skin conditions in cetaceans are gathering interest within the marine mammal health community. Even our very own Dawn Barlow, Dr. Leigh Torres, and Acacia Pepperassessed the skin conditions in New Zealand blue whales in their recent publication. Most visible skin lesions or markings on cetaceans are caused by parasites, shark bits, fungal infections, and fishery or boat interactions (Leone et al. 2019, Sweeney and Ridgway 1985). However, there is very little scientific literature about allergic reactions in marine mammals, let alone cetaceans. That being said, I managed to find a few critical pieces of information supporting the theory that marine mammals do in fact have allergies that can produce dermal reactions similar to hives in humans.
In one study, three captive bottlenose dolphins developed reddened skin, sloughing, macules, and wheals on their ventral surfaces (Monreal-Pawlowsky et al. 2017). The medical staff first noticed this atopic dermatitis in 2005 and observed the process escalate over the next decade. Small biopsy samples from the affected areas on the three dolphins coincided with the appearance of four pollens in the air within the geographic region: Betula, Pistacia, Celtis, and Fagus (Monreal-Pawlowsky et al. 2017). Topical prednisone treatments were applied to the affected areas at various dosages that slowly resolved the skin irritations. Researchers manufactured an allergy vaccine using a combination of the four pollens in hopes that it would prevent further seasonal outbreaks, but it was unsuccessful. In the coming years, the facility intends to adjust the dosages to create a successful vaccine.
In the three top images, visible skin irritation including redness, macules, wheals, and sloughing are present. In the image below, the above animal was treated with methylprednisolone and the skin irritation subsides. (Monreal-Pawlowsky et al. 2017)
In addition to the above study, there is an unpublished case of suspected allergic reaction to another pollen that produces a pruritic reaction on the ventral areas of dolphins on a seasonal basis (Vicente Arribes, personal communication). Although there are only a few documented cases of environmentally-triggered allergic reactions that are visible on the dermal layer of cetaceans, I believe this evidence makes the case that some cetaceans suffer from allergies much like us. So, next time you’re enjoying the beautiful blooms and annoyingly scratch your eyes, know that you are not alone.
Image Source: FurEver Family
Citations:
Barlow DR, Pepper AL and Torres LG (2019) Skin Deep: An Assessment of New Zealand Blue Whale Skin Condition. Front. Mar. Sci. 6:757.doi: 10.3389/fmars.2019.00757
Hellman LT, Akula S, Thorpe M and Fu Z (2017) Tracing the Origins of IgE, Mast Cells, and Allergies by Studies of Wild Animals. Front. Immunol. 8:1749. doi: 10.3389/fimmu.2017.01749
Leone AB, Bonanno Ferraro G, Boitani L, Blasi MF. Skin marks in bottlenose dolphins (Tursiops truncatus) interacting with artisanal fishery in the central Mediterranean Sea. PLoS One. 2019;14(2):e0211767. Published 2019 Feb 5. doi:10.1371/journal.pone.0211767
Monreal-Pawlowsky T, Fernández-Bellon H, Puigdemont A (2017) Suspected Allergic Reaction in Bottlenose Dolphins (Tursiops truncatus). J Vet Sci Ani Husb 5(1): 108. doi: 10.15744/2348-9790.5.108
Reite OB, Evensen O. Inflammatory cells of teleostean fish: a review focusing on mast cells/eosinophilic granule cells and rodlet cells. Fish Shellfish Immunol (2006) 20:192–208. doi:10.1016/j.fsi.2005.01.012
Sweeney, J. C., & Ridgway, S. H. (1975). Common diseases of small cetaceans. J. Am. Vet. Med. Assoc, 167(7), 533-540.
Wong GW, Zhuo L, Kimata K, Lam BK, Satoh N, Stevens RL. Ancient originof mast cells. Biochem Biophys Res Commun (2014) 451:314–8. doi:10.1016/j.bbrc.2014.07.124
By: Alexa Kownacki, Ph.D. Student, OSU Department of Fisheries and Wildlife, Geospatial Ecology of Marine Megafauna Lab
As technology has developed over the past ten years, toxins
in marine mammals have become an emerging issue. Environmental toxins are
anything that can pose a risk to the health of plants or animals at a dosage.
They can be natural or synthetic with varying levels of toxicity based on the
organism and its physiology. Most prior research on the impacts toxins before
the 2000s was conducted on land or in streams because of human proximity to
these environments. However. with advancements in sampling methods, increasing
precision in laboratory testing, and additional focus from researchers, marine
mammals are being assessed for toxin loads more regularly.
A dolphin swims through a diesel slick caused by a small oil spill in a port. (Image Source: The Ocean Update Blog)
Marine mammals live most of their lives in the ocean or other aquatic systems, which requires additional insulation for protection from both cold temperatures and water exposure. This added insulation can take the form of lipid rich blubber, or fur and hair. Many organic toxins are lipid soluble and therefore are more readily found and stored in fatty tissues. When an organic toxin like a polychlorinated biphenyl (PCB) is released into the environment from an old electrical transformer, it persists in sediments. As these sediments travel down rivers and into the ocean, these toxic substances slowly degrade in the environment and are lipophilic (attracted to fat). Small marine critters eat the sediment with small quantities of toxins, then larger critters eat those small critters and ingest larger quantities of toxins. This process is called biomagnification. By the time a dolphin consumes large contaminated fishes, the chemical levels may have reached a toxic level.
The process by which PCBs accumulate in marine mammals from small particles up to high concentrations in lipid layers. (Image Source: World Ocean Review)
Marine mammal scientists are teaming with biochemists and ecotoxicologists to better understand which toxins are more lethal and have more severe long-term effects on marine mammals, such as decreased reproduction rates, lowered immune systems, and neurocognitive delays. Studies have already shown that higher contaminant loads in dolphins cause all three of these negative effects (Trego et al. 2019). As a component of my thesis work on bottlenose dolphins I will be measuring contaminant levels of different toxins in blubber. Unfortunately, this research is costly and time-consuming. Many studies regarding the effects of toxins on marine mammals are funded through the US government, and this is where the public can have a voice in scientific research.
Rachel Carson examines a specimen from a stream collection site in the 1950s. (Image Source: Alfred Eisenstaedt/ The LIFE picture collection/ Getty Images.)
Prior to the 1960s, there were no laws regarding the discharge of toxic substances into our environment. When Rachel Carson published “Silent Spring” and catalogued the effects of pesticides on birds, the American public began to understand the importance of environmental regulation. Once World War II was over and people did not worry about imminent death due to wartime activities, a large portion of American society focused on what they were seeing in their towns: discharges from chemical plants, effluents from paper mills, taconite mines in the Great Lakes, and many more.
Discharge from a metallic sulfide mine collects in streams in northern Wisconsin. (Image Source: Sierra Club)
However, it was a very different book regarding pollutants in the environment that caught my attention – and that of a different generation and part of society – even more than “Silent Spring”. A book called “The Lorax”. In this 1972 children’s illustrated book by Dr. Seuss, a character called the Lorax “speaks for the trees”. The Lorax touches upon critical environmental issues such as water pollution, air pollution, terrestrial contamination, habitat loss, and ends with the poignant message, “Unless someone like you cared a whole awful lot, nothing is going to get better. It’s not.”
The original book cover for “The Lorax” by Dr. Seuss. (Image source: Amazon.com)
Within a decade, the US Environmental Protection Agency (EPA) was formed and multiple acts of congress were put in place, such as the National Environmental Policy Act, Clean Air Act, Clean Water Act, and Toxic Substances Control Act, with a mission to “protect human health and the environment.” The public had successfully prioritized protecting the environment and the government responded. Before this, rivers would catch fire from oil slicks, children would be banned from entering the water in fear of death, and fish would die by the thousands. The resulting legislation cleaned up our air, rivers, and lakes so that people could swim, fish, and live without fear of toxic substance exposures.
The Cuyahoga River on fire in June 1969 after oil slicked debris ignited. (Image Source: Ohio Central History).
Fast forward to 2018 and times have changed yet again due to fear. According to a Pew Research poll, terrorism is the number one issue that US citizens prioritize, and Congress and the President should address. The environment was listed as the seventh highest priority, below Medicare (“Majorities Favor Increased Spending for Education, Veterans, Infrastructure, Other Govt. Programs.”). With this societal shift in priorities, research on toxins in marine mammals may no longer grace the covers of the National Geographic, Science, or Nature, not for lack of importance, but because of the allocation of taxpayer funds and political agendas. Meanwhile, long-lived marine mammals will still be accumulating toxins in their blubber layers and we, the people, will need to care a whole lot, to save the animals, the plants, and ultimately, our planet.
The Lorax telling the reader how to save the planet. (Image Source: “The Lorax” by Dr. Seuss via the Plastic Bank)
Citations:
“Majorities Favor Increased Spending for Education,
Veterans, Infrastructure, Other Govt. Programs.” Pew Research Center for the
People and the Press, Pew Research Center, 11 Apr. 2019,
www.people-press.org/2019/04/11/little-public-support-for-reductions-in-federal-spending/pp_2019-04-11_federal-spending_0-01-2/.
Marisa L. Trego, Eunha Hoh, Andrew Whitehead, Nicholas M. Kellar, Morgane Lauf, Dana O. Datuin, and Rebecca L. Lewison. Environmental Science & Technology201953 (7), 3811-3822. DOI: 10.1021/acs.est.8b06487
By Alejandro Fernánez Ajó, PhD student at NAU and GEMM Lab research technician
Although
commercial whaling is currently banned and several whale populations show
evidence of recovery, today´s whales are exposed to a variety of other human
stressors (e.g., entanglement in fishing gear, vessel strikes, shipping noise,
climate change, etc.; reviewed in Hunt et al., 2017a). The recovery and
conservation of large whale populations is particularly important to the
oceanic environment due to their key ecological role and unique biological
traits, including their large body size, long lifespan and sizable home ranges
(Magera et al., 2013; Atkinson et al., 2015; Thomas and Reeves, 2015). Thus,
scientists must develop novel tools to overcome the challenges of studying
whale physiology in order to distinguish the relative importance of the different
impacts and guide conservation actions accordingly (Ayres et al., 2012; Hunt et
al., 2013).
To this end,
baleen hormone analysis represents a powerful tool for retrospective assessment
of patterns in whale physiology (Hunt et al., 2014, 2016, 2017a, 2017b, 2018;
Lysiak et. al., 2018; Fernández Ajó et al., 2018; Rolland et al., 2019).
Moreover, hormonal panels, which include multiple hormones, are helping to
better clarify and distinguish between the physiological effects of different
sources of anthropogenic and environmental stressors (Ayres et al., 2012;
Wasser et al., 2017; Lysiak et al., 2018; Romero et al., 2015).
What is Baleen? Baleen is a stratified epithelial tissue consisting of long, fringed plates that grow downward from the upper jaw, which collectively form the whale´s filter-feeding apparatus (Figure 1). This tissue accumulates hormones as it grows. Hormones are deposited in a linear fashion with time so that a single plate of baleen allows retrospective assessment and evaluation of a whales’ physiological condition, and in calves baleen provides a record of the entire lifespan including part of their gestation. Baleen samples are also readily accessible and routinely collected during necropsy along with other samples and relevant information.
Figure 1: Top: A baleen plate from a southern right whale calf (Source: Fernández Ajó et al. 2018). Bottom: A southern right whale with mouth open exposing its baleen (photo credit: Stephen Johnson).
Why are the
Southern Right Whales calves (SRW) dying in Patagonia?
I am a Fulbright Ph.D. student in the Buck Laboratory at Northern Arizona University since Fall 2017, a researcher with the Whale Conservation Institute of Argentina (Instituto de Conservación de Ballenas) and Field Technician for the GEMM Lab over the summer. I focus my research on the application and development of novel methods in conservation physiology to improve our understanding of how physiological parameters are affected by human pressures that impact large whales and marine mammals. I am especially interested in understanding the underlaying causes of large whale mortalities with the aim of preventing their occurrence when possible. In particular, for my Ph.D. dissertation, I am studying a die-off case of Southern Right Whale (SRW) calves, Eubalaena australis, off Peninsula Valdés (PV) in Patagonia-Argentina (Figure 2).
Prior to
2000, annual calf mortality at PV was considered normal and tracked the
population growth rate (Rowntree et al., 2013). However, between 2007 and 2013,
558 whales died, including 513 newborn calves (Sironi et al., 2018). Average
total whale deaths per year increased tenfold, from 8.2 in 1993-2002 to 80 in
2007-2013. These mortality levels have never before been observed for the
species or any other population of whales (Thomas et al., 2013, Sironi et al.,
2018).
Figure 2: Study area, the red dots along the shoreline indicate the location where the whales were found stranded at Península Valdés in 2018 (Source: The Right Whale Program Research Report 2018, Sironi and Rowntree, 2018)
Among several hypotheses proposed to explain these elevated calf mortalities, harassment by Kelp Gulls, Larus dominicanus, on young calves stands out as a plausible cause and is a unique problem only seen at the PV calving ground. Kelp gull parasitism on SRWs near PV was first observed in the 1970’s (Thomas, 1988). Gulls primarily harass mother-calf pairs, and this parasitic behavior includes pecking on the backs of the whales and creating open wounds to feed on their skin and blubber. The current intensity of gull harassment has been identified as a significant environmental stressor to whales and potential contributor to calf deaths (Marón et al., 2015b; Fernández Ajó et al., 2018).
Figure 3: The significant preference for calves as a target of gull attacks highlights the impact of this parasitic behavior on this age class. The situation continues to be worrisome and serious for the health and well-being of newborn calves at Península Valdés. Left: A Kelp Gull landing on whale´s back to feed on her skin and blubber (Photo credit: Lisandro Crespo). Right: A calf with multiple lesions on its back produced by repeated gull attacks (Photo credit: ICB).
Quantifying gull inflicted wounds
Photographs of gull wounds on whales taken during necropsies and were quantified and assigned to one of seven objectively defined size categories (Fig. 4): extra-small (XS), small (S), medium (M), large (L), extra-large (XL), double XL (XXL) and triple XL (XXXL). The size and number of lesions on each whale were compared to baleen hormones to determine the effect of the of the attacks on the whales health.
Impact
factors such as injuries, predation avoidance, storms, and starvation promote
an increase in the secretion of the glucocorticoids (GCs) cortisol and
corticosterone (stress hormones), which then induce a variety of physiological
and behavioral responses that help animals cope with the stressor. Prolonged exposure
to chronic stress, however, may exceed the animal’s ability to cope with such
stimuli and, therefore, adversely affects its body condition, its health, and
even its survival. Triiodothyronine (T3), is the most biologically active form
of the thyroid hormones and helps regulate metabolism. Sustained food
deprivation causes a decrease in T3 concentrations, slowing metabolism to
conserve energy stores. Combining GCs and T3 hormone measures allowed us to
investigate and distinguish the relative impacts of nutritional and other
sources of stressors.
Combining these novel methods produced unique results about whale physiology. With my research, we are finding that the GCs concentrations measured in calves´ baleen positively correlate with the intensity of gull wounding (Figure 4, 1 and 2), while calf’s baleen thyroid hormone concentrations are relative stable across time and do not correlate with intensity of gull wounding (Figure 4 – 3). Taken together these findings indicate that SRW calves exposed to Kelp gull parasitism and harassment experience high levels of physiological stress that compromise their health and survival. Ultimately these results will inform government officials and managers to direct conservation actions aimed to reduce the negative interaction between Kelp gulls and Southern Right Whales in Patagonia.
Figure 4: Physiological stress correlates with number of gull lesions (1 and 2). According to the best-fit linear model, immunoreactive baleen corticosterone (B) and cortisol (F) concentrations increased with wound severity (i.e. number of gull lesions). However, nutritional status indexed by baleen immunoreactive triiodothyronine (T3) concentrations does not correlate with the number of gull lesions (3). (Fernández Ajó et al. 2019, manuscript under revision)
Baleen hormones represent a powerful tool for
retrospective assessments of longitudinal trends in whale physiology by helping
discriminate the underlying mechanisms by which different stressors may affect
a whale’s health and physiology. Moreover, while most sample types used for
studying whale physiology provide single time-point measures of current
circulating hormone levels (e.g., skin or respiratory vapor), or information
from previous few hours or days (e.g., urine and feces), baleen tissue provides
a unique opportunity for longitudinal analyses of hormone patterns. These
retrospective analyses can be conducted for both stranded or archived
specimens, and can be conducted jointly with other biological markers (e.g.,
stable isotopes and biotoxins) to describe migration patterns and exposure to pollutants.
Further research efforts on baleen hormones should focus on completing
biological validations to better understand the hormone measurements in baleen
and its correlation with measurements from alternative sample matrices (i.e.,
feces, skin, blubber, and respiratory vapors).
References:
Atkinson, S.,
Crocker, D., Houser, D., Mashburn, K., 2015. Stress physiology in marine
mammals: how well do they fit the terrestrial model? J. Comp. Physiol. B. 185,
463–486. https://doi.org/10.1007/s00360-015-0901-0.
Ayres, K.L.,
Booth, R.K., Hempelmann, J.A., Koski, K.L., Emmons, C.K., Baird, R.W.,
Balcomb-Bartok, K., Hanson, M.B., Ford, M.J., Wasser, S.K., 2012. Distinguishing
the impacts of inadequate prey and vessel traffic on an endangered killer whale
(Orcinus orca) population. PLoS ONE.
7, e36842. https://doi.org/10.1371/journal.pone.0036842.
Fernández
Ajó, A.A., Hunt, K., Uhart, M., Rowntree, V., Sironi, M., Marón, C.F., Di
Martino, M., Buck, L., 2018. Lifetime glucocorticoid profiles in baleen of
right whale calves: potential relationships to chronic stress of repeated
wounding by Kelp Gull. Conserv. Physiol. 6, coy045. https://doi.org/10.1093/conphys/coy045.
Hunt, K.,
Lysiak, N., Moore, M., Rolland, R.M., 2017a. Multi-year longitudinal profiles
of cortisol and corticosterone recovered from baleen of North Atlantic right
whales (Eubalaena glacialis). Gen.
Comp. Endocrinol. 254: 50–59. https://doi.org/10.1016/j.ygcen.2017.09.009.
Hunt, K.E.,
Hunt, K.E., Lysiak, N.S., Matthews, C.J.D., Lowe, C., Fernández-Ajo, A.,
Dillon, D., Willing, C., Heide-Jørgensen, M.P., Ferguson, S.H., Moore, M.J.,
Buck, C.L., 2018. Multi-year patterns in testosterone, cortisol and
corticosterone in baleen from adult males of three whale species. Conserv.
Physiol. 6, coy049. https://doi.org/10.1093/conphys/coy049.
Hunt, K.E.,
Hunt, K.E., Lysiak, N.S., Moore, M.J., Rolland R.M., 2016. Longitudinal
progesterone profiles in baleen from female North Atlantic right whales
(Eubalaena glacialis) match known calving history. Conserv. Physiol. 4, cow014.
https://doi.org/10.1093/conphys/cow014.
Hunt, K.E.,
Lysiak, N.S., Moore, M.J., Seton, R.E., Torres, L., Buck, C.L., 2017b. Multiple
steroid and thyroid hormones detected in baleen from eight whale species.
Conserv. Physiol. 5, cox061. https://doi.org/10.1093/conphys/cox061.
Hunt, K.E.,
Moore, M.J., Rolland, R.M., Kellar, N.M., Hall, A.J., Kershaw, J., Raverty,
S.A., Davis, C.E., Yeates, L.C., Fauquier, D.A., Rowles, T.K., Kraus, S.D.,
2013. Overcoming the challenges of studying conservation physiology in large
whales: a review of available methods. Conserv. Physiol. 1: cot006. https://doi.org/10.1093/conphys/cot006.
Hunt, K.E.,
Stimmelmayr, R., George, C., Hanns, C., Suydam, R., Brower, H., Rolland, R.M.,
2014. Baleen hormones: a novel tool for retrospective assessment of stress and
reproduction in bowhead whales (Balaena mysticetus). Conserv. Physiol. 2,
cou030. doi: https://doi.org/10.1093/conphys/cou030.
Lysiak, N.,
Trumble, S., Knowlton, A., Moore, M., 2018. Characterizing the duration and
severity of fishing gear entanglement on a North Atlantic right whale
(Eubalaena glacialis) using stable isotopes, steroid and thyroid hormones in
baleen. Front. Mar. Sci. 5: 168. https://doi.org/10.3389/fmars.2018.00168.
Marón, C.F.,
Beltramino, L., Di Martino, M., Chirife, A., Seger, J., Uhart, M., Sironi, M.,
Rowntree, V.J., 2015b Increased wounding of southern right whale (Eubalaena
australis) calves by Kelp Gulls (Larus dominicanus) at Península Valdés,
Argentina., PLoS ONE. 10, p. e0139291. https://doi.org/10.1371/journal.pone.0139291.
Marón, C.F.,
Rowntree, V.J., Sironi, M., Uhart, M., Payne, R.S., Adler, F.R., Seger, J.,
2015a. Estimating population consequences of increased calf mortality in the
southern right whales off Argentina. SC/66a/BRG/1 presented to the IWC
Scientific Committee, San Diego, USA. Available at: https://iwc.int/home
Rolland,
R.M., Graham, K.M., Stimmelmayr, R., Suydam, R. S., George, J.C., 2019. Chronic
stress from fishing gear entanglement is recorded in baleen from a bowhead
whale (Balaena mysticetus). Mar. Mam. Sci. https://doi.org/10.1111/mms.12596.
Romero, L.M.,
Platts, S.H., Schoech, S.J., Wada, H., Crespi, E., Martin, L.B., Buck, C.L.,
2015. Understanding Stress in the Healthy Animal – Potential Paths for
Progress. Stress. 18(5), 491-497.
Rowntree,
V.J., Uhart, M.M., Sironi, M., Chirife, A., Di Martino, M., La Sala, L.,
Musmeci, L., Mohamed, N., Andrejuk, J., McAloose, D., Sala, J., Carribero, A.,
Rally, H., Franco, M., Adler, F., Brownell, R. Jr, Seger, J., Rowles, T., 2013.
Unexplained recurring high mortality of southern right whale Eubalaena
australis calves at Península Valdés, Argentina. Mar. Ecol. Prog. Ser. 493:275–289. https://doi.org/10.3354/meps10506.
Sironi, M. Rowntree, V.,
Di Martino, M., Alzugaray, L.,Rago, V., Marón, C.F., Uhart M., 2018. Southern
right whale mortalities at Península Valdes, Argentina: updated information for
2016-2017. SC/67B/CMP/06 presented to the IWC Scientific Committee, Slovenia.
Available at: https://iwc.int/home.
Sironi, M.
Rowntree, V., Snowdon, C., Valenzuela, L., Marón C., 2009. Kelp Gulls (Larus
dominicanus) feeding on southern right whales (Eubalaena australis) at
Península Valdes, Argentina: updated estimates and conservation implications.
SC/61/BRG19. presented to the IWC Scientific Committee, Portugal. Available at:
https://iwc.int/home.
Thomas, P.,
Uhart, M., McAloose, D., Sironi, M., Rowntree, V.J., Brownell, Jr. R., Gulland,
F.M.D., Moore, M., Marón, C., Wilson, C., 2013. Workshop on the southern right
whale die-off at Península Valdés, Argentina. SC/60/BRG15 presented to the IWC
Scientific Committee, South Korea. Available at: https://iwc.int/home
Wasser, S.K.,
Lundin, J.I., Ayres, K., Seely, E., Giles, D., Balcomb, K., Hempelmann, J.,
Parsons, K., Booth, R., 2017. Population growth is limited by nutritional
impacts on pregnancy success in endangered Southern Resident killer whales
(Orcinus orca). PLoS ONE. 12, e0179824. https://doi.org/10.1371/journal.pone.0179824.
By Leila Lemos, PhD Student (hopefully PhD candidate soon)
Here I am with the first GEMM Lab blog post of 2018.
Many people begin a New Year thinking about the future and planning goals to achieve in the following year, and that’s exactly how I am starting my year. After two and a half years of my PhD program, my classes and thesis project are nearing the end. However, a large hurdle stands between me and my finish line: my preliminary exams (as opposed to final exams that happen when I defend my thesis).
Oregon State University requires two sets of preliminary examinations (a.k.a. “prelims”) in order to become a PhD candidate. Thus, planning my next steps is essential in order to accomplish my main objective: a successful completion of these two exams.
The first set of exams comprises written comprehensive examinations to be taken over the course of a week (Monday to Friday), where each day belongs to a different member of my committee. The second type of exam is an oral preliminary examination, conducted by my doctoral committee. The written and oral prelims may cover any part of my proposed research topic as described in the proposal I submitted during my first PhD year.
In order to better understand this entire process, I met with Dr. Carl Schreck, a Fisheries and Wildlife Department professor and one of the members of my committee. He has been through this prelim process many times with other students and had good advice for me regarding preparation. He told me to meet with all of my committee members individually to discuss study material and topics. However, he said that I should first define and introduce myself with a title to each committee member, so they know how to base and frame exam questions. But, how do I define myself?
How do you define yourself? Source: www.johngarvens.com/wpcontent/uploads/2013/02/how_do_you_ define_yourself.jpeg
As part of my PhD committee, Dr. Schreck is familiar with my project and what I am studying, so he suggested the title “Conservation Physiologist”. But, do I see myself as a Conservation Physiologist? Will this set-up have implications for my future, such as the type of job I am prepared for and able to get?
I can see it is important to get this title right, as it will influence my exam process as well as my scientific career. However, it can be hard and somewhat tricky when trying to determine what is comprised by your work and what are the directions you want to take in your future. I believe that defining the terms conservationist and physiologist, and what they encompass, is a good first step.
To me, a conservation specialist works for the protection of the species, their habitats, and its natural resources from extinction and biodiversity loss, by identifying and mitigating the possible threats. A conservation specialist’s work can help in establishing new regulations, conservation actions, and management interventions. As for an animal physiology specialist, their research may focus on how animals respond to internal and external elements. This specialist often studies an animal’s vital functions like reproduction, movement, growth, metabolism and nutrition.
According to Cooke et al. (2013), conservation specialists focus on population characteristics (e.g., abundance and structure) and indicators of responses to environmental perturbations and human activities. Thus, merging conservation and physiology disciplines enables fundamental understanding of the animal response mechanisms to such threats. Using animal physiology as a tool is valuable for developing cause-and-effect relationships, identifying stressor thresholds, and improving ecological model predictions of animal responses. Thus, conservation physiology is an inter-disciplinary field that provides physiological evidence to promote advances in conservation and resource management.
My PhD project is multidisciplinary, where the overall aim is to understand how gray whales are physiologically responding to variability in ambient noise, and how their hormone levels vary across individual, time, body condition, location, and noise levels. I enjoy many aspects of the project, but what I find myself most excited about is linking information about sex, age, body condition, and cortisol levels to specific individuals we observe multiple times in the field. As we monitor their change in body condition and hormones, I am highly motivated to build these whale ‘life-history stories’ in order to better understand patterns and drivers of variability. Although we have not yet tied the noise data into our analyses of whale health, I am very interested to see how this piece of the puzzle fits into these whale ‘life-history stories’.
In this study, animal physiology facilitates our stories. Scientific understanding is the root of all good conservation, so I believe that this project is an important step toward improved conservation of baleen whales. Once we are able to understand how gray whales respond physiologically to impacts of ocean noise, we can promote management actions that will enhance species conservation.
Thus, I can confidently say, I am a Conservation Physiologist.
Me, in Newport, OR, during fieldwork in 2017. Source: Sharon Nieukirk, 2017.
Over the next three months I will be meeting with my committee members and studying for my prelims. I hope that this process will prepare me to become a PhD candidate by the time my exams come around in March. Then, I will have accomplished my first goal of 2018, so I can go on to plan for the next ones!
References:
Cooke SJ, Sack L, Franklin CE, Farrell AP, Beardall J, Wikelski M, and Chown SL. What is conservation physiology? Perspectives on an increasingly integrated and essential science. Conserv Physiol. 2013; 1(1): cot001. Published online 2013 Mar 13. doi: 10.1093/conphys/cot001.