By Dawn Barlow, PhD Candidate, OSU Department of Fisheries, Wildlife and Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
“What is the weather going to be like tomorrow?” “How long will it take to drive there, with traffic?” We all rely on forecasts to make decisions, such as whether to bring a rain jacket, when to get in the car to arrive at a certain destination on time, or any number of situations where we want a prediction of what will happen in the near future. Statistical models underpin many of these examples, using past data to inform future predictions.
Early on in graduate school, I was told that “all models are wrong, but some models work.” Any model is essentially a best approximation, using mathematical relationships, of how we understand a pattern. Models are powerful tools in ecology, enabling us to distill complex, dynamic, and interacting systems into terms and parameters that can be quantified. This ability can help us better understand our study systems and use that understanding to make predictions. We will never be able to describe every nuance of an ecosystem. Instead, the challenge is to collect enough information to build an informed model that can enhance our understanding, without over-simplifying or unnecessarily complicating the system we aim to describe. As Dr. Simon Levin stated in his 1989 seminal paper:
“A good model does not attempt to reproduce every detail of the biological system; the system itself suffices for that purpose as the most detailed model of itself. Rather, the objective of a model should be to ask how much detail can be ignored without producing results that contradict specific sets of observations, on particular scales of interest.”1
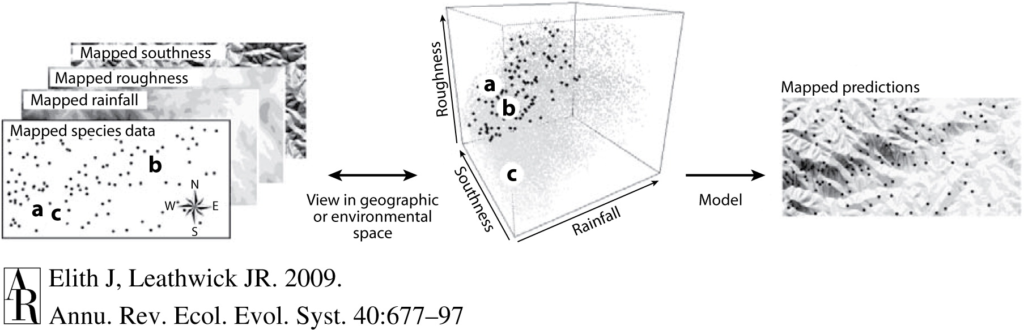
Species distribution models (SDMs) are the particular branch of models that underpin much of my PhD research on blue whale ecology and distribution in New Zealand. SDMs are mathematical algorithms that correlate observations of a species with environmental conditions at their observed locations to gain ecological insight and predict spatial distributions of the species (Fig. 1)2. The model is a best attempt to quantify and describe the relationships between predictors, e.g., temperature and the observed species distribution pattern. For example, blue whale occurrence is higher in areas of lower temperatures and greater krill availability, and these relationships can be described with models3. So, a model essentially takes all the data available, and synthesizes that information in terms of the relationships between the predictors (environment) and response (species occurrence). Then, we can look at the fitted relationships to ask what we would expect from the species distribution pattern when temperature, or krill availability, or any other predictor, is at a particular value.

So, if a model is simply a mathematical description of how terms interact to produce a particular outcome, how do predictions work? To make a spatial prediction, e.g., a map of the probability of a species being present, you need two things: a model describing the functional relationships between species presence and your environmental predictors, and the values of your predictor variables on the day you are interested in predicting to. For example, you may need to obtain a map of sea surface temperature, productivity, temperature anomaly, and surface currents on a day you want to know where whales are expected to be. Your model is the applied across that stack of spatial environmental layers and, based on the functional relationships derived by the model, you get an estimate of the probability of species occurrence based on the temperature, productivity, anomaly, and surface current values at each location. By applying the model over a range of values, you can obtain a continuous surface with the probability of presence, in the form of a map. These maps are typically for the past or present because that is when we can typically acquire spatial environmental layers. However, to make predictions for a future time of interest, we need to have spatial environmental layers for the future.
Forecasts are predictions for the future. Recent advances in technology and computing have led to an emergence of environmental and ecological forecasting tools that are being developed around the world to produce marine forecasts. These tools include predictions of the physical environment such as ocean temperatures or currents, and biological patterns such as where species will be distributed in space and timing of events like salmon spawning or lobster landings4. The ability to generate forecast of marine ecosystems is of particular interest to resource users and managers because it can allow them to be proactive rather than reactive. Forecasts enable us to anticipate events or patterns and prepare, rather than having to respond in real-time or after the fact.
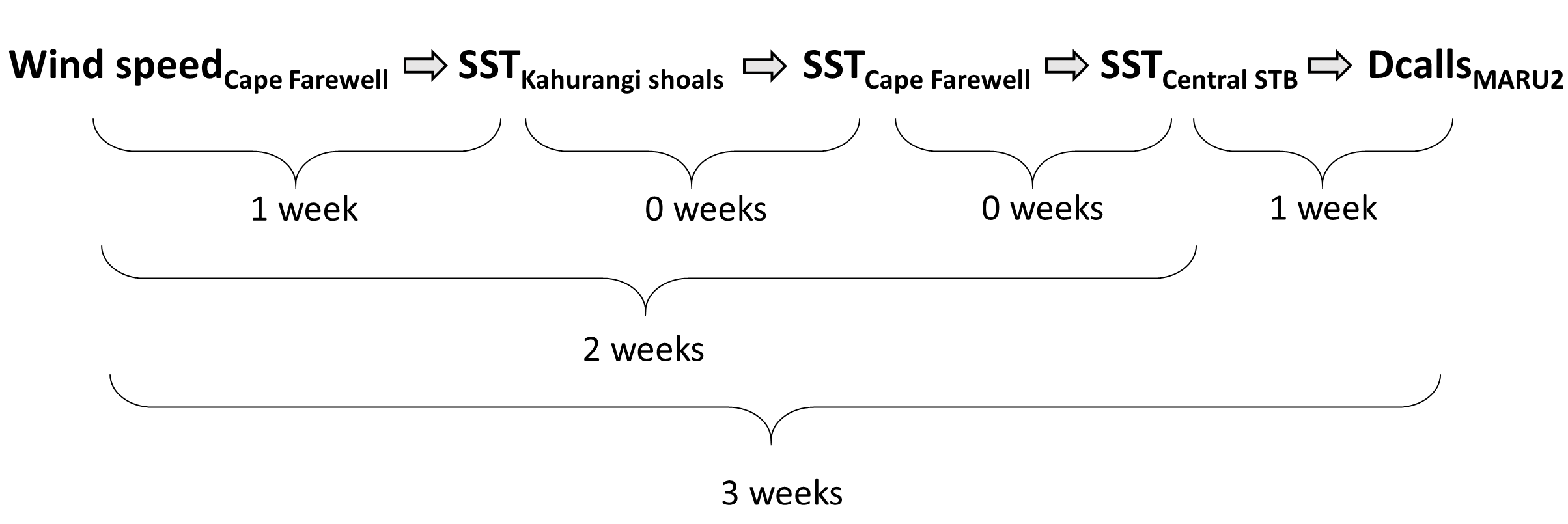
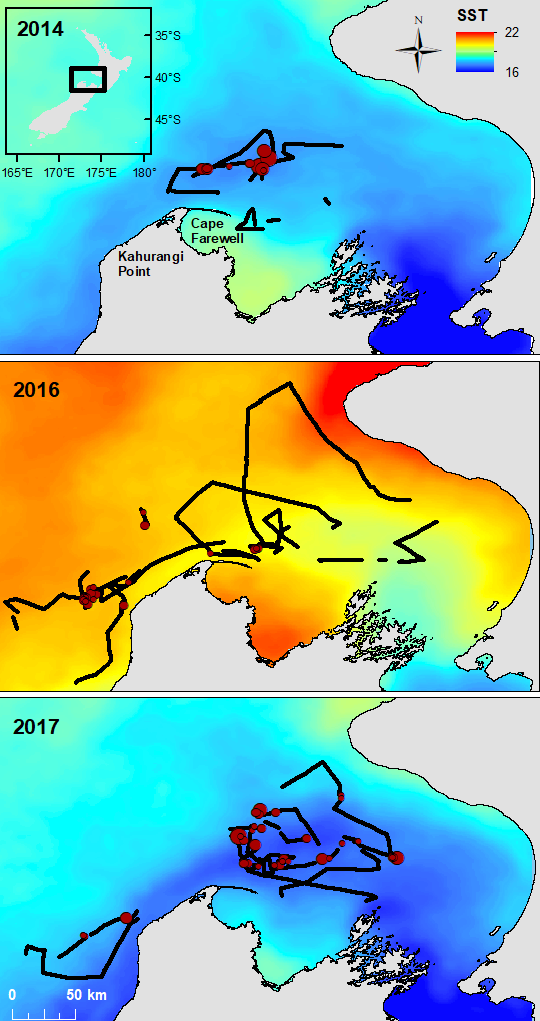
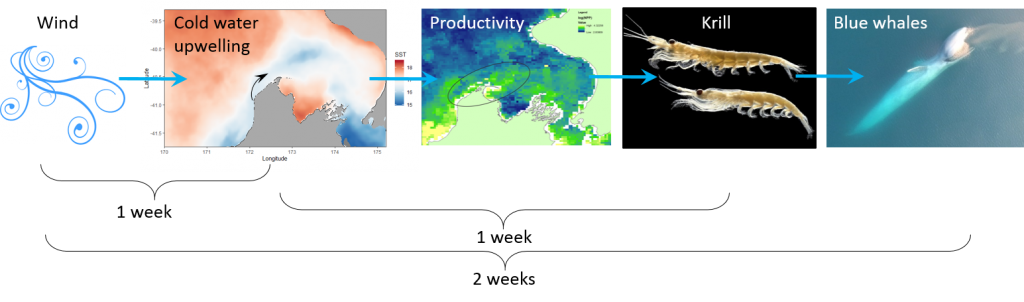
The South Taranaki Bight region in New Zealand is an area where blue whale foraging habitat frequently coincides with industry pressures, including petroleum and mineral extraction, exploration for petroleum reserves using seismic airgun surveys, vessel traffic between ports, and even an ongoing proposal for seabed mining5. Static spatial restrictions to mitigate impacts from these activities on blue whales may be met with resistance from industry user groups, but dynamic spatial management6–8 of blue whale habitat could be more attractive and acceptable. The key for successful dynamic management is knowing where and when to put those boundaries; and this is where ecological forecast models can show their strength. If we can predict suitable blue whale habitat for the future, proactive regulations can be applied to enhance conservation management in the region. Can we develop reliable and useful ecological forecasts for the South Taranaki Bight? Well, given that we have already developed robust models of the relationships between blue whales and their habitat3 and have documented the spatial and temporal lags between wind, upwelling, and blue whales9, we feel confident that we can develop forecast models to predict where blue whales will be in the STB region. As we continue working hard toward this goal, we invite you to check back for our findings in the future. So, consider this blog post a forecast of sorts, and stay tuned!

References:
1. Levin, S. A. The problem of pattern and scale. Ecology 73, 1943–1967 (1992).
2. Elith, J. & Leathwick, J. R. Species Distribution Models: Ecological Explanation and Prediction Across Space and Time. Annu. Rev. Ecol. Evol. Syst. 40, 677–697 (2009).
3. Barlow, D. R., Bernard, K. S., Escobar-Flores, P., Palacios, D. M. & Torres, L. G. Links in the trophic chain: Modeling functional relationships between in situ oceanography, krill, and blue whale distribution under different oceanographic regimes. Mar. Ecol. Prog. Ser. 642, 207–225 (2020).
4. Payne, M. R. et al. Lessons from the first generation of marine ecological forecast products. Front. Mar. Sci. 4, 1–15 (2017).
5. Torres, L. G. Evidence for an unrecognised blue whale foraging ground in New Zealand. New Zeal. J. Mar. Freshw. Res. 47, 235–248 (2013).
6. Hyrenbach, K. D., Forney, K. A. & Dayton, P. K. Marine protected areas and ocean basin management. Aquat. Conserv. Mar. Freshw. Ecosyst. 10, 437–458 (2000).
7. Maxwell, S. M. et al. Dynamic ocean management: Defining and conceptualizing real-time management of the ocean. Mar. Policy 58, 42–50 (2015).
8. Oestreich, W. K., Chapman, M. S. & Crowder, L. B. A comparative analysis of dynamic management in marine and terrestrial systems. Front. Ecol. Environ. 18, 496–504 (2020).
9. Barlow, D. R., Klinck, H., Ponirakis, D., Garvey, C. & Torres, L. G. Temporal and spatial lags between wind, coastal upwelling, and blue whale occurrence. Sci. Rep. 11, (2021).