Allison Dawn, GEMM Lab Master’s student, OSU Department of Fisheries, Wildlife and Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
In last week’s blog, GEMM Lab postdoc Dawn Barlow discussed the uncertain future of upwelling response to climate change and how findings from the Shanks et al., 2009 “Paradigm lost? . . .” study implies that nearshore systems are likely decoupled from offshore upwelling processes. In a follow up to that paper, Shanks and co-authors found that the heterogeneity of coastline morphology (i.e., rocky or sandy) across several Oregon nearshore study sites explained zooplankton retention differences. Indeed, not only are there differences between offshore and nearshore upwelling dynamics, but there are also site-specific factors to consider when it comes to understanding changes in zooplankton retention along the Oregon coast (Shanks et. al, 2010).
I spend a lot of time thinking about what drives the variability in abundance and distribution of zooplankton prey of gray whales at our Port Orford study site over our long-term study period (2015-2022). For this blog, I want to briefly touch on a few interconnected dynamics in this nearshore PCFG gray whale foraging site that may affect their prey availability. Specifically, the interplay between shoreline topography, temperature, and habitat complexity.
Interplay between shoreline morphology and thermal fronts
Several years before the “Paradigm lost? . . .” paper, Shanks led a study that investigated how holoplankton (a group of plankton in which mysids and amphipods belong) retention varies across three sites near Cape Arago and one site in Port Orford (Shanks et a., 2003). Here the authors noted that the Port Orford Bight causes an “upwelling shadow”, which is a region of water protected from upwelling-favorable winds. This shadow results in a small-scale warm water feature in the lee of the Port Orford Bight, which may serve as an important retention and recirculation zone for primary productivity (Graham et al., 1997). Discovering this “upwelling shadow” was not the intention of this paper, so the depth and breadth of the warm water plume within our study area has yet to be mapped (see Figure 1 for another West Coast example). However, “upwelling shadows” can act as convergence zones associated with greater zooplankton biomass (Morgan & Fisher, 2010; Ryan et al., 2010, Woodson et al., 2007) and thus may be an important feature to consider in our spatial analyses of drivers of prey availability to gray whales in our Port Orford study region.
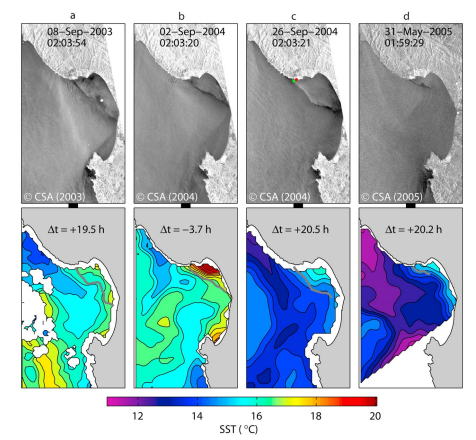
Figure 1. Example of an “upwelling shadow” in Monterey Bay. Remotely sensed oceanographic convergent zones (top panel) and sea surface temperature (SST; lower panel) changes over time: a) Sept 8th 2003, b) Sept 2nd 2004, c) Sept 26th 2004, and d) May 31st 2005. Each time period demonstrates that the lee side of Point Año Nuevo is consistently warmer than the surrounding area. Figure source: Ryan et al., 2020.
Habitat complexity: rugosity and kelp
Not only could the unique shoreline in Port Orford contribute to zooplankton aggregations, but the subtidal marine environment is characterized by a range of unique habitat types: rocky reef, kelp beds, and sandy bottom habitat. Structural habitat complexity has been well documented in coral reef systems to be strongly linked with zooplankton prey availability and biodiversity of planktonic grazers (Richardson et al., 2017; Darling et al., 2017; Kuffner et al., 2007; Gladstone, 2007). Structural complexity can be measured in various ways, but quantifying rugosity (or surface “roughness”) is a widely accepted approach. However, only a few studies have demonstrated predator response to rugose habitats in Oregon nearshore rocky reefs (Rasmuson et al., 2021), and there is a dearth of knowledge linking rugosity to marine mammal predation (Cimino et al., 2020).
Rugosity serves several purposes in the marine environment. A rugose habitat creates micro-habitats for predator evasion, provides greater surface area for kelp recruitment (Cruz et al., 2014; Toohey et al., 2007), and generates turbulence that circulates vital micronutrients for filter-feeding zooplankton and ultimately drives foraging effort at fine scales (Ottersen et al., 2010).
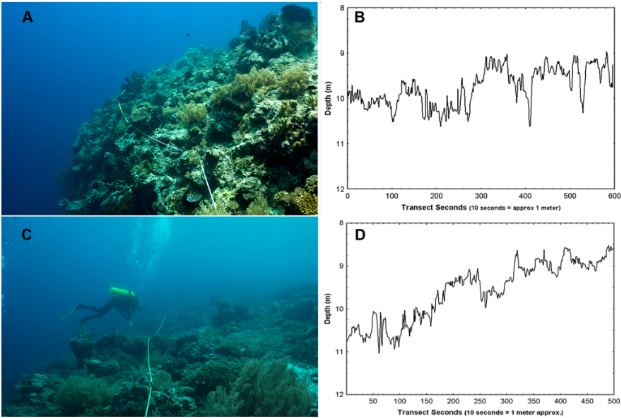
Figure 2. Example images of habitat rugosity as measured by SCUBA transects. A) High-relief coral habitat with B) quantified depth (m) over transect seconds (10 seconds = 1 meter) and C) Low-relief coral habitat with D) quantified depth (m) over transect seconds (10 seconds = 1 meter). Figure source: Dustan et al., 2013.
Rugosity-generated turbidity might also help explain the zooplankton abundance variation we see across our sampling stations in Port Orford. In Lisa’s recent work showing evidence for a trophic cascade, a decline in bull kelp is overall strongly linked to a decline in zooplankton and gray whale foraging in Port Orford. However, there are sampling stations that, despite a significant loss in kelp, still had an abundance of mysids and hosted gray whale feeding activity in 2021 and 2022. Could this mean that those rocky reef stations, which are more rugose than the sandy bottom habitats, produced enough turbulence to support zooplankton prey? This hypothesis is consistent with several studies that found kelp abundance becomes less relevant with increasing habitat complexity (Trebilco et al., 2016; Anderson, 1994; Choat & Ayling et al., 1987; Larson, 1984).
There certainly may be other physical or oceanographic factors that create turbidity at these stations. However, as my REU mentee Zoe Sax has been investigating, we think that turbidity could be a metric of primary productivity, which supports zooplankton growth.
Figure 3 is a map of the average secchi disk values, which provide us with a measure of turbidity (the deeper we see the disk the less turbidity) in 2021 at our 12 sampling stations and their relation to kelp cover.
Last year was a low kelp year, but Mill Rocks still had a few bull kelp canopies. In Mill Rocks where there was rocky reef with kelp, we see secchi values were low (meaning turbidity was high). This is in contrast to the areas in the sandy bottom regions (no kelp, low rugosity: specifically MR16, TC4, TC6, and TC10) with the lightest values, meaning low turbidity.
Then, in Tichenor Cove specifically, we see that station TC1 has very little kelp but high turbidity; interestingly this site was a favored foraging spot for gray whales in 2021 and happens to be the closest station to the “upwelling shadow” I described earlier. I hope to conduct rugosity measurements in the near future so we can investigate these linkages further.

Figure 3. Map of two study sites, Tichenor Cove and Mill Rocks, with twelve sampling stations in Port Orford, OR and their average secchi disk values (meters) in 2021. Kelp abundance shown in light green polygons.
Conclusion
This focus on topography, temperature, and habitat complexity to understand zooplankton variation does not discount that upwelling is an important factor for Oregon nearshore ecology. Menge & Menge 2013 found that upwelling accounted for ~50% of ecological variance in rocky intertidal regions. However, these findings occurred across large spatial areas of about 100 km, while our TOPAZ sampling in Port Orford is on a much finer scale. Variation in ecological patterns at different, hierarchical scales are well-documented (Levin, 1992; Ottersen et al., 2010). Uncovering the “mosaic of processes”, as Shanks et al., 2003 describes, that drives nearshore zooplankton dynamics is equally challenging as it is fascinating, and I look forward to sharing more results from my Master’s work soon.
Did you enjoy this blog? Want to learn more about marine life, research and conservation? Subscribe to our blog and get weekly updates and more! Just add your name into the subscribe box below!
References
Anderson, T. W. (1994). Role of macroalgal structure in the distribution and abundance of a temperate reef fish. Marine ecology progress series. Oldendorf, 113(3), 279-290.
Choat, J. H., & Ayling, A. M. (1987). The relationship between habitat structure and fish faunas on New Zealand reefs. Journal of experimental marine biology and ecology, 110(3), 257-284.
Darling, E. S., Graham, N. A., Januchowski-Hartley, F. A., Nash, K. L., Pratchett, M. S., & Wilson, S. K. (2017). Relationships between structural complexity, coral traits, and reef fish assemblages. Coral Reefs, 36(2), 561-575.
Dustan, P., Doherty, O., & Pardede, S. (2013). Digital reef rugosity estimates coral reef habitat complexity. PloS one, 8(2), e57386.
Gladstone, W. (2007). Selection of a spawning aggregation site by Chromis hypsilepis (Pisces: Pomacentridae): habitat structure, transport potential, and food availability. Marine Ecology Progress Series, 351, 235-247.
Graham, W. M., & Largier, J. L. (1997). Upwelling shadows as nearshore retention sites: the example of northern Monterey Bay. Continental Shelf Research, 17(5), 509-532.
Kuffner, I. B., Brock, J. C., Grober-Dunsmore, R., Bonito, V. E., Hickey, T. D., & Wright, C. W. (2007). Relationships between reef fish communities and remotely sensed rugosity measurements in Biscayne National Park, Florida, USA. Environmental biology of fishes, 78(1), 71-82.
LARSON, R. J., & DeMARTINI, E. E. (1984). SAN ONOFRE, CALIFORNIA. Fishery Bulletin, 82(1-2), 37.
Levin, S. A. (1992). The problem of pattern and scale in ecology. Ecology, 73(6), 1943-1967.
Londoño Cruz et al. (2014) Londoño Cruz E, Mesa-Agudelo LAL, Arias-Galvez F, Herrera-Paz DL, Prado A, Cuellar LM, Cantera J. Distribution of macroinvertebrates on intertidal rocky shores in Gorgona Island, Colombia (Tropical Eastern Pacific) Revista de Biología Tropical. 2014;62(1):189–198. doi: 10.15517/rbt.v62i0.16275
Menge, B. A., & Menge, D. N. (2013). Dynamics of coastal meta-ecosystems: the intermittent upwelling hypothesis and a test in rocky intertidal regions. Ecological Monographs, 83(3), 283-310.
Morgan, S. G., & Fisher, J. L. (2010). Larval behavior regulates nearshore retention and offshore migration in an upwelling shadow and along the open coast. Marine Ecology Progress Series, 404, 109-126.
Ottersen, G., Kim, S., Huse, G., Polovina, J. J., & Stenseth, N. C. (2010). Major pathways by which climate may force marine fish populations. Journal of Marine Systems, 79(3-4), 343-360.
Rasmuson, L. K., Blume, M. T., & Rankin, P. S. (2021). Habitat use and activity patterns of female deacon rockfish (Sebastes diaconus) at seasonal scales and in response to episodic hypoxia. Environmental Biology of Fishes, 104(5), 535-553.
Richardson, L. E., Graham, N. A., Pratchett, M. S., & Hoey, A. S. (2017). Structural complexity mediates functional structure of reef fish assemblages among coral habitats. Environmental Biology of Fishes, 100(3), 193-207.
Ryan, J. P., Fischer, A. M., Kudela, R. M., McManus, M. A., Myers, J. S., Paduan, J. D., … & Zhang, Y. (2010). Recurrent frontal slicks of a coastal ocean upwelling shadow. Journal of Geophysical Research: Oceans, 115(C12).
Shanks, A. L., McCulloch, A., & Miller, J. (2003). Topographically generated fronts, very nearshore oceanography and the distribution of larval invertebrates and holoplankters. Journal of Plankton Research, 25(10), 1251-1277.
Shanks, A. L., & Shearman, R. K. (2009). Paradigm lost? Cross-shelf distributions of intertidal invertebrate larvae are unaffected by upwelling or downwelling. Marine Ecology Progress Series, 385, 189-204.
Shanks, A. L., Morgan, S. G., MacMahan, J., & Reniers, A. J. (2010). Surf zone physical and morphological regime as determinants of temporal and spatial variation in larval recruitment. Journal of Experimental Marine Biology and Ecology, 392(1-2), 140-150.
Toohey, B. D., Kendrick, G. A., & Harvey, E. S. (2007). Disturbance and reef topography maintain high local diversity in Ecklonia radiata kelp forests. Oikos, 116(10), 1618-1630.
Trebilco, R., Dulvy, N. K., Stewart, H., & Salomon, A. K. (2015). The role of habitat complexity in shaping the size structure of a temperate reef fish community. Marine Ecology Progress Series, 532, 197-211.
Woodson, C. B., Eerkes-Medrano, D. I., Flores-Morales, A., Foley, M. M., Henkel, S. K., Hessing-Lewis, M., … & Washburn, L. (2007). Local diurnal upwelling driven by sea breezes in northern Monterey Bay. Continental Shelf Research, 27(18), 2289-2302.
