By Rachel Kaplan, PhD student, Oregon State University College of Earth, Ocean, and Atmospheric Sciences and Department of Fisheries and Wildlife, Geospatial Ecology of Marine Megafauna Lab
What are the most unexpected things you’ve done on Zoom in the last year? Since the pandemic dramatically changed all our lives in 2020, I think we’ve all been surprised by the diversity of things we’ve done remotely. I’ve baked bagels with a friend in Finland, done oceanography labs from my kitchen, had dance parties with people across the country, and conducted an award ceremony for my family’s Thanksgiving scavenger hunt – all on Zoom. Over the last several months, I’ve also mentored an Undergraduate Research, Scholarship, & the Arts (URSA) Engage student, named Amanda. Although we haven’t met in person yet, we’ve been connecting over Zoom since October.
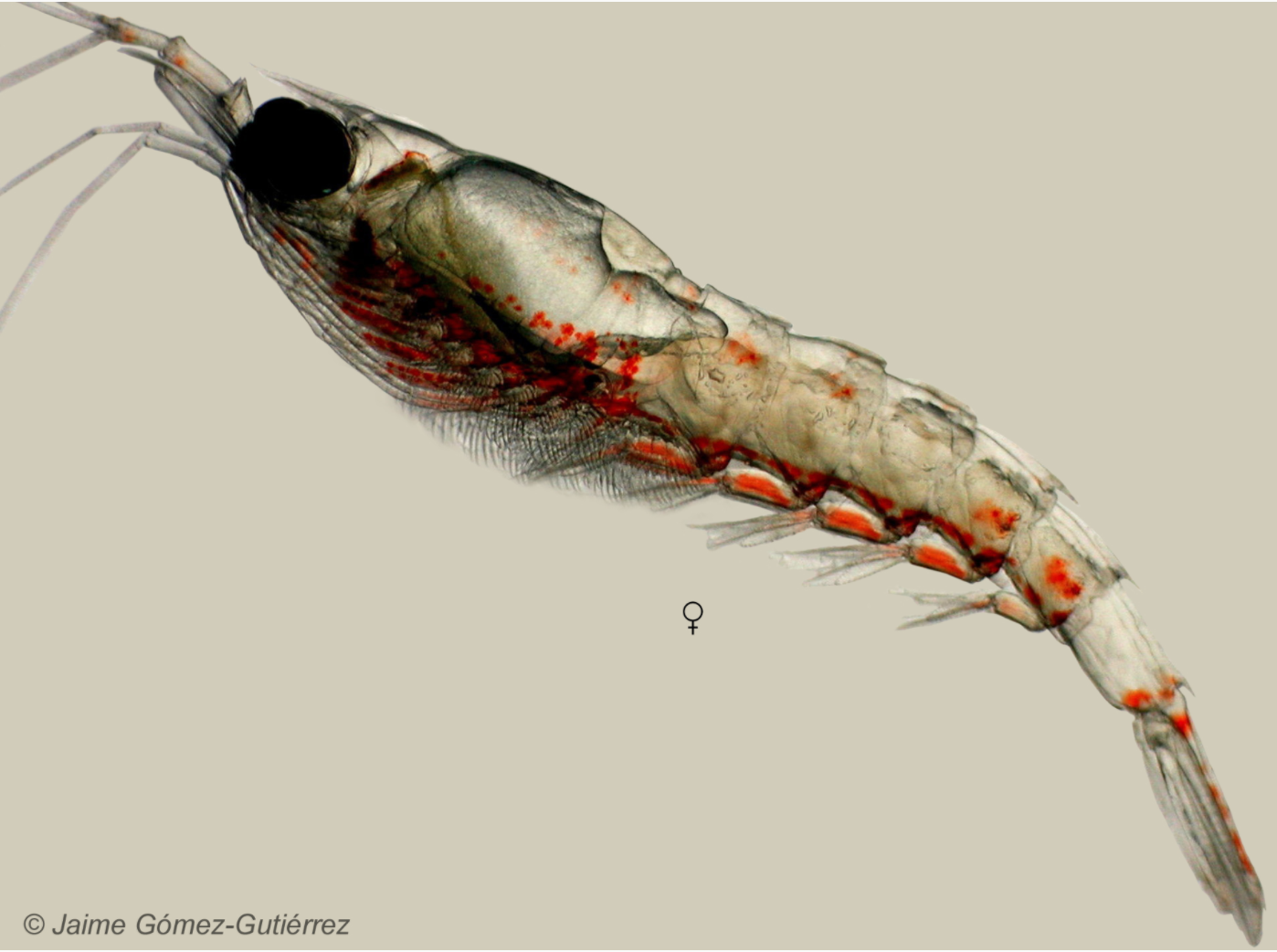
Amanda is an Ocean Sciences student working with me and Dr. Kim Bernard (CEOAS) to conduct a literature review about the two species of krill found off the coast of Oregon. Thysanoessa spinifera and Euphausia pacifica are an important food source for many of the animals that live off our coast — including blue, humpback, and fin whales. I am trying to learn how krill distributions shape those of humpback and blue whales as part of project OPAL, as well as which oceanographic factors drive krill abundances and distributions.
We’re also interested in T. spinifera and E. pacifica for the crucial roles they serve in ecosystems, beyond providing dinner for whales. Krill do many things that are beneficial to ecosystems and people, termed “ecosystem services.” These include facilitating carbon drawdown from the surface ocean to the deep, supporting lucrative fisheries species like salmon, flatfish, and rockfish, and feeding seabirds like auklets and shearwaters. We want to understand more fully the niche that T. spinifera and E. pacifica each fill off the coast of Oregon, which will help us anticipate how these important animals can be impacted by forces such as global climate change and marine management efforts.
Trying to understand the ecosystem services fulfilled by krill is inherently interdisciplinary, which means we have to learn a lot of new things, making this project a lot of fun. The questions Amanda and I have pursued together have ranged from intensely specific, to surprisingly broad. How many calories do blue whales need to eat in a day? How many krill do salmon need to eat? How big are krill fecal pellets, and how fast do they sink?
Trying to answer these questions has basically amounted to a heroic scouring of the internet’s krillscape by Amanda. She has hunted down papers dating back to the 1960s, pulled together findings from every corner of the world, and pursued what she refers to as “treasure troves” of data. In the process, she has also revealed the holes that exist in the literature, and given us new questions. This is the basis of the scientific process: understanding the current state of knowledge, identifying gaps in that knowledge, and developing the questions and methods needed to fill those gaps.
Euphausia pacifica (source: University of Irvine California, Peter J. Bryant).
Filling in knowledge gaps about T. spinifera and E. pacifica can help us better understand these animals, the ecosystems where they live, and the whales and other animals that depend on them for prey. It’s exciting to know that we will have the opportunity to help fill some of these gaps, as both Amanda and I continue this research over the course of our degrees.
Being able to engage in remote research and mentorship has been really rewarding, and it has shown me how far we’ve all come over the last year. Learning how to work together remotely has been crucial as we have adjusted to the funny new normal of the pandemic. As much as I miss working with people in person, I’ve learned that there’s a lot of great connection to be found even in remote collaboration – I’ve loved meeting Amanda’s pets on Zoom, learning about her career goals, and seeing her incredibly artistic representations of the carbon cycle held up to the camera.
Even though most of our conversations take place on Zoom from our homes, this research still feels plugged into a bigger community. Amanda and I also join Kim’s bigger Zooplankton Ecology Lab meetings, which include two other graduate students and eight undergraduate students, all of whom are working on zooplankton ecology questions that span from the Arctic to the Antarctic. Even though we’ve never met in person, a supportive and curious community has developed among all of us, which I know will persist when we can move back to in-person research and mentorship.
Clara Bird, Masters Student, OSU Department of Fisheries and Wildlife, Geospatial Ecology of Marine Megafauna Lab
The field season can be quite a hectic time of year. Between long days out on the water, trouble-shooting technology issues, organizing/processing the data as it comes in, and keeping up with our other projects/responsibilities, it can be quite overwhelming and exhausting.
But despite all of that, it’s an incredible and exciting time of year. Outside of the field season, we spend most of our time staring at our computers analyzing the data that we spend a relatively short amount of time collecting. When going through that process it can be easy to lose sight of why we do what we do, and to feel disconnected from the species we are studying. Oftentimes the analysis problems we encounter involve more hours of digging through coding discussion boards than learning about the animals themselves. So, as busy as it is, I find that the field season can be pretty inspiring. I have recently been looking through our most recent drone footage of gray whales and feeling renewed excitement for my thesis.
At the moment, my thesis has four central questions: (1) Are there associations between habitat type and gray whale foraging tactic? (2) Is there evidence of individualization? (3) What is the relationship between behavior and body condition? (4) Do we see evidence of learning in the behavior of mom and calf pairs? As I’ve been organizing my thoughts, what’s become quite clear is how interconnected these questions are. So, I thought I’d take this blog to describe the potential relationships.
Let’s start with the first question: are there associations between habitat types and gray whale foraging tactics? This question is central because it relates foraging behavior to habitat, which is ultimately associated with prey. This relationship is the foundation of all other questions involving foraging tactics because food is necessary for the whales to have the energy and nutrients they need to survive. It’s reasonable to think that the whales are flexible and use different foraging tactics to eat different prey that live in different habitats. But, if different prey types have different nutritional value (this is something that Lisa is studying right now; check out the COZI project to learn more), then not all whales may be getting the same nutrients.
The next question relates to the first question but is not necessarily dependent on it. It’s the question of individualization, a topic Lisa also explored in a past blog. Within our Oregon field sites we have documented a variety of gray whale foraging tactics (Torres et al. 2018; Video 1) but we do not know if all gray whales use all the tactics or if different individuals only use certain tactics. While I think it’s unlikely that one whale only uses one tactic all the time, I think we could see an individual use one tactic more often than the others. I reason that there could be two reasons for this pattern. First, it could be a response to resource availability; certain tactics are more efficient than others, this could be because the tactic involves capturing the more nutritious prey or because the behavior is less energetically demanding. Second, foraging tactics are socially learned as calves from their mothers, and hence individuals use those learned tactics more frequently. This pattern of maternally inherited foraging tactics has been documented in other marine mammals (Mann and Sargeant 2009; Estes et al. 2003). These questions between foraging tactic, habitat and individualization also tie into the remaining two questions.
My third question is about the relationship between behavior and body condition. As I’ve discussed in a previous blog, I am interested in assessing the relative energetic costs and benefits of the different foraging tactics. Is one foraging tactic more cost-effective than another (less energy out per energy in)? Ever since our lab’s cetacean behavioral ecology class, I’ve been thinking about how my work relates to niche partitioning theory (Pianka 1974).This theory states that when there is low prey availability, niche partitioning will increase. Niche partitioning can occur across several different dimensions: for instance, prey type, foraging location, and time of day when active. If gray whales partition across the prey type dimension, then different whales would feed on different kinds of prey. If whales partition resources across the foraging location dimension, individuals would feed in different areas. Lastly, if whales partition resources across the time axis, individuals would feed at different times of day. Using different foraging tactics to feed on different prey would be an example of partitioning across the prey type dimension. If there is a more preferable prey type, then maybe in years of high prey availability, we would see most of the gray whales using the same tactics to feed on the same prey type. However, in years of low prey availability we might expect to see a greater variety of foraging tactics being used. The question then becomes, does any whale end up using the less beneficial foraging tactic? If so, which whales use the less beneficial tactic? Do the same individuals always switch to the less beneficial tactic? Is there a common characteristic among the individuals that switched, like sex, age, size, or reproductive status? Lemos et al. (2020) hypothesized that the decline in body condition observed from 2016 to 2017 might be a carryover effect from low prey availability in 2016. Could it be that the whales that use the less beneficial tactic exhibit poor body condition the following year?
My fourth, and final, question asks if foraging tactics are passed down from moms to their calves. We have some footage of a mom foraging with her calf nearby, and occasionally it looks like the calf could be copying its mother. Reviewing this footage spiked my interest in seeing if there are similarities between the behavior tactics used by moms and those used by their calves after they have been weaned. While this question clearly relates to the question of individualization, it is also related to body condition: what if the foraging tactics used by the mom is influenced by her body condition at the time?
I hope to answer some of these fascinating questions using the data we have collected during our long field days over the past 6 years. In all likelihood, the story that comes together during my thesis research will be different from what I envision now and will likely lead to more questions. That being said, I’m excited to see how the story unfolds and I look forward to sharing the evolving ideas and plot lines with all of you.
References
Estes, J A, M L Riedman, M M Staedler, M T Tinker, and B E Lyon. 2003. “Individual Variation in Prey Selection by Sea Otters: Patterns, Causes and Implications.” Source: Journal of Animal Ecology. Vol. 72.
Mann, Janet, and Brooke Sargeant. 2009. “ Like Mother, like Calf: The Ontogeny of Foraging Traditions in Wild Indian Ocean Bottlenose Dolphins ( Tursiops Sp.) .” In The Biology of Traditions, 236–66. Cambridge University Press. https://doi.org/10.1017/cbo9780511584022.010.
Pianka, Eric R. 1974. “Niche Overlap and Diffuse Competition” 71 (5): 2141–45.
Soledade Lemos, Leila, Jonathan D Burnett, Todd E Chandler, James L Sumich, and Leigh G. Torres. 2020. “Intra‐ and Inter‐annual Variation in Gray Whale Body Condition on a Foraging Ground.” Ecosphere 11 (4). https://doi.org/10.1002/ecs2.3094.
Torres, Leigh G., Sharon L. Nieukirk, Leila Lemos, and Todd E. Chandler. 2018. “Drone up! Quantifying Whale Behavior from a New Perspective Improves Observational Capacity.” Frontiers in Marine Science 5 (SEP). https://doi.org/10.3389/fmars.2018.00319.
By Leila Lemos, PhD candidate, Fisheries and Wildlife Department, OSU
As previously mentioned in one of Florence’s blog posts, the GEMM Lab holds monthly lab meetings, where we share updates about our research and discuss articles and advances in our field, among other activities.
In a past lab meeting we were asked to bring an article to discuss that had inspired us in the past to conduct research in the marine field or in our current position. I brought to the meeting a literature review regarding methodologies to overcome the challenges of studying conservation physiology in large whales [1]. This article discusses different non-invasive or minimally invasive matrices (e.g., feces, blow, skin/blubber) that can be gathered from whales, and what types of analyses could be carried out, as well as their pros and cons.
One of the possible analyses that can be performed with fecal samples that was discussed in the article is the gut microflora (i.e., bacterial gut community) via genetic analysis. Since my PhD project analyzes fecal samples to determine/quantify stress responses in gray whales, we have since discussed the possibility of integrating this extra parameter to our analysis.
But… what is the importance of analyzing the gut microflora of a whale? What is the relationship between microflora and stress responses? Should we really use our limited sample size, time and money to work on this extra analysis? In order to be able to answer all of these questions, I began reading some articles of the field to better understand its importance and what kind of research questions this analysis can answer.
The gut of a mammal comprises a natural habitat for a large and dynamic community of bacteria [2] that is first developed in early life. Colonization of facultative bacteria (i.e., aerobic bacteria) begins at birth [3], and later, anaerobic bacteria also colonizes the gut. In humans, at the age of 1 year old, the microbiome should have a stable adult-like signature (Fig. 1).
Figure 01: Development of the microbiome in early life. Source: [3]
The gut bacterial community is important for the physiology and pathology of its host and plays an important role in mammal digestion and health [2], responsible for many metabolic activities, including:
fermentation of non-digestible dietary residue and endogenous mucus [2];
recovery of energy [2];
recovery of absorbable nutrients [2];
cellulose digestion [4];
vitamin K synthesis [4];
important trophic effects on intestinal epithelia (cell proliferation and differentiation) [2];
angiogenesis promotion [4];
enteric nerve function [4];
immune structure [2];
immune function [2];
protection of the colonized host against invasion by alien microbes (barrier effect) [2];
Despite all the benefits, the bacterial community might also be potentially harmful when changes in the community composition (i.e., dysbiosis) occur due to the use of antibiotics, illness, stress, aging, lifestyle, bad dietary habits [4], and prolonged food and water deprivation [5]. Thus, potential pathological disorders might emerge when the microbiome community changes, such as allergy, obesity, diabetes, autism, multisystem organ failure, gastrointestinal and prostate cancers, inflammatory bowel diseases (IBD), and cardiovascular diseases [2, 4].
Changes in gut bacterial composition may also alter the brain-gut axis and the central nervous system (CNS) signaling [3]. More specifically, the core pathway affected is the hypothalamic-pituitary-adrenal (HPA) axis, which is activated by physical/psychological stressors. According to a previous study [6], the microbial community in the gut is critical for the development of an appropriate stress response. In addition, the microbial colonization in early life should occur within a certain time window, otherwise an abnormal development of the HPA axis might happen.
However, the gut microbiome can not only affect the HPA axis, but the opposite can also occur [3]. Signaling molecules released by the axis can alter the gastrointestinal (GIT) environment (i.e., motility, secretion, and permeability) [7]. Stress responses, as well as diseases, may also alter the gut permeability, causing the bacteria to cross the epithelial barrier (reducing the overall numbers of bacteria in the gut), activating immune responses that also alter the composition of the bacterial community in the gut [8, 9].
Figure 02: Communication between the brain, gut and microbiome in a healthily and in a stressed or diseased (mucosal inflammation) mammal. Source: [3]
Thus, when thinking about whales, monitoring of the gut microflora might allow us to detect changes caused by factors such as aging, illness, prolonged food deprivation, and stressful events [2, 5]. However, since these are two-way factors, it is important to find an association between bacterial composition alterations and stressful events, such as the presence of predators (e.g., killer whales), illness (e.g., bad body condition), prolonged food deprivation (e.g., low prey availability and high competition), noise (e.g., noisy vessel traffic, fisheries opening and seismic surveys), and stressful reproductive status (e.g., pregnancy and lactating period). Examination of possible shifts in the gut microflora may be able to detect and be linked to many of these events, and also forecast possible chronic events within the population. In addition, the bacterial community monitoring study could aid in validating the hormone data (i.e., cortisol) we have been working with.
Therefore, the main research questions that arise in this context that can aid in elucidating the stress physiology in gray whales are:
What is the microflora community content in guts of gray whales along the Oregon coast?
Is it possible to detect shifts in the gut microflora from our gray fecal samples over time?
How do gut microflora and cortisol levels correlate?
Am I able to correlate shifts in gut microflora with any of the stressful events listed above?
We can answer so many other questions by analyzing the microbiome of baleen whales. Microbiomes are mainly correlated with host diet [10], so the composition of a microbiome can be associated with specific diets and functional gut capacity, and consequently, be linked to other animal populations, which helps to decode evolutionary questions. Results of a previous study on baleen whale microbiomes [10] point out that whales harbor unique gut microbiomes that are actually similar to those of terrestrial herbivores. Baleen whales and terrestrial herbivores have a shared physical structure of the GIT tract itself (i.e., multichambered foregut) and a shared hole for fermentative metabolisms. The multichambered foregut of baleen whales fosters the maintenance of the gut microbiome that is capable of extracting relatively unavailable nutrients from zooplankton (i.e., chitin, “sea cellulose”).
Figure 03: The similarities between whale and other terrestrial herbivore gut microbiomes: sea and land ruminants. Source: [11]
Thus, the importance of studying the gut microbiome of a baleen whale is clear. Monitoring of the bacterial community and possible shifts can help us elucidate many questions regarding diet, overall health, stress physiology and evolution. Thinking about my PhD project, it may also help in validating our cortisol level results. I am confident that a microbiome analysis would significantly enhance my studies on the health and ecology of gray whales.
References
Hunt, K.E., et al., Overcoming the challenges of studying conservation physiology in large whales: a review of available methods.Conservation Physiology, 2013. 1: p. 1-24.
Guarner, F. and J.-R. Malagelada, Gut flora in health and disease.The Lancet, 2003. 360: p. 512–519.
Grenham, S., et al., Brain–gut–microbe communication in health and disease.Frontiers in Physiology, 2011. 2: p. 1-15.
Zhang, Y., et al., Impacts of Gut Bacteria on Human Health and Diseases.International Journal of Molecular Sciences, 2015. 16: p. 7493-7519.
Bailey, M.T., et al., Stressor exposure disrupts commensal microbial populations in the intestines and leads to increased colonization by Citrobacter rodentium.Infection and Immunity, 2010. 78: p. 1509–1519.
Sudo, N., et al., Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice.The Journal of Physiology, 2004. 558: p. 263–275.
Rhee, S.H., C. Pothoulakis, and E.A. Mayer, Principles and clinical implications of the brain–gut–enteric microbiota axis Nature Reviews Gastroenterology & Hepatology, 2009. 6: p. 306–314.
Kiliaan, A.J., et al., Stress stimulates transepithelial macromolecular uptake in rat jejunum.American Journal of Physiology, 1998. 275: p. G1037–G1044.
Dinan, T.G. and J.F. Cryan, Regulation of the stress response by the gut microbiota: Implications for psychoneuroendocrinology.Psychoneuroendocrinology 2012. 37: p. 1369—1378.
Sanders, J.G., et al., Baleen whales host a unique gut microbiome with similarities to both carnivores and herbivores.Nature Communications, 2015. 6(8285): p. 1-8.