By Kim Bernard, Associate Professor, Oregon State University College of Earth, Ocean, and Atmospheric Sciences
Euphausiids, commonly known as “krill”, represent a globally distributed family of pelagic crustacean zooplankton, spanning from tropical to polar oceans. These remarkable organisms inhabit a vast range of marine habitats, from nearshore coastal waters to the expansive open ocean, and from the sea surface to abyssal depths. Notably, they claim the title of the largest biomass among non-domestic animal groups on Earth! Beyond their sheer abundance, euphausiids play a pivotal role in shaping global marine food webs, supporting both economically significant fisheries and extensive populations of marine megafauna.

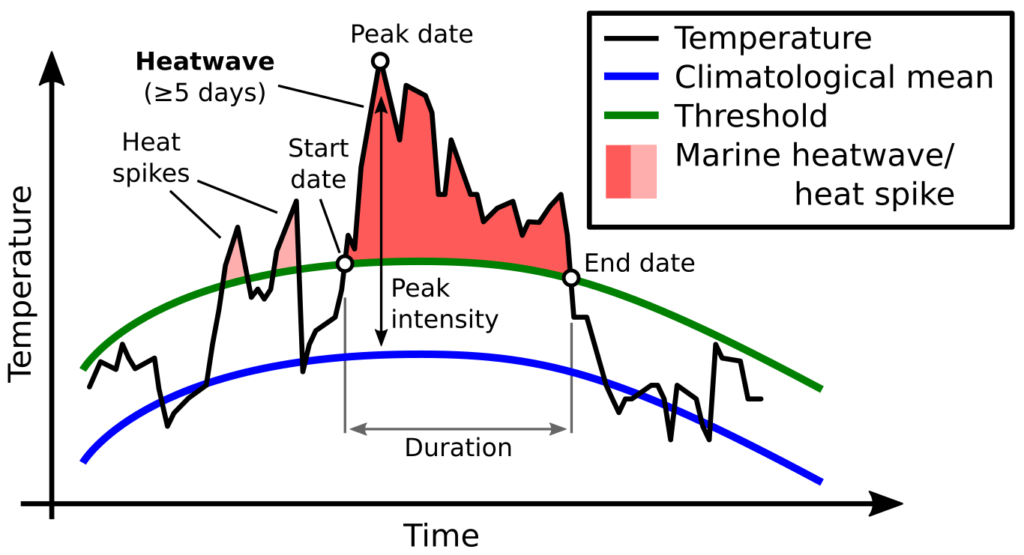
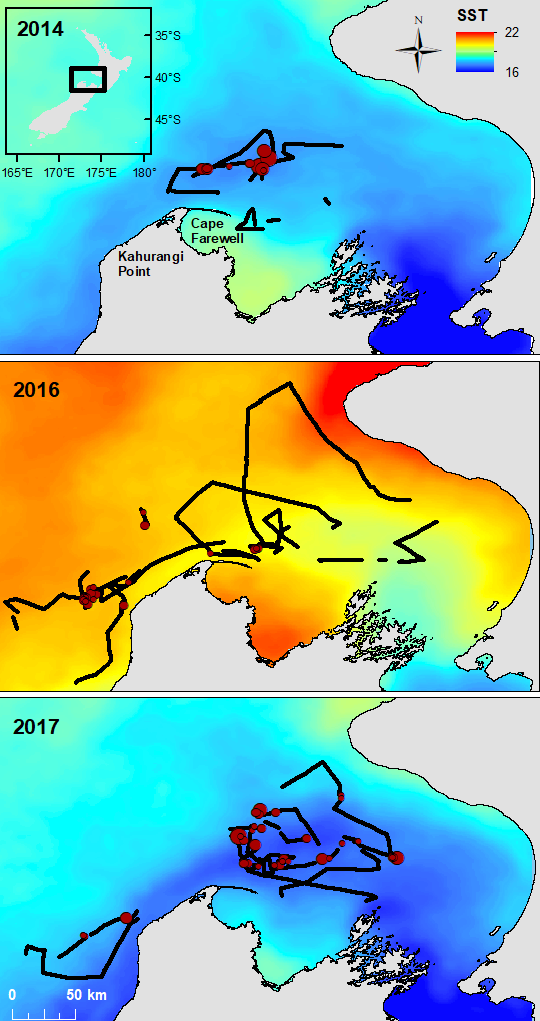
As our planet continues to warm, the ongoing and anticipated shifts in the distribution and biomass of krill populations herald potential disruptions to marine ecosystems and food webs globally. Marine heatwaves, which are expected to increase in frequency, intensity, and duration in the coming decades, have a significant impact on global krill populations, with knock-on effects through food webs. At our home-base off the coast of Oregon, a severe marine heatwave in 2014-2016 resulted in altered krill distributions and reduced biomass, causing a suite of ecological implications ranging from decline in salmon health to increased occurrence of whale entanglements in fishing gear (Daly et al. 2017; Santora et al. 2020).


Here, off the coast of New Zealand, the krill species Nyctiphanes australis (Figure 1) is an important prey item for many marine predators, including slender tuna (Allothunnus fallai), Australian salmon (Kahawai, Arripis trutta), Jack mackerel (Trachurus declivis), short-tailed shearwater (Puffinus tenuirostris) (O’Brien 1988), and of course, the reason we are out here, blue whales (Balaenoptera musculus brevicauda) (Torres et al. 2020). In a precursor study to the SAPPHIRE project, members of our current research team demonstrated the potential negative impacts that marine heatwaves can have off the coast of New Zealand. During that study, our team noted declines in the abundance and changes in the distribution patterns of Nyctiphanes australis during a marine heatwave compared to normal conditions, with subsequent negative impacts on the distribution and behavior of the local New Zealand blue whale population (Barlow et al. 2020). The impetus of the SAPPHIRE project is to improve our understanding of the physiological mechanisms underlying the observed changes in both krill and blue whale populations, with the goal to better predict future changes.
As a zooplankton ecologist and “kriller”, my role on the SAPPHIRE project is to further our knowledge on the prey, Nyctiphanes australis. There are several components to this part of our research: (1) mapping distribution patterns of krill, (2) measuring the quality of krill as prey to whales, and (3) running experiments to test how warming affects krill physiology. To map the krill distribution patterns, we are using active acoustics (Figure 2). To measure the quality of krill, we first need to collect them, and we do that using a Bongo net (Figure 3) that gets towed behind the boat targeting krill we find using the echosounder.

Once we have the krill, we’ll flash freeze them in liquid nitrogen and take them back to Oregon where we’ll measure the amount of protein, fats (lipids), and calories each one contains. Finally, for the experiments on temperature effects, we will use live krill collected with the Bongo net placed individually into 1L Nalgene bottles, each outfitted with oxygen sensors so that we can measure the respiration rates of krill at a range of temperatures they would experience during normal conditions and marine heatwaves (Figure 4).

References
Barlow DR, Bernard KS, Escobar-Flores P, Palacios DM, Torres LG (2020) Links in the trophic chain: modeling functional relationships between in situ oceanography, krill, and blue whale distribution under different oceanographic regimes. Marine Ecology Progress Series 642:207-225. https://doi.org/10.3354/meps13339
Daly EA, Brodeur RD, Auth TD (2017) Anomalous ocean conditions in 2015: impacts on spring Chinook salmon and their prey field. Marine Ecology Progress Series 566:169-182. https://doi.org/10.3354/meps12021
O’Brien DP (1988) Surface schooling behaviour of the coastal krill Nyctiphanes australis (Crustacea: Euphausiacea) off Tasmania, Australia. Marine Ecology Progress Series 42: 219-233.
Santora JA, Mantua NJ, Schroeder ID, Field JC, Hazen EL, Bograd SJ, Sydeman WJ, Wells BK, Calambokidis J, Saez L, Lawson D, Forney KA (2020) Habitat compression and ecosystem shifts as potential links between marine heatwave and record whale entanglements. Nature Communications 11(1):536. doi: 10.1038/s41467-019-14215-w.
Torres LG, Barlow DR, Chandler TE, Burnett JD. 2020. Insight into the kinematics of blue whale surface foraging through drone observations and prey data. PeerJ 8:e8906 https://doi.org/10.7717/peerj.8906