Science Behind the Scenes: Pacific Northwest Pollinator Gardens Post date September 13, 2025 Comments 3
Science Behind the Scenes: Searching for Microplastics in Garden Soils Post date August 30, 2025 Comments 5
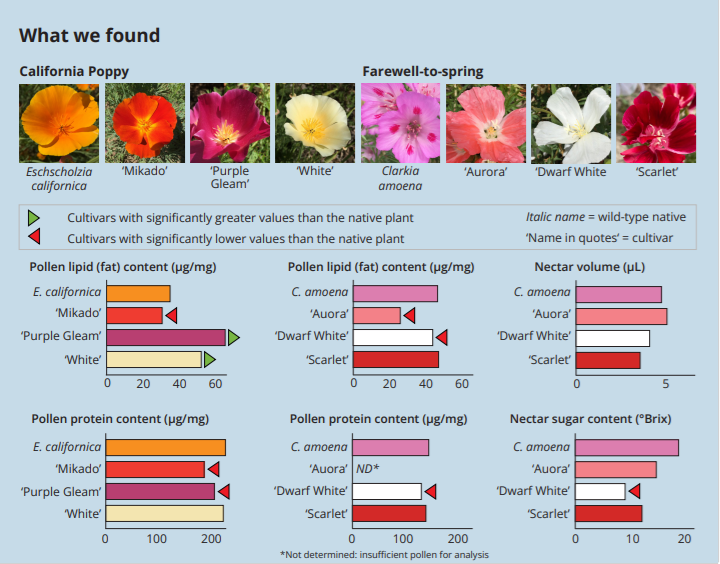
An Update on Native Plant Studies from The Garden Ecology Lab at Oregon State University Post date February 17, 2025 Comments 2
Cultivating Urban Agriculture: Recap of the Grow(in)’ On! Visioning Summit 2024 Post date October 28, 2024