I have been so grateful for the opportunity to assist Jen Hayes with her nativar research project. For the past year and a half, I have learned so much in the world of plant and pollinator relationships. One of the most valuable things I have learned, which I use every day in my own garden, is how to quickly identify bees. In the field, we observe each flower plot for five minutes and record the different things that visit. Being able to identify a fly from a bee, or a bee from a wasp is very important in order to obtain accurate data. I wanted to share the process we use to quickly identify bees in the field and hopefully answer some questions you may have had about what was buzzing in your garden.
A small black Halictidae bee examining the Douglas aster cultivar, Sauvie Snow. The photo was taken by Tyler Spofford at Oak Creek Center for Urban Horticulture in Corvallis, OR.
Once the sun has risen and the chill of the morning has left, bees begin their foraging routine. Male bumblebees, out foraging too late, emerge from the layers of Zinnia flowers. Hundreds of bees possibly pass through your garden in a single day, from flower to flower, collecting pollen and nectar. But what are they? Are they native? Are they helpful in the garden? These are all questions I have whenever I see something buzzing on the mint, exploring the flower patch, or pollinating the tomatoes.
The first things I always look for are the antennae and the number of wings. The flies that are most often confused with bees have short, almost non-existent antennae, whereas bees will have noticeable, segmented antennae. The wings are also something to look: flies only have one pair of wings and bees have two, the forewings and hindwings. Be sure to look closely! There are flies known as hoverflies or syrphid flies that have incredible mimicry adaptations. Look at these two insects on this butterfly bush. Although almost identical, you can see the top insect does not have noticeable antennae. That’s because it is a honey bee mimic!
The bottom insect is a honey bee and the top insect is a syrphid fly that is a mimic of honey bees. The photo was taken by Tyler Sato Spofford at Lewis Brown Horticulture Farm in Corvallis, OR.
If you have established that it is a bee and not a fly, there are other things to look for to identify the bee to a more specific taxonomic level. The coloration of the bee could help if it is green or red, but there are many bees that have different variations of black and white. What I like to look for next is the pollen baskets, also known as the corbiculae. Megachilidae bees carry the pollen on the underside of their abdomen, like bright yellow furry bellies. Another distinguishing factor for Megachilidae bees is how they fly. The abdomen of the bee will usually curl upwards while in flight. Mellisodes bees carry the pollen on their hind legs but the baskets are dramatically bigger than honeybees or bumbles. We like to think of them as pollen pants! Mellisodes bees are also known as long-horn bees because of their disproportionately long antennae.
A mellisodes collecting pollen. Look at those pollen pants!
If the bee does not have any special coloration or noticeably different corbiculae, it could be one of many other genera we have in Oregon. Halictidae bees range in size from the tip of a pen to the size of a penny. They are usually black or black and white and are VERY difficult to distinguish in the field. There are details we have to look for in the lab such as the number of “panels” in the wings or if they have one versus two sub-antennal sutures.
The other bees we see while doing research include wasps, honeybees, and bumblebees. There are so many variations of Bombus here in Oregon it is almost like a scavenger hunt. Because of all the color and striping variations, we use the PNW Bumblebee Atlas to help us identify species in the field.
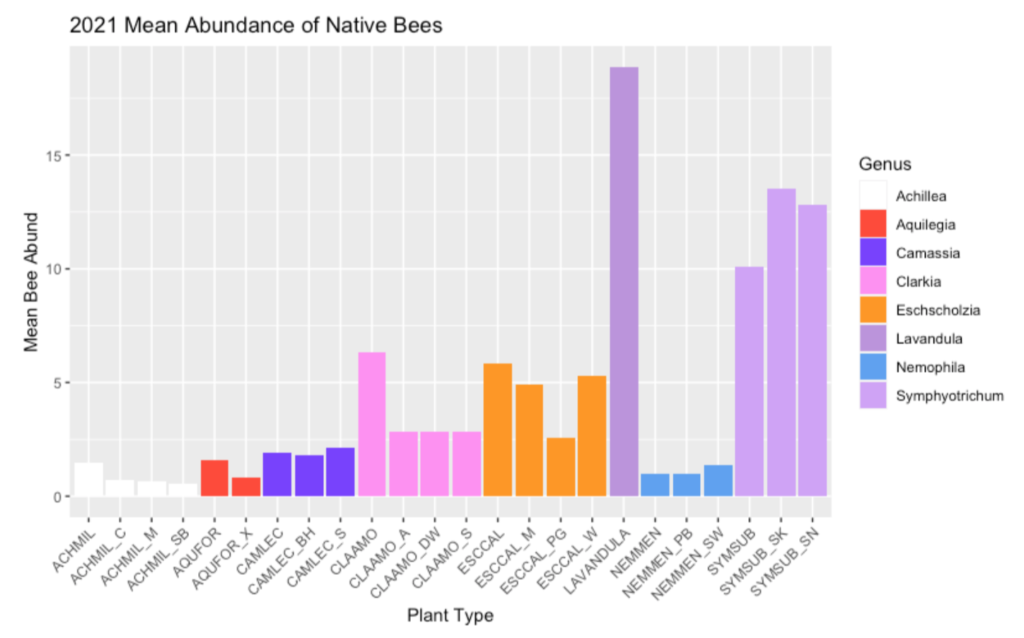
Our second field season studying pollinator visitation to Oregon native plants and native cultivars spanned from April to late September of 2021, although if Douglas Aster had any say in the matter, we would likely still be sampling. The densely blooming Symphyotrichum subspicatum continued to produce a smattering of new flowers through November of last year, and we predict it will do the same this year, too!
Our field crew this summer included Tyler, Svea, Mallory and I. Together, we sampled on 33 different dates across the growing season, allowing us to collect around 2000 physical pollinator specimens, and observe 6,225 unique interactions between pollinators and our study plants! This season we conducted floral trait measurements (including the dimensions of flowers), took multispectral photos, and additionally collected pollen from a subset of our study plants.
From left to right: Mallory vacuum-sampling off of Douglas Aster 'Sauvie Snow', Tyler shaking a farewell-to-spring flower to get pollen off of it, and Svea photographing Baby Blue Eyes 'Penny Black'.
This year, we introduced a third cultivar for California poppy (Eschscholzia californica ‘Purple Gleam’), yarrow (Achillea millefolium ‘Moonshine’), and farewell-to-spring (Clarkia amoena ‘Scarlet’). The new cultivars were established in the spring, which resulted in a late bloom for the annuals, so we expect to see them blooming during their typical period in 2022. The Achillea ‘Moonshine’ replaced Achillea ‘Salmon Beauty’ in being the most abundant yarrow cultivar; it began blooming almost immediately as it was planted into our field site and is still continuing to push out blooms through October alongside the Douglas Asters.
The plant groups in our study: the larger circles with orange text are the native plants, and the smaller circles and turquoise text are the cultivars. The top row contain the perennials yarrow, western red columbine, great camas, and Douglas aster. The bottom row shows the three annuals farewell-to-spring, California poppy, and baby blue eyes.
In addition to watching new plants bloom in the study garden, we had the opportunity to observe many incredible pollinators in the field this summer. We saw a hummingbird visit the Western Red Columbine, we tried to capture videos of leaf-cutter bees snipping little petal pieces off of farewell-to-spring, and at a neighboring plot we observed a male wool-carder bee section off an entire patch of Salvia for a female bee.
On the left: Farewell-to-spring 'Scarlet' with crescents cut out of the petals by leafcutter bees. Top right: A female wool-carder bee (Anthidium manicatum) collecting trichomes from Yarrow 'Calistoga'. Middle right: A leafcutter bee with a piece of petal from Farewell-to-spring 'Dwarf White'. Bottom right: A leaf cutter bee removing a piece of petal from Farewell-to-spring 'Aurora'.
We were also able to take a couple educational field trips this field season in order to learn about pollinator studies ongoing outside of Oak Creek. In June, we went up to the North Willamette Research and Extension Center in Aurora, OR to listen to three talks about pollinators at the Blueberry Field Day. We learned how to score the productivity of honeybee hives, how to properly don a the top of a bee suit, about blueberry’s best pollinators, and blueberry research projects at the University of Washington.
In August, we made a trip to Bend for a different kind of study… an artistic one! We travelled to the High Desert Museum in order to visit Jasna Guy and Lincoln Best’s exhibit “In Time’s Hum…”. Jasna is a brilliant artist inspired by pollinators, which translates into the subject of her pieces as well as her artistic media. Many of her pieces are made using encaustic (a method of painting using wax, bee’s wax in her case!), dipped directly into bee’s wax, or involve pollinators in some other format, including her color study of pollen, which attempts to replicate the colors of fresh pollen as well as the colors after bees have mixed them with nectar. In the center of exhibit were two cases filled with bees collected and identified by Linc, surrounding some of the dried plant specimens they forage on.
These field trips were a wonderful way to see what other pollinator work is happening in our broader community and to inspire future studies. It was especially exciting to see how Jasna and Linc combined art and science with their exhibit, which is something many of us in the Garden Ecology Lab are interested in.
1. Mallory, Svea, and Jen at the blueberry Field Day. 2. Svea, Jen, Mallory, and Tyler at the High Desert Museum. 3. A panorama of the "... In Time's Hum ... " exhibit. 4-5. Art on the outside of the exhibit. 6. A snapshot of two pollen samples from Jasna Guy's pollen color study.
While we cannot make conclusions until we complete our final field season, we are excited to report some of the variation in visitation between native plants and native cultivars that we have observed in our first two field seasons. In the first field season, our observations of native bees foraging on the study plants revealed three plant groups to have variable amounts of visitation. Yarrow, farewell-to-spring, and California poppy all had at least one cultivar that received substantially less native bee visits than the native type. In our second year, all three of farewell-to-spring’s cultivars received less visitation than the native Clarkia amoena. Poppy had only one cultivar with less native bee activity than the native (Purple Gleam), and in the case of Douglas Aster, both of the cultivars actually had more visitation by native bees than the native.
Figure 1: Average Abundance of Foraging Native Bees during 5-Min Observations in 2021. Individual plants are color-coded by genus. The naming scheme combines the first three letters of the genus and specific epithet; cultivars are denoted by an underscore and a 1-2 letter code to identify them. For example, AQUFOR is the native Aquilegia formosa, and AQUFOR_XT is Aquilegia x ‘XeraTones’.
Flowers and bees have one of the most well-known symbiotic relationships ever formed. Flowers rely on bees for pollination, and bees rely on flowers for nectar and pollen. It is generally understood that flowers act as advertisements to attract bees. However, less is known about what exactly bees are seeing and how that can change once humans get involved. This project is focused on the changes that can arise after a plant is cultivated, and how these changes can affect pollinator preference of a flower.
While changes made by breeders might not seem all that drastic to our eyes, we have little idea if that is the case for bees. Often breeders will change flowers for aesthetic purposes. This can have unknown consequences. These changes might not seem like such a big issue since the flowers are still colorful. However, bee vision is very different from humans, with bees having the ability to see into the UV spectrum. This means that while we might think we are only changing the bloom size or the color, we could also be unintentionally changing UV messaging visible only to the bees.
The purpose of this study is to use UV photography to explore these invisible differences between the native and cultivar. We also want to determine if the differences have a tangible impact on pollinator preference. This study is ongoing, but the images so far have shown a few native/cultivar sets that have a marked difference in UV markers between native and cultivars. While the study has only just started, our excitement and curiosity have not abated. This is an entirely new foray into pollinator relationships and mechanisms and could open up the world of bees and flowers in a brand new way.
An example of a UV photo of a nemophila flower, with a UV marking in the center, highlighted in blue
My name is Mallory Mead, and I am new to the Garden Ecology Lab! I am an undergrad studying Horticulture and minoring in Entomology, and I started a few weeks ago as an assistant to Jen Hayes on her study of pollinator attraction to native plants and nativars.
I enrolled in Oregon State’s URSA Engage program, which gives undergrads a taste of research experience in the Winter and Spring of their first year, and joined a project studying how mason bees might be impacted by climate change with Dr. Jim Rivers of the department of Forest Ecosystems and Society. The study seeks to examine the effects of warming temperatures on mason bee behavior and the development of brood.
The Western US’s native species of mason bee, the Blue Orchard Bee (BOB) is known to be an excellent orchard pollinator. On many orchard crops they are more efficient at pollination than honey bees on a per individual basis, and so the commercial management of BOBs is being explored as honey bee colonies suffer management challenges and colony losses in recent years.
Mason bees have a short lifespan of 4 to 6 weeks. Emerging in the early spring, males die shortly after mating, while females build nests in holes in wood or reeds. They forage for pollen and nectar to form provision masses in which they lay their eggs. They also collect mud to form partitions between each provision mass and to cap the nest once it is full. Their offspring will feed on the provisions and metamorphose into cocooned adults to overwinter in their cells and emerge the following spring.
To ensure the bees had ample nutrient resources, the project was conducted next to the organic cherry orchard at OSU’s Lewis Brown Farm. Before the cherries bloomed, 6 nest structures were designed and constructed by Jim, Ron Spendal (a mason bee house conisuerrier) and Aaron Moore of Revolution Robotics.
Nest structures, solar panels, and camcorders at Lewis Brown Farm.
Each structure has 3 shelves with 16 nest holes each, lined with paper straws so that the nests can be easily removed and examined. The structures are solar powered, and each shelf is heated to a different increment above the ambient temperature i.e. + 0°C , + 2°C, + 4°C, + 6°C, + 8°C, + 10 °C, and + 12°C. These differentials represent many potential warming outcomes of climate change.
Nest Structure Number 2 with labelled component parts. A. The Electronics control box. B. Cocoon-release box. C. Shelves sandwiched by heating pads, and lined with paper nesting straws
Our Hypotheses
We predicted that female mason bees will select the warmer nests first, and that females will leave nests earlier in the morning to begin foraging because they will reach the critical internal temperature necessary for flight sooner.
If heated bees have a greater window of foraging time, then we predict they’ll be able to construct nests at a faster rate in the warmer nests.
With greater nest construction will come a greater production of offspring from the bees in the warmed nests.
But…
In terms of offspring quality, we predict that offspring of heated nests will emerge as weak individuals and mortality will be the highest for the heated brood.
…and we are pretty confident about this last prediction.
Insects are poikilothermic meaning their internal temperatures are determined by the environment. Past studies by researchers Bosch and Kemp have reported that mason bees who are overwintered at warm temperatures will “use up their metabolic reserves and are likely to die during the winter”. And a more recent study by researchers at the University of Arizona found that mason bees subjected to heating resulted in reduced body mass, fat content and high mortality of the mason bee offspring.
Data Collection
One of the latest male mason bees to emerge, surrounded by empty cocoons in the release box.
Our mason bees started hatching from cocoons in mid-April and began to colonize the nest structures. I captured video footage of the bees as they emerged in the morning to forage. If bees from heated nest sites emerge earlier, this will support our hypotheses that they reach their critical-for-flight temperature earlier, and get a leg-up on foraging compared to their neighbors.
I also conducted “nest checks” to track the rate of nest construction along with two other research assistants.
In the fall, the nest tubes will be extracted to examine the reproductive output, and in the following spring, offspring will be assessed for rates of mortality, offspring mass, and fat content.
Obstacles
Some of the challenges along the way have included dealing with insect pests. Spiders were easygoing inhabitants of the nest straws, for they only nested in empty straws, so we’d swap them out for a clean one. The earwigs were much more pervasive, and went for the already inhabited nests. As generalist foragers, the earwigs took advantage of provision balls of nectar and pollen that had not yet been sealed off by mud. Once I read that earwigs will indeed eat the mason bee eggs that are laid into the provision masses, I knew it was crucial to remove the earwigs from all nests, but these feisty creatures proved determined to stay. We ordered some tanglefoot, a sticky substance to trap the earwigs on their way up the structure post, and meanwhile I coaxed earwigs out with tiny pieces of grass. Jabbing them repeatedly would eventually provoke them to charge at the blade of grass and fall out from the straw.
Yellowjackets were another opportunistic nester. They’d sneak into the cocoon boxes to build nests, and always gave me a start when opening the tiny boxes. I removed their nests with an extended grabber tool and would destroy them in any way I could. I feel immensely lucky not to have been stung through this process.
The most terrifying surprise during the project was a fat snake that was living in the solar panel battery box. It popped out at me hissing while I conducted a routine check. Alas, I was too spooked to take on this unexpected visitor and let it leave on its own time.
Preliminary Findings & Observations
By mid-May, a pretty clear pattern was emerging. At each structure, the control shelf’s nests (+ 0 °C) were full and capped with mud, while the hottest shelves were almost completely empty. We will analyze nest check data to confirm that these patterns are not just arising by chance, but a study that was released this past April showed another species of mason bee in Poland following the same pattern of nest site preference and selection for cooler nest sites.
The mason bees’ unexpected behavior of avoiding the heated chambers may lead to trouble during the second part of the experiment because this means our sample size for heated offspring has become so tiny, but here it is important to note that this is mason bee project is a pilot study and so the data collected this year will simply influence more specific future research.
these preliminary findings make me think that mason bees have an ingrained sense to avoid warm nests, which might show mason bees’ adaptability in the face of climate change, that is, if they can manage to continue finding cool nests. People managing mason bees find that nests facing the morning sun are the most attractive to the bees, but I wonder how long it will be before temperatures rise and mason bees start avoiding these sunny nests.
Moving Forward
By the end of May, I’d only see a few the mason bees per visit, so the season was clearly coming to an end. I wrapped up data collection and am now spending the summer extracting data from the video footage, and checking up on the bees to ensure they are safe and sound until Fall inspections.
I am wishing the best to both the wild bees in our region and those in our study, as the temperatures skyrocket this week but with this summer’s heat wave, I don’t think we need to simulate climate change; it is right here before us. Even though it is practically inevitable that temperatures will rise to dangerous heights in my generation’s lifetime, there is so much life to be saved, and there is no time to waste.
“Blue Orchard Bee, Osmia lignaria” by SeabrookeLeckie.com is licensed under CC BY-NC-ND 2.0″
I distinctly remember the day that I decided I wanted to study wild bees. I was sitting in a darkened auditorium at the American Museum of Natural History in New York City, listening to Claire Kremen deliver the plenary address in a symposium focused on invertebrate conservation.
In her address, Dr. Kremen shared the results of her research on watermelon farms in California’s Central Valley. Like all cucurbits, watermelon requires insect pollination to set fruit. Watermelon, in particular, has a high pollination requirement: it takes at least eight bee visits to deposit the 500-1,000 viable pollen grains needed to set harvestable fruit in seeded watermelons. Seedless watermelons require between 16 and 24 visits by a bee in order to set fruit. Most growers meet the high pollination requirement of melons by renting and placing honey bee hives in fields. Dr. Kremen’s research suggested that a different approach might be possible.
Honey bees are important agricultural pollinators, in large part because they can be moved en mass and placed into farm fields at the exact moment that pollination services are needed.Photo by Isabella Messer.
Dr. Kremen and her colleagues studied three types of watermelon fields that varied in their pest management practices (organic or conventional) and their proximity (near or far) to native habitat in the foothills of the California coast range. These fields were: organic near, organic far, and conventional far. Watermelon fields that used conventional pest management practices and that were located near the foothills were not included in this study because this type of farm did not exist in the region. The team determined the number of pollen grains that different bee species deposited on watermelon, by presenting different bees with a single watermelon blossom that had yet to receive any insect visits. After a blossom was visited by a single bee, the flower was bagged and tagged accordingly, and the number of pollen grains deposited on the stigma by that single bee was counted in the lab. They repeated this process for 13 different bee species.
Next, the team sat in watermelon fields and observed what types of bees visited watermelon blossoms, in different types of farm fields. Watermelon flowers are only receptive to pollination visits for a single day. They recorded the sex and species of each bee visitor to the blossoms. Based upon these species specific counts, combined with the pollen deposition data (above), they were able to assess how much each particular bee species contributed to the production of harvest-ready watermelon.
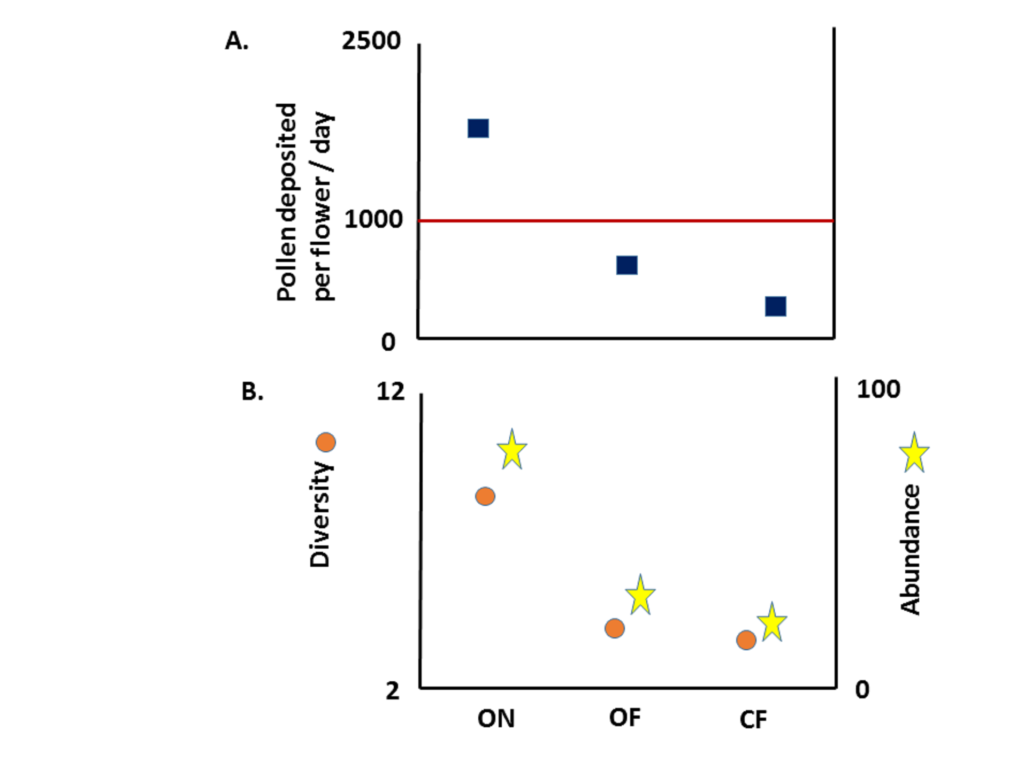
Dr. Kremen found that pollination surpassed 1,000 pollen grains needed to set harvestable fruit per flower in the organic near fields but not in the organic far or the conventional far field (Fig 1, part A). Furthermore, she found that this outcome could be tied to the greater diversity and abundance of bees in organic near fields, compared to the other two types of fields (Fig 1, part B).
Figure 1. The number of pollen grains deposited on watermelon stigmas (A), and the diversity (orange circles) and abundance (yellow stars) of bees (B) on farms that were classified as organic near (ON), organic far (OF), and conventional far (CF). The number of pollen grains required to set marketable fruit (1000) is noted via a red threshold line in A. (modified from Kremen et al. 2002).
Dr. Kremen also found that, even though no single species of wild bee was as effective as managed honey bees, the collective group of wild bees surpassed the effectiveness of honey bees in organic near fields (Figure 2). Interestingly, honey bees were most effective as crop pollinators in the conventional far fields and least effective as crop pollinators in the organic near fields. This may be because few other flowers were in bloom in the conventional far fields, so that honey bees concentrated their attention on the crop at hand. In the organic near fields, a greater diversity of flowering plants likely competed for the pollination services of honey bees.
Figure 2. The cumulative contribution of native bees, compared to the contribution of honey bees to the pollination requirement of watermelons on organic near (orange circles), organic far (navy squares), and conventional far (yellow stars) farms (modified from Kremen et al. 2002).
Wild bees were able to fully satisfy the pollination requirements of a crop with an extremely high pollination requirement because broad spectrum insecticides were not used, and the foothills provided year-round and protected habitat for the bees. This story blew my mind!
Prior to that conference, I had never given wild bees much thought. They’re mostly solitary nesters, with small bodies, that only forage for a few days to a few weeks. They tend to be inefficient foragers, particularly when compared to the juggernaut of a honey bee hive. Whereas wild bees are akin to a single vendor on Etsy, honey bees seemed the unbeatable Amazon!
Dr. Kremen’s work showed the potential value that wild bees have to agriculture. And her work was published just prior to the global onset of colony collapse disorder in honey bees in 2006. It set off a worldwide discussion about what to do about honey bee losses. Should scientists put time and effort into saving a single, non-native species (the honey bee), or should we work to conserve or build habitat around farm fields while also reducing insecticide use?
I was incredibly hopeful that the simultaneous threat to honey bees and promise of wild bees might promote heavier investments in agroecology, including the conservation of bee-friendly habitat around farms. During this time period, I was also in the early stages of documenting wild bee biodiversity in community and residential gardens, and I was surprised that abundance and diversity of garden bees was much higher than I had anticipated.
Back in 2004, I started to see gardens, and the abundance and diversity of wild bees that they host, as a potential solution to the problem of colony collapse disorder. Although I continue to be fascinated by the potential role of home and community gardens as a safe haven for bees from agricultural stresses, the urgency of this question has faded. Colony collapse disorder does not currently plague honey bees, due in large part to federal investments in studying, understanding, and mediating the factors that contribute to failing hives. With honey bees doing much better, attention has somewhat faded on the potential role of wild bees as crop pollinators. Still, work in this area continues and may rise to renewed importance, should colony collapse disorder again present a major challenge to United States agriculture.
Wild bees, including this leaf-cutter bee in the genus Stelis are also potentially important crop pollinators. However, many farm practices, such as regular insecticide sprays and mono-cultural cropping systems, make farms inhospitable to wild bees. Photo by Isabella Messer.
Kremen, Williams, Thorp. 2002. Crop pollination from native bees at risk from agricultural intensification. Proc. Natl. Acad. Sci. 99: 16812-26816. DOI: 10.1073/pnas.262413599.
Our colleague, Brooke Edmunds, was kind enough to shoot and edit this short video on two of our current lab projects: Jen Hayes’ study of native plants and nativars and Tyler Spofford’s study of the economic costs and benefits of growing vegetables in bucket gardens.
As we near the end of our 2020 field season, stay tuned for research updates.
[Preface: For the past few years, I have written a column for the Hardy Plant Society of Oregon’s (HPSO) Quarterly Magazine. It has been a wonderful experience, as the HPSO provides excellent editorial assistance. Below, I share my most recent article for the HPSO Quarterly, and thank Eloise Morgan and her team for helping to improve and elevate my writing.]
I spend my nights thinking about gardens: not about the plants that I want to purchase or the crops that I want to plant. Instead, I puzzle over how to study a system that is incredibly variable (from person to person, or even in the same person’s garden from year to year) and complex (with more plant species than just about any other system that has been studied). Gardens are both wild and managed, and unlike other systems I have worked, it is impossible to divorce human behavior from the ecology and evolution of the garden.
In this series, I wanted to share five scientific studies that have had a large role in shaping how I think about gardens. Because of space limitations, I will share the first study in this article. I will wrap up the remaining four studies, in subsequent issues. The five studies are:
Simberloff and Wilson (1969). This study commenced 54 years ago, and yet remains a ‘must read’ for any ecology student. In 1966, Dan Simberloff and Ed Wilson selected six small mangrove islands off the coast of Florida. The islands varied in distance from the mainland coast, from near to far (Figure 1a), as well as size, from small to large (Figure 1b)
Figure 1. In Simberloff and Wilson’s experiment, they selected mangrove islands that varied in their (a) distance from the mainland (the coastline of Florida) and (b) their size. Attribution: Hdelucalowell15 / CC BY-SA (https://creativecommons.org/licenses/by-sa/4.0)
Simberloff and Wilson constructed a scaffold that encircled the edge of each island, covered the scaffold with a tarp, and then proceeded to ‘defaunate’ each island with methyl bromide pesticide. In other words, they killed every arthropod on the islands. After removing their ‘death tents’, and over the course of the next year, they carefully monitored, cataloged, and counted every arthropod that arrived and survived on each island. What they discovered was formulated into the ‘Theory of Island Biogeography’, or a theory about how organisms colonize new habitat, and assemble into a biological community.
They found that islands that were closer to the mainland coast of Florida were colonized earlier, and accumulated species faster, compared to islands that were farther (Figure 2). They also found that species would accumulate on each island, over time, until a maximum peak is reached (not shown). Then, the number of species would begin to drop, as ecological interactions (such as competition for food) would allow some species to prosper, while others went locally extinct. They found that smaller islands were more prone to species extinctions, than larger islands (Figure 2).
Size, distance, age: those are the three things that Simberloff and Wilson predicted would govern the diversity and assembly of organisms within a habitat.
My first faculty position was at Fordham University in New York City, where I studied pollinators in 18 community gardens in Harlem and in the Bronx. During the course of this study, I was inspired by Simberloff and Wilson. I could not help but see the 600+ community gardens that dot the landscape of New York City as islands of green in a sea of concrete.
We expected that gardens that had been long-established would have more pollinator species than newer gardens. We expected that larger gardens would host more pollinator species than smaller gardens. And, we expected that gardens that were closer to ‘mainland’ sources of pollinators, such as Central Park or the New York Botanical Garden, would have more species of pollinator than those that were distant.
We were wrong on two out of three predictions (Matteson and Langellotto 2010). Larger gardens had more pollinator species than smaller gardens, but neither distance nor age had any impact. I was so disappointed that we did not find an effect of distance, or of garden age. I had visions of ‘revitalizing’ the Theory of Island Biogegraphy for urban landscapes, but it was not to be. If anything, our study suggested that the ‘sea of concrete’ was not exactly a wasteland, afterall. The street trees, potted plants, windowsill gardens, and patio gardens all provided resources for urban pollinators, even in one of the most densely populated and heavily developed cities in the world.
This study showed me that it will be much more difficult to track pollinator movements among urban gardens, than I had hoped. We tried to use a traditional mark-recpture approach (see Matteson and Langellotto 2012), but out of 476 marked butterflies we only found four in a garden other than which it was marked and released. We were searching for the ‘needle’ of small butterflies in the ‘haystack’ of the New York City landscape. My students tried to follow pollinators as they left our study gardens, and almost got hit by a car, as they were running across the street. We played around with the molecular markers of a few bumblebees (see Morath 2007), to see if there was evidence of genetic differentiation, but were stymied by a lack of reliable primers that could help us look for any genetic differences in bees from different gardens. And then I moved to the Willamette Valley, where gardens are islands of green in an ocean of green. Understanding what draws pollinators to particular gardens will be even more difficult in this landscape, where pollinators have so many other choices for finding nectar and pollen.
Based upon our initial results from our Portland Garden study (2017-2019), I think I have a new hypothesis as to what might draw pollinators to home and community gardens. Our second study year (2018) was characterized by a hot and dry summer. Our first sampling season was also dry, but the spring months were wet, and the summer was cooler. In 2018, we collected far more bees (abundance) and more types of bees (species) than we collected in 2017 or 2019. In 2018, the landscape of the Willamette Valley was toast! Almost all flowering plant materials seems to shut down photosynthesis, so that they could conserve pressure water that would otherwise escape through open stomates. In this type of situation, bees seemed to concentrate in home gardens, which seemed to be one of the few places where they could reliably find nectar and pollen.
If this is the case, gardens aren’t necessarily going to be an important source of floral resources across all years. In a good year, there should be other plants in bloom in the greater landscape that bees can use. But in a hot, dry year, gardens may become an even more important refuge for bees. Most gardeners provide irrigation, which extends the bloom season beyond what is natural in the valley. Or, gardeners select plants that can prosper and bloom without supplemental irrigation, such as goldenrod or Douglas aster. It’s important to note that, even in the hot, dry weather of 2018, we still collected more bees from gardens that used drip irrigation, rather than overhead sprinklers. I think that the overhead irrigation physically blocks bees from navigating through a garden, which lessens their abundance and diversity.
Ultimately, I hope that our studies can lead us to a more predictive model of the resource value of home gardens to pollinators. The goal isn’t necessarily to understand what gardeners should do to attract pollinators, but to describe the conditions where gardens become increasingly important to pollinator conservation. In addition, I’d love to describe the value of gardens, relative to other habitat types, to pollinators. And finally, I hope to better understand the direction and movement of pollinators between gardens and other habitat types.
Pollinator syndromes are the characteristics or traits of a flower that appeal to a particular pollinator. These traits often help pollinators locate flowers and the resources (e.g. pollen or nectar) that the flowers have to offer.
Syndromes include bloom color, the presence of nectar guides, scents, nectar, pollen, and flower shapes. We can use these traits to predict what pollinators might be attracted to certain flowers or we can use these tools to guide us to pick the right plant for the right pollinator!
Bees, for example, are most attracted to flowers that have white, yellow, blue, or ultra-violet blooms.
Blue Flax (Linum lewissii)
Male long-horned bee on a white bindweed flower
Orange bumblebee (Bombus sp.) on a sunflower
Pollinator Syndromes for Bees & Butterflies
Table adapted from the North American Pollinator Protection Campaign
Trait
Bees
Butterflies
Color
White, yellow, blue, UV
Red, purple
Nectar Guides
Present
Present
Odor
Fresh, mild, pleasant
Faint but fresh
Nectar
Usually present
Ample, deeply hidden
Pollen
Limited; often sticky or scented
Limited
Flower Shape
Shallow; with landing platform, tubular
Narrow tube with long spur; wide landing pad
What are nectar guides?
Nectar guides are visual cues, such as patterns or darker colors in the center of a flower, that lead pollinators to nectar or pollen. These cues are beneficial to plants and their pollinators because they can reduce flower handling time, which allows bees to visit more flowers and transfer more pollen in a shorter amount of time.
Northern Blue Flag Iris (Iris versicolor).
The petals (yellow arrow) and sepals (red arrow) both have dark purple nectar guides. The yellow portion of the sepals may also be a nectar guide!
Image courtesy of Mike LeValley and the Isabella Conservation District Environmental Education Program
While the iris’s nectar guides are visible to humans and their pollinators, this is not always the case. Some flowers have nectar guides only visible in ultra-violet light. The video below shows how different flowers look to us (visible light), and simulates what the flowers look like to butterflies (red, green blue, and UV) and to bees (green, blue, UV).
What about pinks and purples?
Red-flowering currant (Ribes sanguineum)
It’s not uncommon to see bees visiting flowers that are colors outside of their typical pollinator syndromes. In the spring in Oregon, we see bees visiting red-flowering currants, many pink and magenta rhododendrons, plum blossoms, and cherry blossoms. Lavender, catnip, and other mint-family plants too are common on pollinator planting lists, but tend to have purple flowers.
Pollinator syndromes can help us understand these anomalies. These flowers may appear differently in ultraviolet light or may have strong nectar guides that encourage bees to visit them, despite how they look to us. Alternatively, these flowers might have rich reserves of pollen and nectar that draw bee visits.
How else do we know if a flower is a good choice for bees?
Many people have developed plant lists based on personal observations, so there are many pollinator plant lists available to choose plants from. Many nurseries include pollinator attraction information with their planting guidelines too. While these are often based on anecdotal evidence, many researchers (including Aaron and I) are working to provide empirical evidence for plant selections.
To find native plants to attract bees and other pollinators, I recommend starting your plant selections by checking out your local NRCS Plant Materials program.
Over the past few months, I have shared data on bees and other insects that we have collected from Portland-area gardens. For every garden insect we study (except for butterflies, which can be identified to species by sight), we use lethal collection methods. This is because most insects can only be identified to species after close examination under the microscope. In fact, some insects require dissection before we can get them to species.
Bombus sitkensis male, with abdomen dissected, in order to make a species-level identification.
It seems odd that we kill bees in order to help understand how we can build gardens that can help to conserve bees. By collecting and killing bees and other insects, what role were we playing in promoting insect decline? How do projects, such as our own as well as the Oregon Bee Atlas, factor into bee declines?
That’s an excellent question, and one that we often ask ourselves. When we collect bees, we work to make sure that we are not needlessly causing harm. For example, our pan traps are good for collecting small bees, but are not good at collecting larger bees, including reproductive queens. When we hand-collect bees, we avoid taking queen bees. In fact, of the 2,716 bees that we collected in 2017-2019, only three were queens. We limited our sampling frequency to three times per year, and limited our sampling effort to 10 minutes of hand-collecting time and six pan traps, per garden. Even with these precautions, we are still faced with the question: does our research, or the research of others who collect and kill insects, harm the very species we are trying to conserve?
Water pan traps, used to collect garden bees and other small, flying insects. Insects are attracted to the color. When they land in the soapy water, they break the surface tension, drown, and die.
To address this question, I turn to the scientific literature. Gezon and colleagues set up an experiment to see whether lethal sampling for bees using pan traps and netting (the same methods we use in our research) has negative effects on bee abundance or bee diversity. For five years, they sampled nine sites every two weeks during the flowering season. They compared bee abundance and bee diversity in these repeatedly-sampled sites, to metrics from 17 comparable sites that were only sampled once. They found no significant difference in bee capture rate, bee species richness, or bee abundance between sites that were sampled repeatedly versus those that were sampled once. When they partitioned bees according to nesting habit (e.g. cavity, soil, wood, etc.), social structure (e.g. eusocial or not), and body size (e.g. small, medium, and large bees) they also found no significant differences in bee capture rates of single-sample versus repeat-sampled sites. They did catch more pollen specialists in repeated-sample sites than in single sample sites. However, the magnitude of the effect was relatively small, and did not represent a large change in catch rate between single-sample versus repeat-sampled sites. I suspect that the authors caught more pollen specialists at their repeat-sampled sites, because pollen specialists are fairly rare in time and in space. They drastically increased their odds of intercepting a pollen specialist on their repeatedly-sampled sites.
Gezon and colleagues suggest a few hypotheses
that could explain why increased sampling effort had no significant effect on
bee abundance or diversity. First, they suggest that reducing bee populations
by sampling could benefit the bees that remain, by reducing competition for
limited resources. If this is the case, bee populations can compensate for some
losses due to sampling, by increasing reproduction in the bees that remain
behind. Second, they note that if bees were sampled after they have mated and
laid eggs, the overall impact of removing a bee from via sampling will be
fairly small. Finally, they note that most bees are solitary, and that most
solitary bees have short flight seasons. In this case, sampling every two weeks
may not result in bee declines, if researchers are effectively collecting a new
species during each sampling event.
I can breathe a bit easier. The data suggests that our research is not immediately responsible for documented bee declines. Still, I know that I can personally do more to help protect bees in my own garden. Even though our lab group studies native plants, I have not yet planted Aster subspicatus (Douglas’ Aster) in my own garden. This will be my mission for 2020: to find and plant this gorgeous perennial at home. In 2018 and 2019, it bloomed from mid June through mid November at our study plots in Aurora, OR, with peak bloom (75% or more of the plant in bloom) lasting one month! And, from 2017-2019, it was always a top five plant for native bee abundance. I give this Pacific Northwest native plant my highest recommendation for home gardens! There are plants that attract more native bees, such as Phacelia heterophylla. But, no other plant that we studied offers the triple threat of beauty, bees, and longevity.
Douglas’ aster (Aster subspicatus) is currently my favorite garden plant for bees.
For my dissertation research, I am studying which native Willamette Valley wildflowers are most visited by pollinators and natural enemies for use in home gardens and urban landscaping. I’ve previously shared preliminary results from my field study on our blog, namely pollinator abundance and richness. For a refresher, here are summaries from 2017, 2018, and 2019.
Initial survey
Determining which of these flowers are most attractive to insects is only half of the equation — I also want flowers that are attractive to gardeners. To investigate this I developed two surveys — thanks to anyone reading who took them! The first simply asked gardeners to rank the aesthetic appeal of my study plants, as well as how likely they would be to utilize them in their home gardens. This allowed me to get a baseline understanding of how appealing these flowers are for use in home gardens and landscaping.
As you can see in the figure below, many of the plants most visited by bees (highlighted in orange) were the least attractive to gardeners (Fig. 1), while plants gardeners liked the most (e.g. Iris, Columbine) were hardly visited by bees. However, its notable that many of these native wildflowers ranked around a four on a 1-5 scale, showing that these flowers do have a high potential appeal for use in landscaping!
Figure 1: Gardener ranked aesthetic appeal of study flowers on a scale of 1-5. Orange bars note plants that were consistently highly visited pollinator plants. N=587
Follow-up survey
The follow-up survey consisted of a subset of ten flowers most visited by bees, and again asked respondents to rank the aesthetic appeal and likelihood of planting for each of these flower species. Then, they were shown facts about and images of bees that visit each flower species, and asked whether they viewed each plant species more favorably, less favorably, or the same. Finally, they were asked to re-rank how attractive they found the flower species and how likely they would be to use the species in their garden, both on a scale of 1-5.
Gardener acceptance
This second survey showed a remarkable increase in gardener acceptance of pollinator friendly native plants after being educated on plant-pollinator associations. Over 80% of respondents stated that they viewed Clarkia amoena as more attractive after gain, and over 60% of respondents viewed Phacelia heterophylla, Madia elegans, and Gilia capitata as more attractive (Fig. 2).
Figure 2: Percent of respondents viewing flower species as more attractive after learning about pollinator associations. N=184.
Likelihood of planting
After learning about the benefits these flowers provide to pollinators, gardeners were also more likely to plant all ten flower species (Fig. 3). Notably, they were 40% more likely to plant Phacelia heterophylla, (a species that ranked as the least aesthetically appealing overall in the first survey). As a whole, they were also over 20% more likely to plant Solidago canadensis, Clarkia amoena. Similar increases were also observed in likelihood of planting Oreganum vulgare and Nepeta cataria. Many of the plants that showed a smaller percent change are species that started out with a higher aesthetic appeal (e.g. Gillia capitata, Lavendula intermedia, Aster subspicatus), meaning gardeners were already very likely to include these plants in their home garden before learning about the ecological benefits they provide.
Figure 3: Percent change in respondent’s likelihood of planting each top pollinator flower after learning about the pollinators associated with each. N=184
Ecological beauty
What does this all mean? This suggests that although native plants are frequently denounced as being less attractive than showy garden species, many home gardeners are still willing to use native flowers in their landscaping. Additionally, this lends credence to the concept of “ecological beauty” – that many gardeners are willing to utilize plants that will increase the habitat value and wildlife diversity in their yards.
















































{kind=link}