I have been so grateful for the opportunity to assist Jen Hayes with her nativar research project. For the past year and a half, I have learned so much in the world of plant and pollinator relationships. One of the most valuable things I have learned, which I use every day in my own garden, is how to quickly identify bees. In the field, we observe each flower plot for five minutes and record the different things that visit. Being able to identify a fly from a bee, or a bee from a wasp is very important in order to obtain accurate data. I wanted to share the process we use to quickly identify bees in the field and hopefully answer some questions you may have had about what was buzzing in your garden.
A small black Halictidae bee examining the Douglas aster cultivar, Sauvie Snow. The photo was taken by Tyler Spofford at Oak Creek Center for Urban Horticulture in Corvallis, OR.
Once the sun has risen and the chill of the morning has left, bees begin their foraging routine. Male bumblebees, out foraging too late, emerge from the layers of Zinnia flowers. Hundreds of bees possibly pass through your garden in a single day, from flower to flower, collecting pollen and nectar. But what are they? Are they native? Are they helpful in the garden? These are all questions I have whenever I see something buzzing on the mint, exploring the flower patch, or pollinating the tomatoes.
The first things I always look for are the antennae and the number of wings. The flies that are most often confused with bees have short, almost non-existent antennae, whereas bees will have noticeable, segmented antennae. The wings are also something to look: flies only have one pair of wings and bees have two, the forewings and hindwings. Be sure to look closely! There are flies known as hoverflies or syrphid flies that have incredible mimicry adaptations. Look at these two insects on this butterfly bush. Although almost identical, you can see the top insect does not have noticeable antennae. That’s because it is a honey bee mimic!
The bottom insect is a honey bee and the top insect is a syrphid fly that is a mimic of honey bees. The photo was taken by Tyler Sato Spofford at Lewis Brown Horticulture Farm in Corvallis, OR.
If you have established that it is a bee and not a fly, there are other things to look for to identify the bee to a more specific taxonomic level. The coloration of the bee could help if it is green or red, but there are many bees that have different variations of black and white. What I like to look for next is the pollen baskets, also known as the corbiculae. Megachilidae bees carry the pollen on the underside of their abdomen, like bright yellow furry bellies. Another distinguishing factor for Megachilidae bees is how they fly. The abdomen of the bee will usually curl upwards while in flight. Mellisodes bees carry the pollen on their hind legs but the baskets are dramatically bigger than honeybees or bumbles. We like to think of them as pollen pants! Mellisodes bees are also known as long-horn bees because of their disproportionately long antennae.
A mellisodes collecting pollen. Look at those pollen pants!
If the bee does not have any special coloration or noticeably different corbiculae, it could be one of many other genera we have in Oregon. Halictidae bees range in size from the tip of a pen to the size of a penny. They are usually black or black and white and are VERY difficult to distinguish in the field. There are details we have to look for in the lab such as the number of “panels” in the wings or if they have one versus two sub-antennal sutures.
The other bees we see while doing research include wasps, honeybees, and bumblebees. There are so many variations of Bombus here in Oregon it is almost like a scavenger hunt. Because of all the color and striping variations, we use the PNW Bumblebee Atlas to help us identify species in the field.
Our second field season studying pollinator visitation to Oregon native plants and native cultivars spanned from April to late September of 2021, although if Douglas Aster had any say in the matter, we would likely still be sampling. The densely blooming Symphyotrichum subspicatum continued to produce a smattering of new flowers through November of last year, and we predict it will do the same this year, too!
Our field crew this summer included Tyler, Svea, Mallory and I. Together, we sampled on 33 different dates across the growing season, allowing us to collect around 2000 physical pollinator specimens, and observe 6,225 unique interactions between pollinators and our study plants! This season we conducted floral trait measurements (including the dimensions of flowers), took multispectral photos, and additionally collected pollen from a subset of our study plants.
From left to right: Mallory vacuum-sampling off of Douglas Aster 'Sauvie Snow', Tyler shaking a farewell-to-spring flower to get pollen off of it, and Svea photographing Baby Blue Eyes 'Penny Black'.
This year, we introduced a third cultivar for California poppy (Eschscholzia californica ‘Purple Gleam’), yarrow (Achillea millefolium ‘Moonshine’), and farewell-to-spring (Clarkia amoena ‘Scarlet’). The new cultivars were established in the spring, which resulted in a late bloom for the annuals, so we expect to see them blooming during their typical period in 2022. The Achillea ‘Moonshine’ replaced Achillea ‘Salmon Beauty’ in being the most abundant yarrow cultivar; it began blooming almost immediately as it was planted into our field site and is still continuing to push out blooms through October alongside the Douglas Asters.
The plant groups in our study: the larger circles with orange text are the native plants, and the smaller circles and turquoise text are the cultivars. The top row contain the perennials yarrow, western red columbine, great camas, and Douglas aster. The bottom row shows the three annuals farewell-to-spring, California poppy, and baby blue eyes.
In addition to watching new plants bloom in the study garden, we had the opportunity to observe many incredible pollinators in the field this summer. We saw a hummingbird visit the Western Red Columbine, we tried to capture videos of leaf-cutter bees snipping little petal pieces off of farewell-to-spring, and at a neighboring plot we observed a male wool-carder bee section off an entire patch of Salvia for a female bee.
On the left: Farewell-to-spring 'Scarlet' with crescents cut out of the petals by leafcutter bees. Top right: A female wool-carder bee (Anthidium manicatum) collecting trichomes from Yarrow 'Calistoga'. Middle right: A leafcutter bee with a piece of petal from Farewell-to-spring 'Dwarf White'. Bottom right: A leaf cutter bee removing a piece of petal from Farewell-to-spring 'Aurora'.
We were also able to take a couple educational field trips this field season in order to learn about pollinator studies ongoing outside of Oak Creek. In June, we went up to the North Willamette Research and Extension Center in Aurora, OR to listen to three talks about pollinators at the Blueberry Field Day. We learned how to score the productivity of honeybee hives, how to properly don a the top of a bee suit, about blueberry’s best pollinators, and blueberry research projects at the University of Washington.
In August, we made a trip to Bend for a different kind of study… an artistic one! We travelled to the High Desert Museum in order to visit Jasna Guy and Lincoln Best’s exhibit “In Time’s Hum…”. Jasna is a brilliant artist inspired by pollinators, which translates into the subject of her pieces as well as her artistic media. Many of her pieces are made using encaustic (a method of painting using wax, bee’s wax in her case!), dipped directly into bee’s wax, or involve pollinators in some other format, including her color study of pollen, which attempts to replicate the colors of fresh pollen as well as the colors after bees have mixed them with nectar. In the center of exhibit were two cases filled with bees collected and identified by Linc, surrounding some of the dried plant specimens they forage on.
These field trips were a wonderful way to see what other pollinator work is happening in our broader community and to inspire future studies. It was especially exciting to see how Jasna and Linc combined art and science with their exhibit, which is something many of us in the Garden Ecology Lab are interested in.
1. Mallory, Svea, and Jen at the blueberry Field Day. 2. Svea, Jen, Mallory, and Tyler at the High Desert Museum. 3. A panorama of the "... In Time's Hum ... " exhibit. 4-5. Art on the outside of the exhibit. 6. A snapshot of two pollen samples from Jasna Guy's pollen color study.
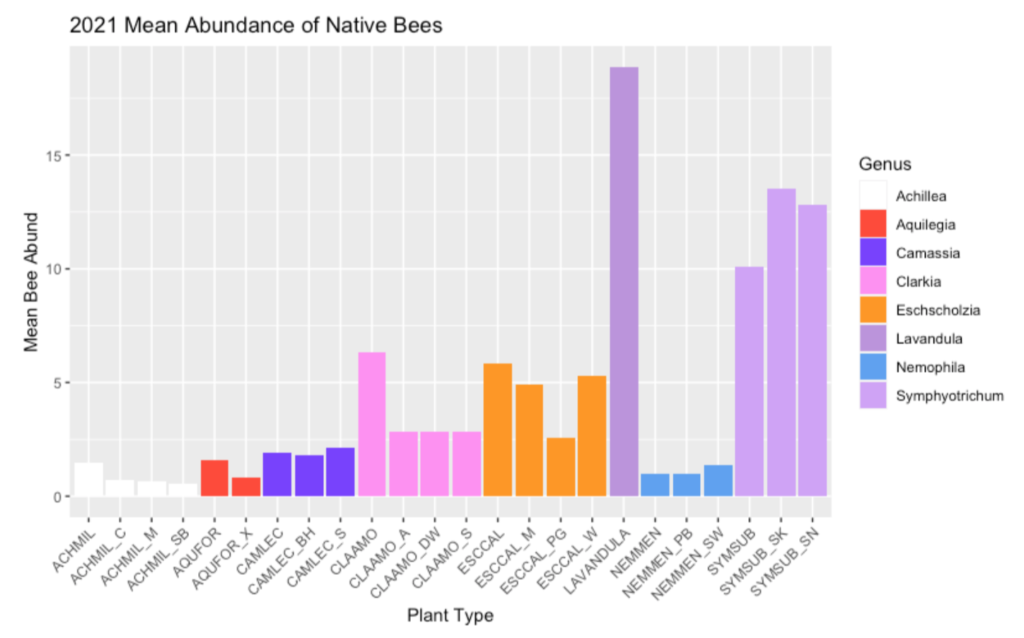
While we cannot make conclusions until we complete our final field season, we are excited to report some of the variation in visitation between native plants and native cultivars that we have observed in our first two field seasons. In the first field season, our observations of native bees foraging on the study plants revealed three plant groups to have variable amounts of visitation. Yarrow, farewell-to-spring, and California poppy all had at least one cultivar that received substantially less native bee visits than the native type. In our second year, all three of farewell-to-spring’s cultivars received less visitation than the native Clarkia amoena. Poppy had only one cultivar with less native bee activity than the native (Purple Gleam), and in the case of Douglas Aster, both of the cultivars actually had more visitation by native bees than the native.
Figure 1: Average Abundance of Foraging Native Bees during 5-Min Observations in 2021. Individual plants are color-coded by genus. The naming scheme combines the first three letters of the genus and specific epithet; cultivars are denoted by an underscore and a 1-2 letter code to identify them. For example, AQUFOR is the native Aquilegia formosa, and AQUFOR_XT is Aquilegia x ‘XeraTones’.
Today, Tyler successfully defended his undergraduate research thesis, entitled ‘Invest in Vegetables: A Cost and Benefit Analysis of Container Grown Roma Tomatoes (Solanum lycopersicum cv. ‘Roma’) and Italian Basil (Ocimum basilicum cv. ‘Italian’)‘.
Tyler Spofford, in the middle of his containerized garden research plot, in the summer of 2020.
His research was inspired by the rush to vegetable gardening, that many households made during the start of the COVID-19 pandemic. Research has shown that there are many benefits to vegetable gardening, including social, emotional, physical, and financial. However, those in rental housing, or otherwise without easy access to land, were largely locked out of accessing these benefits.
Although previous research has shown that in-ground and raised bed vegetable gardening can yield positive economic benefits, to date, no studies (that we know of) have quantified the financial costs and benefits of growing vegetables in containers. Tyler thus set up a system of 5-gallon and 3-gallon bucket gardens, planted with Roma tomatoes, or Roma tomatoes plus Italian basil. He kept careful track of the cost of materials, and the time he spent gardening. He also kept careful track of the harvest he pulled off of each container.
Over the course of his study, he successfully learned about and fought back Septoria leaf spot, and blossom end rot. We learned that Roma tomatoes, in particular, are susceptible to blossom end rot. On top of these horticultural plant problems, Tyler’s research was abruptly halted by the late summer wildfires of 2020, that made air quality unsafe for him and others to continue their work, outdoors.
Despite these challenges, he was able to glean enough data from his project, to share some interesting findings:
None of the containers netted a positive economic benefit, in the first year of gardening, largely because the cost of materials outweighed the financial benefits of the harvest.
If the project were continued into year two, he projects that he would have had a positive financial outcome for the tomatoes grown with basil, in the 5-gallon containers.
Across the course of the season, he only spent 30 minutes tending to each container. Because he had few garden maintenance tasks, the time invested in container gardens was minimal. This is an important finding, for folks who may shy away from gardening because of lack of time.
As expected, the 5-gallon containers yielded more than the 3-gallon tomatoes. The 3-gallon containers stunted plant growth too much, to recommend them as a viable container gardening system. [As a side note, we were given the 3-gallon containers, for free, which is why we included them in the study.]
The fair market value of Roma tomatoes was fairly low (~$1.00 per pound). Thus, the net economic benefit of growing Roma tomatoes was also low. Basil, on the other hand, was a high value specialty crop that helped to raise the overall economic value of crops harvested from the buckets.
If you are interested in seeing Tyler’s thesis defense presentation (~30 minutes), you can do so, at the link below.
Tyler will be graduating in a few days, with a degree in BioResource Research from OSU! He’s worked in our lab group for two years, and has been an absolute joy to learn and work with. We wish him the very best on the next adventures that await him.
Flowers and bees have one of the most well-known symbiotic relationships ever formed. Flowers rely on bees for pollination, and bees rely on flowers for nectar and pollen. It is generally understood that flowers act as advertisements to attract bees. However, less is known about what exactly bees are seeing and how that can change once humans get involved. This project is focused on the changes that can arise after a plant is cultivated, and how these changes can affect pollinator preference of a flower.
While changes made by breeders might not seem all that drastic to our eyes, we have little idea if that is the case for bees. Often breeders will change flowers for aesthetic purposes. This can have unknown consequences. These changes might not seem like such a big issue since the flowers are still colorful. However, bee vision is very different from humans, with bees having the ability to see into the UV spectrum. This means that while we might think we are only changing the bloom size or the color, we could also be unintentionally changing UV messaging visible only to the bees.
The purpose of this study is to use UV photography to explore these invisible differences between the native and cultivar. We also want to determine if the differences have a tangible impact on pollinator preference. This study is ongoing, but the images so far have shown a few native/cultivar sets that have a marked difference in UV markers between native and cultivars. While the study has only just started, our excitement and curiosity have not abated. This is an entirely new foray into pollinator relationships and mechanisms and could open up the world of bees and flowers in a brand new way.
An example of a UV photo of a nemophila flower, with a UV marking in the center, highlighted in blue
In this post, I cover the 2009 paper, “Impact of native plants on bird and butterfly biodiversity in suburban landscapes,” by Karin Burghardt, et al.[i]
*************
This study was published shortly after the first edition of Doug Tallamy’s book, Bringing Nature Home: How Native Plants Sustain Wildlife in Our Gardens.[ii] After decades of studying host plant records of butterfly and moth species, Tallamy was convinced that native plants were critically important to wildlife conservation. About half of all insects are herbivores, and about 70 percent of all herbivores are specialists that are only capable of feeding on a narrow range of plants (see Schoonhoven et al. 2005, Chapter 2, pages 5-9). Specialist insects have developed, over time, the ability to feed on plants that have physical or chemical deterrents that keep generalist insects at bay. Although specialist insects can feed on plants that can be toxic to other insects, they can’t easily switch to feed on novel, non-native plants.
Burghardt and Tallamy’s Study of Native Plants and Caterpillars
Tallamy was Karin Burghardt’s master’s degree advisor and one of her co-authors on the 2009 paper. In their study, they selected six pairs of suburban gardens in central Pennsylvania. Gardens were paired by size and proximity. One garden in each pair featured the conventional landscaping for this region: large lawns, Asian shrubs, Asian understory trees, and native canopy trees. The other garden was landscaped with native ornamentals at each vegetative layer: grasses, shrubs, understory trees, and canopy trees.
They counted the number of caterpillars at 12 points within each garden. Since caterpillars are herbivores, and most insect herbivores are specialists that do best on native plants, they hypothesized that they would find more caterpillars in the native plant gardens. Indeed, this is what they found. Caterpillar abundance was four times greater, and caterpillar species diversity was three times greater, in the native gardens versus the conventional gardens. In addition, Burghardt found that native plant gardens harbored more birds. In fact, native plant gardens had 55 percent more birds and 73 percent more bird species, compared to conventional gardens!
This study demonstrated that gardeners’ choices matter and can clearly influence ecological food chains. Food chains are organized into what are known as trophic levels. Organisms on the same trophic level share the same ecological function and nutritional distance from the sun. Photosynthetic plants are on the first trophic level. Herbivores, or organisms that eat plants, are on the second trophic level. Organisms that eat herbivores, such as birds, are on the third trophic level.
Burghardt and Tallamy demonstrated that what you decide to plant in your garden not only determines the structure of the first trophic level but can also cascade up to affect the second and third trophic levels. As an entomologist, I was not surprised that gardeners’ plant selections could influence the second trophic level. However, I was blown away that these decisions could cascade up to strongly influence the third trophic level.
Garden Ecology Lab Studies of Native Plants and Bees
In the Oregon State University Garden Ecology Lab, we study relationships between native garden plants and native bees. To be honest, I did not expect that native bees would prefer native plants. Whereas the leaves and other vegetative parts of a plant include an array of chemical and physical defenses to protect the plant from insect herbivores, flowers have few such defenses. In fact, flowers function to attract pollinators to a plant.
Thus I was somewhat surprised when Ph.D. student, Aaron Anderson, found that most of the native plants in his study attracted more native bees and more species of native bees than his non-native comparison plants. For example, non-native lavender ‘Grosso’ attracted a large number of bees, but most of these bees were non-native honey bees. By and large, the native plants were better for bee abundance and bee diversity, compared to the non-native comparison plants. In particular, Globe Gilia, Farewell to Spring, Oregon Sunshine, Douglas Aster, and California Poppy were all particularly attractive to native, wild bees in Aaron’s study.
Why might native bees prefer native plants, when flowers don’t have the same chemical and physical deterrents that herbivores must contend with? One hypothesis is that the nectar and pollen in native plants might provide better nutrition to native bees. Another hypothesis is that pollinators are keenly tied into the visual display of native plants. Flower color, size, shape, and ultraviolet markings are all important signals that flowers use to attract the attention of various pollinators. If there are changes in any aspect of this visual display, pollinators may no longer recognize a flowering plant as a good source of pollen or nectar.
Another OSU Ph.D. student, Jen Hayes, is trying to understand why native plants seem to be preferred by native pollinators. As part of her Ph.D. work in the Garden Ecology Lab, Jen is collaborating with an OSU photography student, Svea Bruslind. Svea uses different filters to take photographs of native plants and native cultivars in visible light, ultraviolet light, and in “bee vision” light. We are just getting started on this study, but look forward to reporting our findings in the near future.
[i]Burghardt et al. 2009. Impact of native plants on bird and butterfly biodiversity in suburban landscapes. Conservation Biology 23:219–224.
[ii] Updated and expanded version published as Bringing Nature Home: How You Can Sustain Wildlife with Natives Plants.
Your garden soil contains millions to billions of individual microorganisms, including bacteria, fungi, viruses, and archaea, representing tens of thousands of different microbial species. Humans evolved for millenia in the presence of these environmental microbes associated with vegetation, soil, water, and wildlife. Our immune systems are not only adapted to coexist with the majority of these microbes, but may even require that interaction to function properly. Emerging scientific evidence suggests that exposure to soil microbes trains the immune system, reduces inflammation, and improves mental health (Rook, 2013). For example, the common soil bacterium Mycobacterium vaccae has been found to have positive impacts on stress tolerance and mental health (Matthews and Jenks, 2013), while other research has shown that children exposed to greater microbial diversity, such as that encountered in farming environments, tend to have lower prevalence of autoimmune disorders, including allergies and asthma, than their urban counterparts (Hanski et al., 2012).
The primary goal of the Garden(er) Microbiome Project was to understand how much microbial transfer from soil to skin occurs during gardening activities, what types of microorganisms are transferred, and how long they can persist on the skin. We are also interested in exploring how soil microbial communities vary with different management practices (e.g., organic vs. conventional) and geographic locations, as we know that microbes play critical roles in soil nutrient cycling, carbon sequestration, pollutant degradation, and, of course, crop health.
To accomplish this study, we recruited 40 gardeners to collect microbial samples from their garden soil and from the surface of their skin (hands). All samples were collected in July–September, 2020, and were equally distributed between the Willamette Valley and High Desert regions, as well as between self-reported organic and non-organic management practices. Each volunteer was asked to collect soil samples from three different garden beds and skin microbiome swabs before, after, ~12 hours after, and 24 hours after gardening (Figure 1). To identify bacterial taxa (different types of bacteria) present in the samples, we used Earth Microbiome Project protocols to sequence the V4 region of the bacterial 16S rRNA gene.
Figure 1. Volunteers collected samples from their garden soil and hand surface microbiome.
Preliminary results
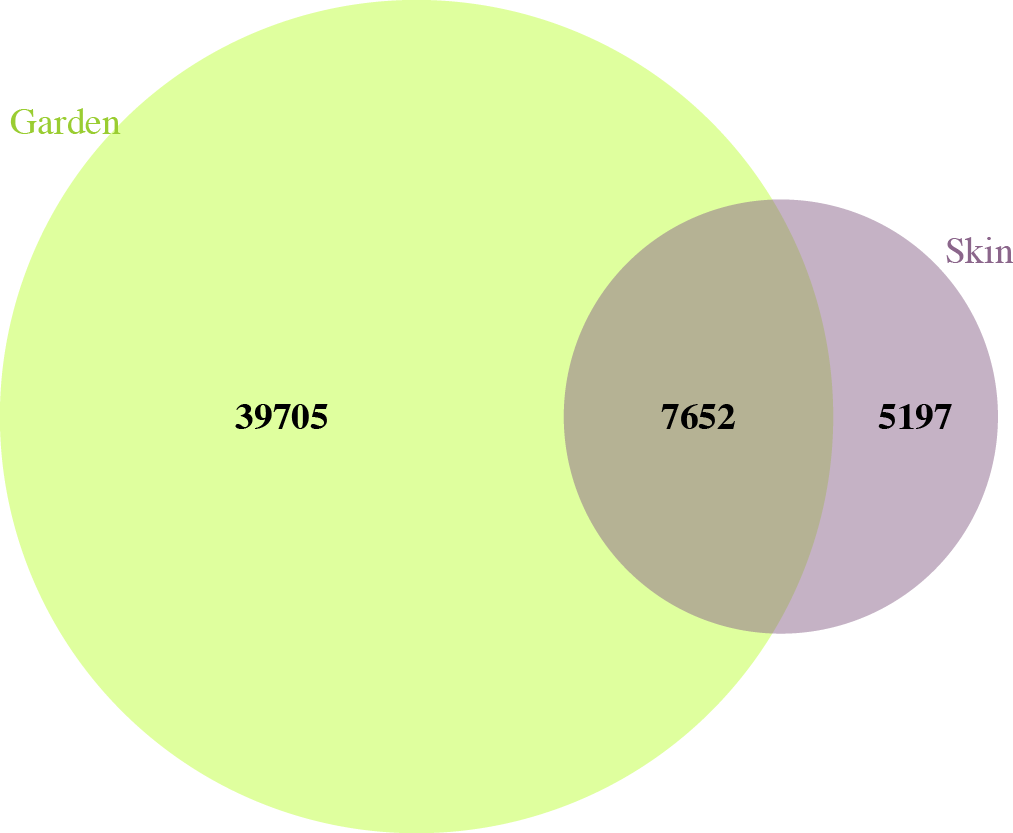
In garden soil samples, we observed over 8.5 million individual bacteria, representing about 45,000 different bacterial species. In skin microbiome samples, we observed over 6 million individual bacteria, representing almost 13,000 different bacterial species. Of all these bacterial species, there were just over 7,500 that were shared between garden soils and gardeners’ skin microbiomes over the course of the study (Figure 2).
Figure 2. Unique and shared bacterial taxa found in garden soil and skin surface samples.
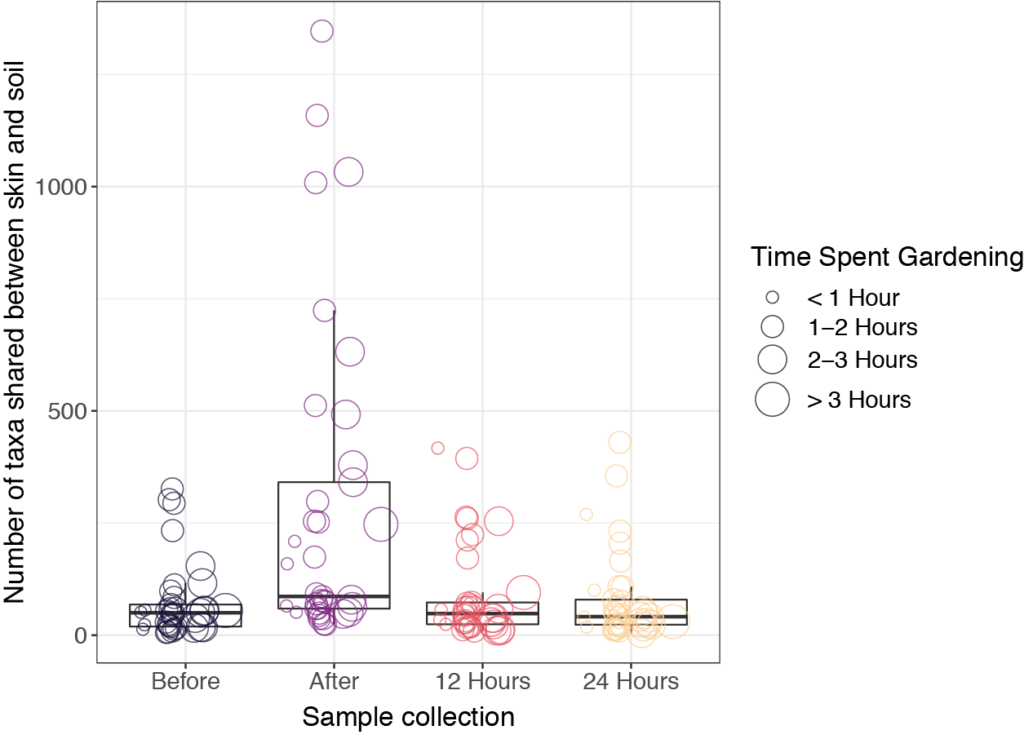
Our initial hypothesis was that skin microbiome samples would be more similar to soil samples immediately after gardening, due to microbial transfer from soil to skin during direct contact. We also expected that the skin microbiome would return to baseline (before gardening) after a period of time, depending on individual behaviors, such as washing hands and bathing. It turned out that soil microbial communities were very different than those found on skin. Interestingly, skin microbiome samples tended to be dominated by a small number of taxa, though they were not always the same taxa at different sampling times. For many of the study participants, we did indeed see an increase in shared taxa for the skin samples collected immediately after gardening (Figure 3). However, soil microbes were generally transient on the skin and were no longer present after 12 hours. We note that the COVID-19 pandemic may have influenced hand-washing behaviors and use of hand sanitizers, which could have had additional unexpected impacts on the skin microbiome.
Figure 3. Bacterial transfer and persistence on skin after gardening.
Though it was beyond the scope of the project to describe life history details about every type of bacteria that was found, we did investigate a handful of taxa that were highly abundant in many samples. In garden soil, many of the most abundant bacteria belonged to the genus Pseudomonas. In a recent paper, Sah and Singh (2016) state, “The genus Pseudomonas encompasses arguably one of the most complex, diverse, and ecologically significant group of bacteria on the planet. Members of the genus are found in large numbers in all the major natural environments (terrestrial, freshwater, and marine) and also form intimate associations with plants and animals.” Importantly for gardens, several species of Pseudomonas are able to promote plant growth, while others are well-known plant pathogens. Members of the genus Sphingomonas were also common soil inhabitants found in this study. Sphingomonads are broadly distributed in the environment, including soil, water, air, and plant leaves. Only one species of Sphingomonas is known to cause disease in humans, typically in hospital-acquired infections (Balkwill et al., 2006). A third genus of interest from garden soils was Streptomyces. This is a large genus with over 500 members that are ubiquitous in soils. They are known to form symbiotic relationships with plants and animals, and they are responsible for the production of over 2/3 of all known antibiotics (Antoraz et al., 2015). Streptomyces also produce the chemical compound geosmin, which gives soil its earthy smell (Seipke et al., 2012).
The composition of skin microbiome samples in this study varied wildly from individual to individual, and sometimes even for the same individual at different time points. Among the most abundant taxa we found were members of the genera Pantoea, Acinetobacter, Bacillus, and Klebsiella, as well as Pseudomonas, which was described above. Generally speaking, these are very diverse genera and are widespread in many environments, including soil and human skin. Some Pantoea species produce antimicrobial compounds that can help control fire blight in fruit trees (Walterson and Stavrinides, 2015). The genus Acinetobacter contains two species of interest for health reasons—A. baumannii is typically found in wet environments and is a notable opportunistic pathogen associated with hospital-acquired infections (Howard et al., 2012), whereas exposure to environmental sources of A. lwoffii is thought to protect against development of allergies, although it can also cause infection in immunocompromised individuals (Debarry et al., 2007). Members of the genus Bacillus have been explored for potential probiotics (Elshaghabee et al., 2017), and the genus Klebsiella is somewhat notorious for its human pathogenic members. However, Klebsiella, Pantoea, and several other members of the Enterobacteriaceae family have highly similar DNA sequences in the region that we targeted, so these composition results should be interpreted cautiously.
Conclusion
This study represents one of the very first investigations of garden soil microbiomes and, to our knowledge, the only one that explores the ability of soil microbes to transfer and persist on human skin after typical gardening activities. Overall, we found that garden soils tend to have far greater bacterial diversity than skin microbiome samples. Bacterial community composition was largely similar across different garden beds, whereas skin microbiome composition varied dramatically. Some soil microbes appeared to transfer onto skin during direct contact with soil, but they were generally gone within 12 hours, suggesting a low ability to permanently colonize skin. However, a daily gardening routine with repeated and extended contact with soil likely reinoculates the skin such that soil microbes are like a regular visitor during the growing season.
The specific ecological role of most microbes, both in soil and on skin, is a relatively new area of investigation garnering intense interest. However, few, if any, concrete recommendations are currently available to guide actions towards improving plant and human health. A primary goal of this study is to gather baseline data for future studies, which are needed to further explore the impact of daily soil contact over longer time periods (e.g. entire growing season), how changes in gardeners’ skin microbiomes compare with non-gardeners, and whether consumption of fresh garden produce affects the gut microbiome.
My name is Mallory Mead, and I am new to the Garden Ecology Lab! I am an undergrad studying Horticulture and minoring in Entomology, and I started a few weeks ago as an assistant to Jen Hayes on her study of pollinator attraction to native plants and nativars.
I enrolled in Oregon State’s URSA Engage program, which gives undergrads a taste of research experience in the Winter and Spring of their first year, and joined a project studying how mason bees might be impacted by climate change with Dr. Jim Rivers of the department of Forest Ecosystems and Society. The study seeks to examine the effects of warming temperatures on mason bee behavior and the development of brood.
The Western US’s native species of mason bee, the Blue Orchard Bee (BOB) is known to be an excellent orchard pollinator. On many orchard crops they are more efficient at pollination than honey bees on a per individual basis, and so the commercial management of BOBs is being explored as honey bee colonies suffer management challenges and colony losses in recent years.
Mason bees have a short lifespan of 4 to 6 weeks. Emerging in the early spring, males die shortly after mating, while females build nests in holes in wood or reeds. They forage for pollen and nectar to form provision masses in which they lay their eggs. They also collect mud to form partitions between each provision mass and to cap the nest once it is full. Their offspring will feed on the provisions and metamorphose into cocooned adults to overwinter in their cells and emerge the following spring.
To ensure the bees had ample nutrient resources, the project was conducted next to the organic cherry orchard at OSU’s Lewis Brown Farm. Before the cherries bloomed, 6 nest structures were designed and constructed by Jim, Ron Spendal (a mason bee house conisuerrier) and Aaron Moore of Revolution Robotics.
Nest structures, solar panels, and camcorders at Lewis Brown Farm.
Each structure has 3 shelves with 16 nest holes each, lined with paper straws so that the nests can be easily removed and examined. The structures are solar powered, and each shelf is heated to a different increment above the ambient temperature i.e. + 0°C , + 2°C, + 4°C, + 6°C, + 8°C, + 10 °C, and + 12°C. These differentials represent many potential warming outcomes of climate change.
Nest Structure Number 2 with labelled component parts. A. The Electronics control box. B. Cocoon-release box. C. Shelves sandwiched by heating pads, and lined with paper nesting straws
Our Hypotheses
We predicted that female mason bees will select the warmer nests first, and that females will leave nests earlier in the morning to begin foraging because they will reach the critical internal temperature necessary for flight sooner.
If heated bees have a greater window of foraging time, then we predict they’ll be able to construct nests at a faster rate in the warmer nests.
With greater nest construction will come a greater production of offspring from the bees in the warmed nests.
But…
In terms of offspring quality, we predict that offspring of heated nests will emerge as weak individuals and mortality will be the highest for the heated brood.
…and we are pretty confident about this last prediction.
Insects are poikilothermic meaning their internal temperatures are determined by the environment. Past studies by researchers Bosch and Kemp have reported that mason bees who are overwintered at warm temperatures will “use up their metabolic reserves and are likely to die during the winter”. And a more recent study by researchers at the University of Arizona found that mason bees subjected to heating resulted in reduced body mass, fat content and high mortality of the mason bee offspring.
Data Collection
One of the latest male mason bees to emerge, surrounded by empty cocoons in the release box.
Our mason bees started hatching from cocoons in mid-April and began to colonize the nest structures. I captured video footage of the bees as they emerged in the morning to forage. If bees from heated nest sites emerge earlier, this will support our hypotheses that they reach their critical-for-flight temperature earlier, and get a leg-up on foraging compared to their neighbors.
I also conducted “nest checks” to track the rate of nest construction along with two other research assistants.
In the fall, the nest tubes will be extracted to examine the reproductive output, and in the following spring, offspring will be assessed for rates of mortality, offspring mass, and fat content.
Obstacles
Some of the challenges along the way have included dealing with insect pests. Spiders were easygoing inhabitants of the nest straws, for they only nested in empty straws, so we’d swap them out for a clean one. The earwigs were much more pervasive, and went for the already inhabited nests. As generalist foragers, the earwigs took advantage of provision balls of nectar and pollen that had not yet been sealed off by mud. Once I read that earwigs will indeed eat the mason bee eggs that are laid into the provision masses, I knew it was crucial to remove the earwigs from all nests, but these feisty creatures proved determined to stay. We ordered some tanglefoot, a sticky substance to trap the earwigs on their way up the structure post, and meanwhile I coaxed earwigs out with tiny pieces of grass. Jabbing them repeatedly would eventually provoke them to charge at the blade of grass and fall out from the straw.
Yellowjackets were another opportunistic nester. They’d sneak into the cocoon boxes to build nests, and always gave me a start when opening the tiny boxes. I removed their nests with an extended grabber tool and would destroy them in any way I could. I feel immensely lucky not to have been stung through this process.
The most terrifying surprise during the project was a fat snake that was living in the solar panel battery box. It popped out at me hissing while I conducted a routine check. Alas, I was too spooked to take on this unexpected visitor and let it leave on its own time.
Preliminary Findings & Observations
By mid-May, a pretty clear pattern was emerging. At each structure, the control shelf’s nests (+ 0 °C) were full and capped with mud, while the hottest shelves were almost completely empty. We will analyze nest check data to confirm that these patterns are not just arising by chance, but a study that was released this past April showed another species of mason bee in Poland following the same pattern of nest site preference and selection for cooler nest sites.
The mason bees’ unexpected behavior of avoiding the heated chambers may lead to trouble during the second part of the experiment because this means our sample size for heated offspring has become so tiny, but here it is important to note that this is mason bee project is a pilot study and so the data collected this year will simply influence more specific future research.
these preliminary findings make me think that mason bees have an ingrained sense to avoid warm nests, which might show mason bees’ adaptability in the face of climate change, that is, if they can manage to continue finding cool nests. People managing mason bees find that nests facing the morning sun are the most attractive to the bees, but I wonder how long it will be before temperatures rise and mason bees start avoiding these sunny nests.
Moving Forward
By the end of May, I’d only see a few the mason bees per visit, so the season was clearly coming to an end. I wrapped up data collection and am now spending the summer extracting data from the video footage, and checking up on the bees to ensure they are safe and sound until Fall inspections.
I am wishing the best to both the wild bees in our region and those in our study, as the temperatures skyrocket this week but with this summer’s heat wave, I don’t think we need to simulate climate change; it is right here before us. Even though it is practically inevitable that temperatures will rise to dangerous heights in my generation’s lifetime, there is so much life to be saved, and there is no time to waste.
“Blue Orchard Bee, Osmia lignaria” by SeabrookeLeckie.com is licensed under CC BY-NC-ND 2.0″
According to the staff at Oak Creek and many other gardeners and farmers I’ve had the opportunity to talk to, it appears that though 2020 was a difficult year for humans, it was truly a remarkable year for gophers and other rodents.
From left to right: wild type Great Camas, Camassia leichtlinii, the native cultivar ‘Sacajawea’, and the native cultivar ‘Caerulea Blue Heaven’.
Gophers & Camas
No matter how often a gopher was trapped and removed from Oak Creek last summer, the next week there would always be a mound of freshly turned soil on the grounds, indicating a new gopher had taken its place. While they seemed to enjoy popping up in some of the Organic Gardening Club’s beds, they had an extra fondness for my own experimental garden beds. Fresh gopher-turned soil was most commonly found in any plot growing our native Camassia leichtlinii (Great Camas) and the plots surrounding them.
Bulb size comparisons for the three varieties included in our study.
We planted our 15 camas plots in the fall of 2019. Five plots were planted with the wild type camas species, Camassia leichtlinii (Great Camas). Five more were planted with the C. leichtlinii cultivar ‘Caerulea Blue Heaven’, and the final five were planted with C. leichtlinii ‘Sacajawea’. By the spring of 2020, the camas plots were relatively untouched, aside from some minor grazing by deer on a handful of plots. In April our three camas varieties began blooming in sequence (the native first, followed by ‘Blue Heaven’ and ‘Sacajawea’, respectively), and by mid June they had all gone to seed.
Deer browsing on early spring shoots of C. leichtlinii ‘Sacajawea’.
Though the gopher troubles seemed to really begin in June, there were signs of their activity that we did not heed. In spring of 2020 I was planting a Clarkia amoena cultivar plug. Upon removing some soil to make room for the plant, I found that the soil seemed to drop off into a massive hole beneath the plot I was planting. I shook some soil loose to fill the hole, planted my Clarkia, and moved on. Later in the season, a different Clarkia plant would be found dead, and upon its removal, another tunnel would be found beneath the top layer of soil.
By August, there had been so much gopher activity in our beds that I decided we needed to conduct a damage assessment. I asked Tyler to dig around in a Camas plot that seemed particularly ravaged by the gophers, to see if he could find any of the original 40 bulbs we had planted. His searching returned no bulbs.
Bulb Thieves
I immediately went through each of the 15 camas plots and rated them with a visual assessment of the gopher activity that we would use to determine how many bulbs likely remained in the plots. The levels we decided on were “low/no damage” “Low damage”, “Moderate Damage”, “High Damage” and “Extreme Damage”. Plots with no damage were expected to have all 40 original planted bulbs. On the other end of the spectrum, plots labeled “Extreme” were expected to have no remaining bulbs.
At the end of our field season, we dug out the bulbs from each of the camas plots so we could assess the actual damage, and so we could install fencing to keep all future gophers out. During the bulb dig, we recorded the total number of bulbs found in each plot. In the table below, I have shared the visual damage rating for each plot, the estimated number of bulbs expected to be in the plots, and the actual number of bulbs we found.
Rep #
Bulb Type
Visual Damage Rating
Estimated Remaining Bulbs
Actual Remaining Bulbs
1
Blue Heaven
Low-no
40
2
2
Blue Heaven
Low-no
40
66
3
Blue Heaven
High
10
53
4
Blue Heaven
High
10
66
5
Blue Heaven
High
10
5
Totals
110
192
1
Native
Extreme
0
0
2
Native
Low
40
8
3
Native
Extreme
0
8
4
Native
Extreme
0
30
5
Native
High
10
3
Totals
50
49
1
Sacajawea
High
10
0
2
Sacajawea
High
10
0
3
Sacajawea
Moderate
20
0
4
Sacajawea
Low-no
40
0
5
Sacajawea
Moderate
20
0
Totals
100
0
Table 1: Camas Plot Estimated and Observed Damage. Damage values are estimates of how many of the original 40 bulbs are likely to remain in each plot.
While our findings from this unexpected study of bulbs were unfortunate, they tell an interesting story. An important point to note is that many of the bulbs have divided since they were planted, which is why in a few cases we found more than the original 40 planted bulbs. Regardless, there is a clear preference for the native C. leichtlinii and native cultivar ‘Sacajawea’ bulbs over the ‘Blue Heaven’ cultivar. We also noticed that any bulbs that were planted more shallow than the recommended 2-3x the height of the bulb were missed by the gophers.
Finding the Gopher Stash
After the exploratory bulb digging, we excavated each of our camas plots to around 1 foot in depth to install fences to keep the gophers from returning to our plots. While digging out the excess soil, we would often find a bulb or two that weren’t located during the initial bulb removal (these numbers are not included in Table 1, as we did not record them). In one section where the three camas types were planted in a row, we excavated a huge section of the garden, and made an amazing discovery (extra Kudos to Tyler who did the bulk of the work on this section).
On one of the walls of the hole, we found a gopher food chamber with thick white roots sticking out of the bottom of it. We removed some soil from the entrance, and discovered a chamber filled with camas bulbs. We carefully removed them and found over 60 bulbs that had been stolen from our plots.
The 3excavated plots, the food chamber, and the pile of 66 bulbs removed from the burrow.
Some of the bulbs were clearly the wild type great camas, identified by their characteristic long neck. The others we suspect to be ‘Sacajawea’ bulbs, as the burrow was found in what used to be a ‘Sacajawea’ plot. Any unknown bulbs were brought to my home and planted in a planter box to be identified in the next couple of months. The ‘Sacajawea’ bulbs have variegated foliage, making them easy to pick out once their shoots appear above the soil. We won’t know if the remaining mystery bulbs are ‘Blue Heaven’ or large wild type bulbs until they bloom in the spring.
Moving Forward
On the left: Jen (me) building a gopher exclosure. On the right: Tyler finishing installing a gopher exclosure.
In November of 2020 we installed our fences, refilled the gaping holes with soil, and replanted all of the camas bulbs, including some supplemental purchased bulbs of each of the three varieties. The native Camas and ‘Blue Heaven’ were successfully replanted with 40 bulbs. We were only able to order enough ‘Sacajawea’ bulbs to achieve a density of 30 bulbs per plot, though they will receive additional geophytes if any of the mystery bulbs turn out to be variegated. The mystery bulbs have yet to push their shoots through the soil, but I will include an update on their identities when I have them.
Thank you to Tyler, Izzy, Max, and my fiancé Elliot for helping out in this laborious process. I absolutely would not have been able to safeguard the new camas plantings without your efforts and support in this process.
This past year presented challenge and change to the Garden Ecology Lab. COVID locked us out of the lab and out of the field for a period of time. We said goodbye to two lab members (Angelee graduated! Cliff decided to move on from graduate school), and said hello to new lab mates (Cara took over Cliff’s project; Gwynne started her post-doc; Tyler, Jay, and Max all joined the lab as undergraduate researchers and research assistants). In addition to COVID and personnel changes, I had orthopedic surgery that took me away from work for a little under a month.
But somehow, despite the challenges and changes, we managed to make progress on several research projects. Below, I present a partial reporting of the Garden Ecology Lab year in review for 2020. Besides each project heading is the name of the project lead(s).
1) Garden Bees of Portland (Gail & Isabella):Jason Gibbs’ group from the University of Manitoba provided final determinations for a particularly difficult group of bees to identify: the Lasioglossum sweat bees. In addition, Lincoln (Linc) Best provided determinations for garden bees collected in 2019. Isabella is entering in some of our last remaining specimens, and I am working through the database of over 2,700 collected specimens to ‘clean’ the data and double check data entry against specimens in hand. There are a few specimens that need to be re-examined by Linc, now that we have determinations from the University of Manitoba, the American Museum of Natural History (Sarah Kornbluth), and a graduate of Jim River’s lab (Gabe Foote).
Altogether, we collected between 76 and 84 species of bee across a combined acreage of 13.2 acres (sum total acreage of 25 gardens). The low end estimate conservatively assumes that each unique morphospecies (i.e. Sphecodes sp. 1 and Sphecodes sp. 2) are a single species, whereas the high end estimate assumes that each is a unique species. A few noteworthy specimens:
We collected one specimen of Pseudoanthidum nanum, which is a non-native species to our area, which seems to be establishing and spreading in Portland. Stefanie Steele from Portland State University is writing a note on this apparent introduction, and is using data associated with our single specimen in her paper.
We collected one specimen of Lasioglossum nr. cordleyi which might or might not be a new species. The notation nr. cordleyi means that this specimen looks similar to L. cordleyi, but that the morphology of this specimen is different enough than the normal ‘type’ for this species, that it catches your attention. Jason Gibbs’ group is retaining that specimen. Further study will be needed to determine if it is indeed a new species, or not.
Some of the species we collected (as well as their ecological characteristics) suggest that gardens might be healthy habitat for bees. For example, we collected 72 specimens of Panurginus atriceps, which is a ground-nesting, spring-flying bee. Previous studies of garden bee fauna found ground-nesting and spring-flying bees to be relatively rare. We found them to be surprisingly (but relatively) common in our collections. We also collected seven putative species and 23 specimens of Sphecodes bees. This type of bee is a social parasite that does not collect nectar or pollen or construct a nest for their brood. Instead, they take advantage of the hard work of other bee species, by laying their eggs in the nest of another female. Parasitic bees are often used as bioindicators of habitat health. They would not be present on a site, unless the site also supported their obligate hosts.
We collected two species of bee that are listed on the IUCN red list for threatened and endangered species: Bombus fervidus (18 specimens) and Bombus caliginosus (10 specimens). I am not yet sure if their presence in urban gardens suggests that these species are recovering, that these species might be urban-associates that would be expected to thrive in urban gardens, and/or if gardens might represent particularly good habitat for these species.
In 2021, I *hope* that I can complete gathering data for this study, so that I can begin to analyze data and write. I hope to make it out to every garden, one last time, to finalize garden maps that will be used to calculate the area allotted to ornamental plants, edible plants, hardscape, and unmanaged areas. Aaron has already mapped out the landscape surrounding each garden at radii of 500 and 1000 meters. Together, these data will be used to understand whether/how garden composition and the surrounding landscape interact to influence bee species richness.
OLYMPUS DIGITAL CAMERA
Gail collecting bees off of flowers, using an insect aspirator (left). A wildflower meadow in one of our largest gardens (center). Some of the over 2,000 specimens we collected, and Isabella pinned and labelled.
2) Native Plants and Pollinators (Aaron Anderson): In February, Aaron successfully defended his dissertation proposal and passed his oral examination, and thus advanced to Ph.D. candidacy!! Since that time, he has been busy sorting, identifying, and counting three years’ of insect samples from his 140 study plots, representing five replicates plots of 23 native plants, four ornamental plants, and a control ~ a task that he finished two weeks ago! His bees have been identified to species by Linc. Aaron has identified the thousands of other insects in his samples to the taxonomic level of family. He is working through analysis of his massive data set, and is simultaneously working on two manuscripts: one focused on just the bees and the other covering all other insects. We plan to turn the key points of these two chapters into an infographic that can be used by gardeners and green industry professionals, to select native plants that support an abundant and diverse assemblage of beneficial insects.
Aaron recently submitted the first paper from his dissertation for publication consideration, to the journal HortTechnology ~ and it was accepted, pending revisions! This paper reports on his survey of gardeners’ impressions of the aesthetic value of his study plants, and includes five specific recommendations for native wildflowers that Pacific Northwest nurseries might consider growing and marketing as pollinator plants (e.g. Gilia capitata, Clarkia amoena, Eschscholzia californica, Madia elegans, and Sidalcea asprella virgata). These plants all fell within the ‘sweet spot’ of being attractive to both pollinators and to gardeners.
Aaron’s plots at the NWREC station remain in place. Although we are through collecting data for Aaron’s study, I am applying for grant funding to study how plant traits ~ both the reward that plants offer pollinators and the displays that they use to attract pollinators ~ change with plant breeding for specific aesthetic traits, and whether/how these changes affect pollinator visitation. We also hope to study how highly attractive pollinator plants function in mixed plantings and in garden settings.
Aaron and Lucas measuring plots in year one (left); Douglas Aster, one of Aaron’s most attractive plants for bees (center); Oregon sunshine was less attractive to bees than we had expected.
3) Bees on Native Plants and Native Cultivars (Jen Hayes):
Jen successfully completed her first field season of research, which is a monumental accomplishment during this time of COVID restrictions on our work. In early 2020, Jen finalized her list of study plants, which included one native species and 1-2 hybrids or native cultivars. This, in and of itself, was a huge accomplishment. Although we started with a much broader list of potential study plants, so many native plants did not have native cultivars or appropriate hybrids available for sale.
Jen’s study plants, which include one native (top photo in each group) and 1-2 native cultivars or native hybrids.
Once Jen and her crew put the plants in the ground, a new set of challenges emerged. For example the native yarrow emerged with pink flowers, which was a clear signal that these plants were not true natives. In addition, the Sidalcea cultivars that Jen and her crew planted came up looking different than the Sidalcea native. This sent Jen on a journey to the OSU Herbarium, where she learned that the Willamette Valley’s native Sidalcea malviflora has been reclassified as Sidalcea asprella, and that the cultivars we purchased were hybrids of Sidalcea malviflora (native to SW Oregon and California). This all suggests a need to work with local nurseries and/or growers of native plants, to see whether or not there needs to be or can be standards for sale of native plants. Should native species and native cultivars be verified or share provenance? Should gardeners be asking for this information? I don’t know, but I think that they’re important questions to consider.
With one field season’s worth of data in hand, the native cultivars were more attractive to all bees (with overall patterns being driven by the abundance of the European honey bee) for all floral sets, except California poppy. When we excluded honey bees from the analysis, to look at (mostly) native bees, no clear pattern of visitation on native plants versus native cultivars emerged. Native California poppy was most attractive to native bees. But, native cultivars of Sidalcea were more attractive to native bees (keeping in mind that in 2020, our native cultivars were not cultivars of our regionally appropriate native plant). For all other plants, there was no difference. We look forward to collecting additional data in 2021 and 2022, to see if the lack of difference in bee visits to native plants versus native cultivars holds up. Particularly for the perennials, we are finding that bee visits change so much from year to year, as the plant becomes established.
Jen, pre-COVID times, discussing her research at the Urban Ecology Research Consortium conference (left). On every study plant, Jen takes data on corolla width and depth, as well as nectar tube length (center). Jen’s study site, with California poppy in the foreground, and a cultivar of California poppy towards the back (right).
4) Garden Microbes in Soil and on Skin (Dr. Gwynne Mhuireach): Dr. Mhuireach successfully recruited 40 gardeners to participate in this study: 20 from western Oregon and 20 from the high desert. She has received and processed all soil samples and all skin swab samples for PCR (genotyping), which will be used to infer the diversity and identity of the soil microbial community in garden soils and on gardeners’ skin. She has also received survey responses from all study participants, so that she can characterize gardeners’ crop types, time in the garden, and gardening practices (e.g. organic, conventional, or mixed).
Dr. Mhuireach then sent me the soil samples, so that I could process them for submission to OSU’s Soil Health Lab. The Soil Health Lab is currently performing the chemical and physical analyses on each soil sample, so that we can determine if there is any relationship between soil characteristics, gardening region (e.g. western Oregon or high desert), crop choices, management practices, and the microbes that can be found in garden soils and/or on gardeners’ skin. Gwynne just received the first data back from the PCR analyses ~ and we can’t wait to share some of the intriguing findings with you, after we’ve had some time to process and digest the data!
Because of COVID-19 lab closures, we are a bit behind where we had hoped to be at this point. We anticipate receiving all data from each service lab by the end of January or in early February. You can read more about Gwynne’s project, here.
**********
Beyond these four studies, Tyler started his BioResource Research project (costs and yield of container grown and intercropped tomotoes), and Isabella worked on her thesis (parasitoids in Portland area gardens). We also collaborated with OSU Computer Science students to turn a database of first frost / last freeze dates that Angelee compiled, into a web-based app (the app is still in beta-testing, but we hope to release it, soon!). I will detail those studies, in another post. But for now, I’m getting excited for the smell of carnitas that is filling the house, and that will go on top of the New Years’ nachos that will help us ring in 2021! I hope that you all have a very Happy New Year, and that 2021 brings health, and happiness, and joy to all.
I distinctly remember the day that I decided I wanted to study wild bees. I was sitting in a darkened auditorium at the American Museum of Natural History in New York City, listening to Claire Kremen deliver the plenary address in a symposium focused on invertebrate conservation.
In her address, Dr. Kremen shared the results of her research on watermelon farms in California’s Central Valley. Like all cucurbits, watermelon requires insect pollination to set fruit. Watermelon, in particular, has a high pollination requirement: it takes at least eight bee visits to deposit the 500-1,000 viable pollen grains needed to set harvestable fruit in seeded watermelons. Seedless watermelons require between 16 and 24 visits by a bee in order to set fruit. Most growers meet the high pollination requirement of melons by renting and placing honey bee hives in fields. Dr. Kremen’s research suggested that a different approach might be possible.
Honey bees are important agricultural pollinators, in large part because they can be moved en mass and placed into farm fields at the exact moment that pollination services are needed.Photo by Isabella Messer.
Dr. Kremen and her colleagues studied three types of watermelon fields that varied in their pest management practices (organic or conventional) and their proximity (near or far) to native habitat in the foothills of the California coast range. These fields were: organic near, organic far, and conventional far. Watermelon fields that used conventional pest management practices and that were located near the foothills were not included in this study because this type of farm did not exist in the region. The team determined the number of pollen grains that different bee species deposited on watermelon, by presenting different bees with a single watermelon blossom that had yet to receive any insect visits. After a blossom was visited by a single bee, the flower was bagged and tagged accordingly, and the number of pollen grains deposited on the stigma by that single bee was counted in the lab. They repeated this process for 13 different bee species.
Next, the team sat in watermelon fields and observed what types of bees visited watermelon blossoms, in different types of farm fields. Watermelon flowers are only receptive to pollination visits for a single day. They recorded the sex and species of each bee visitor to the blossoms. Based upon these species specific counts, combined with the pollen deposition data (above), they were able to assess how much each particular bee species contributed to the production of harvest-ready watermelon.
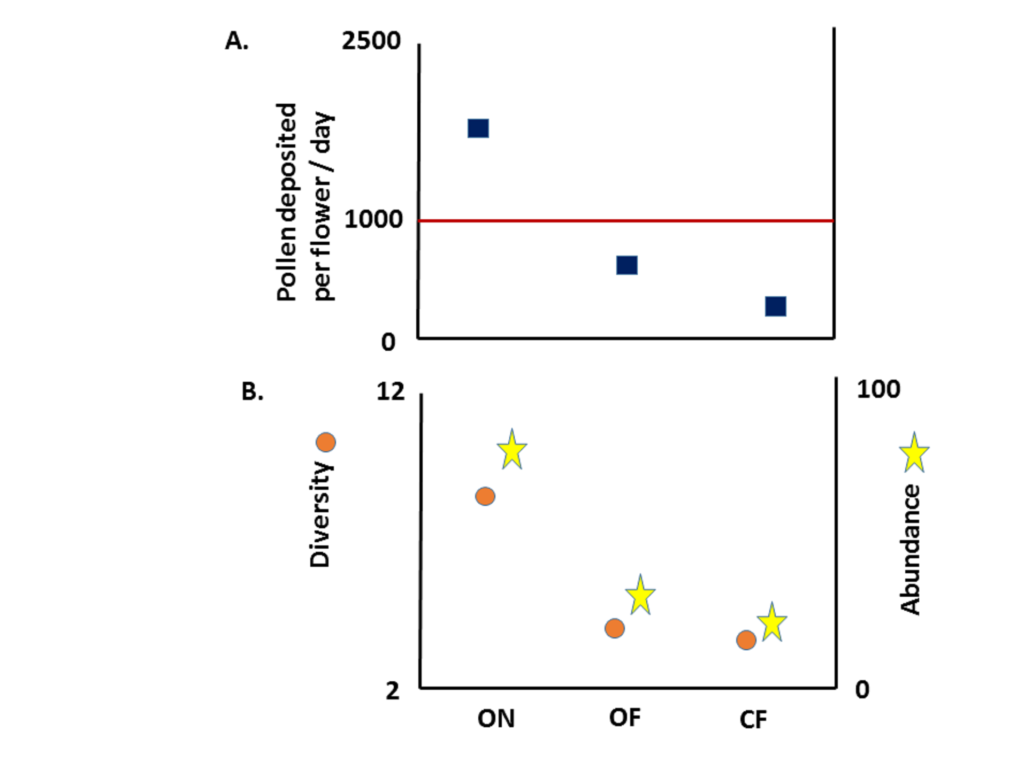
Dr. Kremen found that pollination surpassed 1,000 pollen grains needed to set harvestable fruit per flower in the organic near fields but not in the organic far or the conventional far field (Fig 1, part A). Furthermore, she found that this outcome could be tied to the greater diversity and abundance of bees in organic near fields, compared to the other two types of fields (Fig 1, part B).
Figure 1. The number of pollen grains deposited on watermelon stigmas (A), and the diversity (orange circles) and abundance (yellow stars) of bees (B) on farms that were classified as organic near (ON), organic far (OF), and conventional far (CF). The number of pollen grains required to set marketable fruit (1000) is noted via a red threshold line in A. (modified from Kremen et al. 2002).
Dr. Kremen also found that, even though no single species of wild bee was as effective as managed honey bees, the collective group of wild bees surpassed the effectiveness of honey bees in organic near fields (Figure 2). Interestingly, honey bees were most effective as crop pollinators in the conventional far fields and least effective as crop pollinators in the organic near fields. This may be because few other flowers were in bloom in the conventional far fields, so that honey bees concentrated their attention on the crop at hand. In the organic near fields, a greater diversity of flowering plants likely competed for the pollination services of honey bees.
Figure 2. The cumulative contribution of native bees, compared to the contribution of honey bees to the pollination requirement of watermelons on organic near (orange circles), organic far (navy squares), and conventional far (yellow stars) farms (modified from Kremen et al. 2002).
Wild bees were able to fully satisfy the pollination requirements of a crop with an extremely high pollination requirement because broad spectrum insecticides were not used, and the foothills provided year-round and protected habitat for the bees. This story blew my mind!
Prior to that conference, I had never given wild bees much thought. They’re mostly solitary nesters, with small bodies, that only forage for a few days to a few weeks. They tend to be inefficient foragers, particularly when compared to the juggernaut of a honey bee hive. Whereas wild bees are akin to a single vendor on Etsy, honey bees seemed the unbeatable Amazon!
Dr. Kremen’s work showed the potential value that wild bees have to agriculture. And her work was published just prior to the global onset of colony collapse disorder in honey bees in 2006. It set off a worldwide discussion about what to do about honey bee losses. Should scientists put time and effort into saving a single, non-native species (the honey bee), or should we work to conserve or build habitat around farm fields while also reducing insecticide use?
I was incredibly hopeful that the simultaneous threat to honey bees and promise of wild bees might promote heavier investments in agroecology, including the conservation of bee-friendly habitat around farms. During this time period, I was also in the early stages of documenting wild bee biodiversity in community and residential gardens, and I was surprised that abundance and diversity of garden bees was much higher than I had anticipated.
Back in 2004, I started to see gardens, and the abundance and diversity of wild bees that they host, as a potential solution to the problem of colony collapse disorder. Although I continue to be fascinated by the potential role of home and community gardens as a safe haven for bees from agricultural stresses, the urgency of this question has faded. Colony collapse disorder does not currently plague honey bees, due in large part to federal investments in studying, understanding, and mediating the factors that contribute to failing hives. With honey bees doing much better, attention has somewhat faded on the potential role of wild bees as crop pollinators. Still, work in this area continues and may rise to renewed importance, should colony collapse disorder again present a major challenge to United States agriculture.
Wild bees, including this leaf-cutter bee in the genus Stelis are also potentially important crop pollinators. However, many farm practices, such as regular insecticide sprays and mono-cultural cropping systems, make farms inhospitable to wild bees. Photo by Isabella Messer.
Kremen, Williams, Thorp. 2002. Crop pollination from native bees at risk from agricultural intensification. Proc. Natl. Acad. Sci. 99: 16812-26816. DOI: 10.1073/pnas.262413599.