There are moments in our individual lifetimes that we can define as noteworthy and right now, as I prepare to start my graduate career within the Marine Mammal Institute (MMI) at OSU, I would say this is it for me. As I sit down to write this blog and document how surreal my future adventure is, I simultaneously feel this path is felicitous. After a year of being cooped up due to COVID, time presently seems to be going by at rocket speed. I am moving constantly in through my day to continue running my current life, while simultaneously arranging all that will encompass my new life. And while I answer questions to my 10-year-old daughter who is doing geometry homework in the living room, while hollering “That is not yours!” to the kitchen where the recently adopted feral dog is sticking his entire head under the trash can lid, while arranging our books in a cardboard box at the packing station I set up on the dining room table, I cannot deny a sense of serenity. This moment in my life, becoming a part of the GEMM Lab and MMI, and relocating to Corvallis is great.
This moment’s noteworthiness is emphasized by embarking on probably the most variable-heavy road trip I have planned to date. Since the age of 19, when I left my small mountain town on the Appalachian trail in Pennsylvania, I have transferred locations ~20 times. Due to extensive travel while serving in the Army (various Army trainings and overseas mission deployments), I have bounced around the US and to other countries often. Over time, one becomes acclimated to the hectic nature of this sort of lifestyle, and yet this new adventure holds significance.
So here are the details of the adventure trip that lies ahead: I will drive my 2002 Jeep Grand Cherokee across the country; from Charlottesville, Virginia to Corvallis, Oregon. My projected route will extend 2,822 miles and take ~43 driving hours total. The route will fall within the boundaries of 11 states (see Figure 1.)
Figure 1. Blue Line indicates route from Charlottesville to Corvallis (Google Maps)
Attached to the hitch of the Jeep will be a 6×12 rented cargo trailer containing our treasured books, furniture and things. Inside the Jeep will be three living variables: Mia (the 10-year-old), Angus (hyperactive border collie/ pit bull mix) and Mr. Gibbs (feral pirate dog); all three will need to be closely monitored for potential hiccups in the plan.
If we are going to make it to our destination hotel/Airbnb each night of the trip, I must be organized and calculate road time each day while factoring in breaks to the loo and fueling up. These calculations need to be precise, with little margin for error. I cannot play it too safely either, or it will take us too long to get across the country (I must start my graduate work after all). On the other hand, I cannot realistically expect too many road hours in a day. I think at this point I have got it worked out (Table 1.)
Table 1. Driving Hours and Miles Per Day
When I look back on my career, I had no idea that my not-so-smooth road would lead me to my dream goal of studying marine mammals. I took the Army placement tests at the age of 19, which led me to the field of “information operations” where I earned a great knowledge base in data analysis and encountered fantastic leaders whom I might not have known otherwise. I learned immensely on this path and it set me up very well for moving forward into research and collaboration in the sciences. I am so grateful that my life took this journey because working in the military provided me with the utmost respect for my opportunities and greater empathy for others. This route had many extreme obstacles and was intensely intimidating at times, but I am all the better for it. And I was never able to shake the dream of where I wanted to be (see Figures 2 & 3.) Timing is everything.
Figure 2 & 3. Two of the images of the Pacific coast I have hung up in my house. Keeping my eye on the prize, so to speak.
It will feel great to cross over the Oregon state line. I cannot wait to meet GEMM Lab in-person and all the other wonderful researchers and staff at MMI and Hatfield Marine Science Center. I am eager to step onto the RV Pacific Storm and begin my thesis research on the magnificent cetaceans off the Oregon coast, and hopefully do some good in the end. As I evaluate the logistics of my trip from Charlottesville to Corvallis, I feel relieved rather than overwhelmed. We could attribute this relief to my not-so-smooth road to get to where I am. Looking ahead, of course, I see a road that will require focus, attention, passion, care, and lots of fuel. Even if this road is not completely smooth, I will have my hands on 10 and 2, and feel so grateful and ready to be on it.
¹PhD Student, OSU Department of Fisheries, Wildlife, and Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
²Postdoctoral Scholar, OSU Department of Fisheries, Wildlife, and Conservation Sciences, Cooperative Institute for Marine Resources Studies, Hatfield Marine Science Center
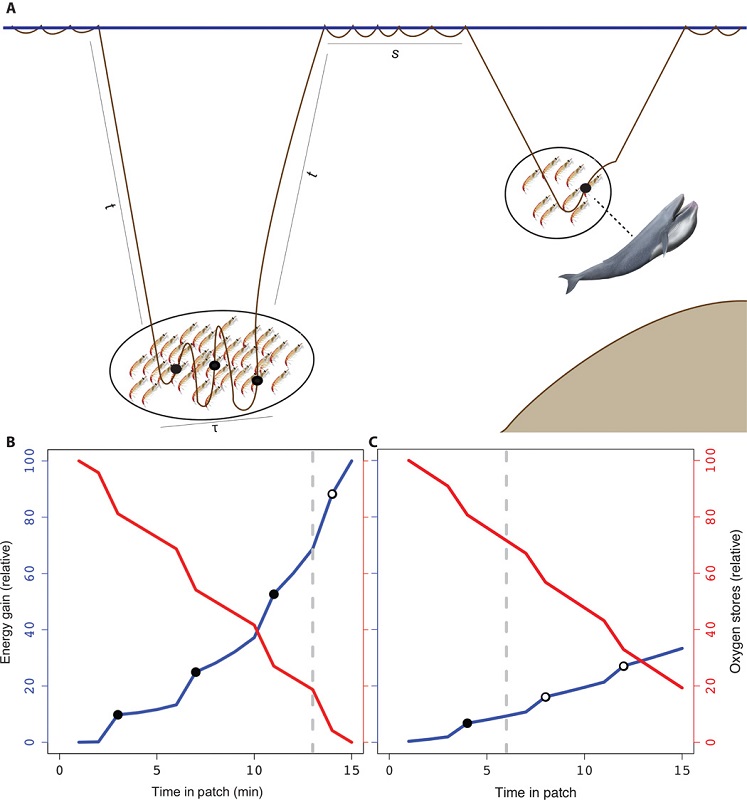
Many aspects of studying cetacean ecology, behavior, population dynamics, health, and sociality depend on being able to see and/or sample cetaceans when they come to the surface. While this research is not necessarily easy given that cetaceans spend the majority of their time underwater out of human sight, it is definitely feasible, as evidenced by decades of cetacean research. However, in order for researchers to observe cetaceans at the surface they need to get out to sea, and this boat-based effort can realistically only be done in good ocean conditions. Any sea-going individual likely uses the Beaufort sea state (BSS) scale as a measure of ocean conditions. For a full breakdown and excellent explanation of what the BSS is, check out our beloved Alexa’s blog; but for the purposes of this blog all you really need to know is that the smaller the BSS (which starts at 0), the calmer the ocean, and the higher the BSS, the rougher & stormier the ocean. There are two main reasons for conducting cetacean research in low BSS: 1) above a certain threshold (usually BSS 4) it becomes difficult to reliably spot and recognize cetaceans at the surface, thus compromising good data collection, and/or 2) to ensure safety and comfort of the research team.
So, when the BSS gets too high, us humans usually do not go out to sea to study cetaceans, which means that the cetaceans, for the most part, go unobserved. So, many questions arise about what cetaceans are doing during these rough ocean conditions. What does an increased BSS mean for them? Are they unfazed by big waves and strong winds, or are they affected by the weather and take longer dives or seek out fairer seas? A conversation among friends sparked our curiosity of what cetaceans do in stormy conditions and inspired us to collaborate on this blog. Here, we report on what is and is not known about cetaceans in storms, and discuss some ideas about how best to quantify the effects of rough sea conditions on cetaceans.
Slide the arrows to compare sea conditions (BSS 1 [left] vs BSS 6 [right]) experienced by Alexa, the GEMM Lab marine mammal observer on the May 2019 Northern California Current cruise onboard NOAA ship Bell M. Shimada. Source: A. Kownacki/GEMM Lab.
A literature search of cetaceans during storms did not generate many results, which was not surprising to us given the above reasons about researchers not being able to survey in rough sea conditions. However, we did find a couple of interesting studies about cetacean behavior and distribution after storms.
Changes in foraging behavior
Autumnal storms in Maryland, USA resulted in less frequent and shorter encounters of bottlenose dolphins in the US Mid-Atlantic Bight. However, dolphins spent a significantly higher percentage of their encounters feeding after storms than they did before or during them (Fandel et al. 2020). Similarly, bottlenose dolphins in Mississippi Sound displayed an approximately 15% increase in foraging activity for up to 2 years following Hurricane Katrina (Smith et al. 2013). These changes in foraging behavior are attributed to shifts in distributions and behavior of dolphin prey species as a result of altered environmental conditions (primarily sea surface temperature and salinity) following the hurricanes.
Out-of-habitat events and strandings
An out-of-habitat event occurs when an animal is displaced out of its typical habitat. Seven of these events were reported following Hurricane Rita, which hit the southwest Louisiana coast in 2005, with bottlenose dolphins found in flooded roadside ditches, canals, shallow flooded fields, and a natural creek area (Rosel & Watts 2008). These locations ranged from 2.5 to 11 km inland from the coast of the Gulf of Mexico, where these dolphins were displaced from. It is believed that the animals were carried inland on the storm surge that accompanied Hurricane Rita and were left stranded in areas that held water the longest once it started receding (Rosel & Watts 2008).
One of the roadside ditches where a bottlenose dolphin was trapped in Louisiana following Hurricane Rita. Taken from Rosel & Watts (2008).
There have been two mass strandings of pygmy killer whales that are believed to have been a result of hurricanes. In 1995, five pygmy killer whales stranded (three of which died, while two were successfully refloated) in the British Virgin Islands a day after Hurricane Marilyn (Mignucci-Giannoni et al. 1999). In 2006, six pygmy killer whales (five of which died) stranded in New Caledonia during and after Hurricane Jim (Clua et al. 2014). Both studies hypothesize that increased energetic costs, as a result of attempting to evade the hurricanes, coupled with animals becoming disoriented and ending up in shallow waters, is what caused them to strand.
While these studies reveal post-storm effects on cetaceans, we still do not know exactly how these individuals behaved during the storms. Did they attempt longer dives to stay away from the rough conditions at the surface, thus becoming disoriented? Or were they behaving normally (i.e. foraging, travelling) and were simply “pushed” into waters that they did not intend to go into? Given that very stormy sea conditions do not allow for visual, boat-based surveys, we need to employ different technologies to study cetacean behavior and distribution during storms.
Passive acoustic monitoring (PAM) is a great tool that can monitor ocean environments for us when the seas are too stormy. Using fixed or mobile platforms, underwater PAM listening devices (hydrophone and data storage) can record sounds in the ocean for us to listen and analyze from shore. With PAM we are able to track the vocalizations of marine mammals as well as other sounds in the environment, such as waves crashing and rain. Anecdotally, we have spent many days at sea in conditions that were too rough for visual observations, but we could safely use our PAM tools to detect cetaceans. So, just because the seas may be too rough to see cetaceans, this fact does not mean that we cannot observe them – we just need to listen instead of look.
There are many tools that can be used to record underwater sounds, including passive acoustic monitoring (PAM; shown in orange), real-time acoustic data collection (green), and active acoustics (blue.) Source: NOAA Fisheries.
A number of studies have investigated whether whales change their vocalization behavior differently in response to changing ambient sound conditions (for example: Dunlop et al. 2010; Fournet et al. 2018). While research on ocean sound levels is often focused on the impact of human-generated or anthropogenic noise, there are also natural, abiotic sound sources (e.g. wind, rain, ice) that can elevate ambient sound levels. One potential animal response to elevated ambient sound levels is to vocalize at a higher intensity, called the Lombard (or cocktail party) effect. This phenomenon is common for us humans – have you ever been at a party and at some point you realize that you are shouting to someone in order to be heard above the noise of the room? That’s the Lombard effect! Humpback whales in Glacier Bay National Park, Alaska, exhibited the Lombard effect in response to both natural and man-made sounds, but the probability of calling was lower when vessels were present compared to times with only natural sounds (Fournet et al. 2018). It is also possible that whales may vocalize at different frequencies, times, or for shorter durations when the ocean becomes louder, which we can easily track with PAM. Unfortunately, PAM is limited to what we are able to hear, so if we do not hear whales we cannot determine if this result is because their vocalizations are masked by higher intensity sounds, if they stopped vocalizing, or if they left the listening area.
Animal-borne tags are another kind of autonomous observation tool that could help us understand cetacean behavior and distribution in storms. Admittedly, the logistics of applying tags before an imminent storm are probably complex. However, the development of medium-duration archival tags may provide a good trade-off between deploying tags long enough before a storm begins, thus providing safe working conditions for the research team, while minimizing potential physical impacts to the animals (Szesciorka et al. 2016). There are currently no published tag studies that document cetacean behavior during storms, but a study of a gray-headed albatross, fitted with a satellite transmitter, that successfully foraged during an Antarctic storm (Catry et al. 2004) shows the promise of using animal-borne tags to answer these questions.
As with many questions about animal behavior, our best option is to combine all of our research tools to piece together evidence about what might be going on in the deep, dark, stormy ocean. Simultaneously collecting acoustic and movement & behavior data through PAM and animal-borne tags, respectively, could allow us to determine how cetaceans behave during storms. While we are probably not poised to tackle these questions right now, perhaps another curious graduate student can take it on for their own PhD research…
References
Catry, P., Phillips, R.A., and J.P. Croxall. Sustained fast travel by a gray-headed albatross (Thalassarchie chrysostoma) riding an Antarctic storm. The Auk 121(4):1208-1213.
Clua, E.E., Manire, C.A., and C. Garrigue. 2014. Biological data of pygmy killer whales (Feresa attenuata) from a mass stranding in New Caledonia (South Pacific) associated with Hurricane Jim in 2006. Aquatic Mammals 40(2):162-172.
Dunlop, R.A., Cato, D.H., and M.J. Noad. 2010. Your attention please: increasing ambient noise levels elicits a change in communication behaviour in humpback whales (Megaptera novaeangliae). Proceedings of the Royal Society B 277(1693):doi.org/10.1098/rspb.2009.2319.
Fandel, A.D., Garrod, A., Hoover, A.L., Wingfield, J.E., Lyubchich, V., Secor, D.H., Hodge, K.B., Rice, A.N., and H. Bailey. 2020. Effects of intense storm events on dolphin occurrence and foraging behavior. Scientific Reports 10:19247.
Fournet, M.E.H., Matthews, L.P., Gabriele, C.M., Haver, S., Mellinger, D.K., and H. Klinck. 2018. Humpback whales Megaptera novaeangliae alter calling behavior in response to natural sounds and vessel noise. Marine Ecology Progress Series 607:251-268.
Mignucci-Giannoni, A.A., Toyos-González, G. M., Pérez-Padilla, J., Rodríguez-López, M. A., and J. Overing. 1999. Mass stranding of pygmy killer whales (Feresa attenuata) in the British Virgin Islands. Journal of the Marine Biological Association of the United Kingdom 80:759-760.
Rosel, P.E., and H. Watts. 2008. Hurricane impacts on bottlenose dolphins in the northern Gulf of Mexico. Gulf of Mexico Science 25:7.
Smith, C.E., Hurley, B.J., Toms, C.N., Mackey, A.D., Solangi, M., and S.A. Kuczaj II. 2013. Hurricane impacts on the foraging patterns of bottlenose dolphins Tursiops truncatus in Mississippi Sound. Marine Ecology Progress Series 487:231-244.
Szesciorka, A.R., Calambokidis, J., and J.T. Harvey. 2016. Testing tag attachments to increase the attachment duration of archival tags on baleen whales. Animal Biotelemetry 4:18.
By Alejandro Fernandez Ajo, PhD student in the Department of Biology, Northern Arizona University, visiting scientist in the GEMM Lab working on the gray whale physiology and ecology project
When a cetacean (whales and dolphins) is ashore or trapped in nearshore waters and cannot return to the open waters, it is considered stranded. Frequently, the stranded animal is in distress, dying, or dead. Although rare, the stranded cetacean can be a healthy animal trapped due to changes in tide or disorientation. Every year many cetacean strandings are reported from along the coasts around the world, and likely many more stranding events go unnoticed when they occur in remote areas. In all cases, the question is: why do cetaceans beach?
Southern right whales stranded at the coast of Peninsula Valdés, Patagonia-Argentina. Photo: Matias DiMartino / Southern Right Whale Health Monitoring Program.
There may be different causes for whales and dolphins to strand on beaches, either dead or alive. Understanding and investigating the causes of cetaceans strandings is critical because they can be indicators of ocean health, can help identify anthropogenic sources of disturbance, and can give insights into larger environmental issues that may also have implications for human health (NOAA). In this context, when scientists are analyzing a stranding event, they consider both possibilities that the event was natural or human-caused and classify strandings according to specific characteristics to study the causes of these events.
Types of cetacean strandings:
Live or Dead Stranding:
A stranding can involve live animals or dead animals if the death occurs in the sea and the body is thrown ashore by wind or currents. In live strandings, when they occur near urbanized areas, usually significant efforts are made to rescue and return the animals to the water; with small odontocetes, sometimes there is success, and animals can be rescued. However, when large whales are beached alive, their own weight out of the water can compress their organs and can cause irreversible internal damage. Although not externally visible, such damage can sometimes cause the death of the animal even after returning to the sea.
According to the number of individuals:
Single strandings occur when only a single specimen is affected at the time. The cetaceans that most frequently strand individually are the baleen (or mysticete) whales, such as right and humpback whales, due to their often solitary habits.
Mass strandings comprise two or more specimens, and in some cases, it can involve tens or even a few hundred animals. The mass strandings are more frequently observed for the odontocetes, such as pilot whales, false killer whales, and sperm whales with more complex social structures and gregarious habits.
Left: Single southern right whale calf stranded at the coast of Peninsula Valdés, Patagonia-Argentina. Ph.: Mariano Sironi / ICB. Right: Mass stranding of common dolphins in Patagonia-Argentina. Photo: www.elpais.com
Unusual Mortality Events
The Marine Mammal Protection Act defines an unusual mortality event (UME) as a stranding event that is unexpected, involves a significant die-off of any marine mammal population, and demands immediate response. Seven criteria make a mortality event “unusual.” Source: https://www.fisheries.noaa.gov.
A marked increase in the magnitude or a marked change in morbidity, mortality, or strandings when compared with prior records.
A temporal change in morbidity, mortality, or strandings is occurring.
A spatial change in morbidity, mortality, or strandings is occurring.
The species, age, or sex composition of the affected animals is different than that of animals that are normally affected.
Affected animals exhibit similar or unusual pathologic findings, behavior patterns, clinical signs, or general physical condition (e.g., blubber thickness).
Potentially significant morbidity, mortality, or stranding is observed in species, stocks, or populations that are particularly vulnerable (e.g., listed as depleted, threatened, or endangered, or declining). For example, stranding of three or four right whales may be cause for great concern, whereas stranding of a similar number of fin whales may not.
Morbidity is observed concurrent with or as part of an unexplained continual decline of a marine mammal population, stock, or species.
The purpose of the classification of a mortality event as a UME is to activate an emergency response that aims to minimize deaths, determine the event cause, or causes, determine the effect of the event on the population, and identify the role of environmental parameters in the event. Such classification authorizes a federal investigation that is led by the expertise of the Working Group on Marine Mammal Unusual Mortality Events to investigate the event. This working group is comprised of experts from scientific and academic institutions, conservation organizations, and state and federal agencies, all of whom work closely with stranding networks and have a wide variety of experience in biology, toxicology, pathology, ecology, and epidemiology.
Southern right whale necropsy and external measurements. Source: Southern Right Whale Health Monitoring Program / ICB.
What can be learned from strandings and UMEs?
Examining stranded marine mammals can provide valuable insight into marine mammal health and identify environmental factors leading to strandings. Through forensic examinations, the aim is to identify possible risks to whales’ health and evaluate their susceptibility to diseases, pollutants, and other stressors. This information can contribute to cetacean conservation through informed management strategies. However, the quality of the data derived from a necropsy (the postmortem examination of carcasses) is highly contingent upon how early the stranding event is reported. As soon as the animal is deceased, decomposition starts, hindering the possibilities of detailed investigations of the cause of death.
Therefore, a solid network that can report and respond quickly to a stranding event is fundamental; this includes trained personnel, infrastructure, funding, and expertise to respond in a manner that provides for animal welfare (in the case of live strandings) and obtains data on marine mammal health and causes of death. Moreover, a coordinated international organization that integrates national marine mammal stranding networks has also been identifying as a critical aspect to enable adequate response to such mortality events. In many locations and countries around the world, funding, logistical support, and training remain challenging to stranding response.
In response to these concerns and needs, at the last World Marine Mammal Conference, which took place in Barcelona in December of 2019, The Global Stranding Network was founded to “enhance and strengthen international collaboration to (1) ensure consistent, high-quality response to stranded marine mammals globally, and (2) support conservation efforts for species under threat of extinction.” Monitoring marine mammal health worldwide can guide conservation and help identify priority areas for management (Gulland and Stockin, 2020).
What to do in case of finding a whale or dolphin on the beach?
When strandings occur, it is essential to know how to act. Unfortunately, untrained people, often with good intentions, can worsen the situation of stress and injury to the animal or can put themselves at risk of injury or exposure to pathogens. If you find a cetacean alive or dead on the beach, the most important things to do are:
Record information about the location and the animal´s characteristics (the species, if known; the animal’s approximate size; and status (alive or dead)).
Keep at a safe distance: the animal may appear dead to the naked eye and not be. It is important to remember that cetaceans are wild animals and that in stressful situations such as strandings, they can try to defend themselves.
Do not touch the animal: one of the causes of strandings is diseases; therefore, it is advisable not to contact the individuals to avoid exposure to potential pathogens.
If the animal is alive, keep a distance from the animal, especially from its head and tail. Prevent children or dogs from approaching the animal.
Keep calm and do not make noise that could disturb the stranded animal.
Do not take the animal out of the water if it is on the shore or return it to the sea if it is on the beach: Such movement could cause serious injuries, or even death.
Do not feed the animal or give it water: keep the blowhole clear because it is where they breathe.
Proceedings of the workshop “Harmonizing Global Stranding Response.” (2020) World marine mammal Conference Barcelona, Catalonia, Spain. Editors: Gulland F and Stockin K; Ecs Special Publication Series No. 62.
Mazzariol S., Siebert U., Scheinin A., Deaville R., Brownlow A., Uhart M.., Marcondes M., Hernandez G., Stimmelmayr R., Rowles T., Moore K., Gulland F., Meyer M., Grover D., Lindsay P., Chansue N., Stockin K. (2020). Summary of Unusual Cetaceans Strandings Events worldwide (2018-2020). SC-68B/E/09 Rev1.
In September of 2016, Leigh Torres, associate professor at Oregon State University, and I attended the 6th International Albatross and Petrel Conference. Somehow, amid all of the science that filled the week, Leigh first saw the Global Fishing Watch fishing map. She shouted with joy. She immediately envisioned a study to assess interactions between seabirds and fishing boats, and started considering a spatial overlap analysis between telemetry tracks of albatross with the Global Fishing Watch database. Such a study could help reduce bycatch, or the incidental catch of non-target species, like seabirds, in fisheries. Five years later, we executed that study in partnership with Global Fishing Watch, one of the first to look at fine-scale overlap between fishing vessels and marine life on the high seas (Orben et al. 2021).
Transparent data means opportunity for analysis
Despite knowing that bycatch from fisheries is a real, significant problem for many albatross populations, we have long struggled to know where birds go, where boats fish, and where the two interact in the vast ocean, especially in largely unregulated international waters. Albatross are long-lived seabirds and 15 out of the 22 species are threatened with extinction. Scientists have been tracking albatross for three decades, but assessing individual seabird encounters with vessels has traditionally been limited by a lack of transparency in fishing activity data. Some seabirds are attracted to fishing vessels because of the bait and offal, but we don’t know the whole story of why some birds approach vessels while others don’t.
When we first put our relatively large datasets together – 9,992 days of albatross tracking data from 150 birds and Global Fishing Watch fishing effort data from 2012-2016 – we weren’t sure what we would find. The ocean is a big place, and so finding where one bird and one vessel overlap is kind of like trying to find a needle in a haystack. Would we have enough encounters between birds and boats for an analysis? Would birds encounter fishing vessels as often as we think?
Measuring encounters between albatross and vessels
After overlaying the tracking data with a gridded daily layer of fishing effort, we identified potential encounters between birds and fishing boats. We identified when an albatross could detect a vessel, at a radius or 30 kilometers, and when an albatross had a close encounter with a vessel, within a radius of 3 kilometers (following methods developed in Collet Patrick & Weimerskirch, 2015). Then, we investigated factors that influenced the occurrence and duration of close encounters, considering the bird’s behavior, environmental conditions and habitat, fishing vessel and fisheries characteristics, and temporal variables, such as time of day and month.
Species variation of encounters
We conducted our analysis for three species of albatross that forage in the north Pacific ocean, Laysan albatross, black-footed albatross, and short-tailed albatross.
Adult black-footed albatrosses approached vessels for a close encounter 61.9 percent of the time they detected a fishing vessel.
Adult Laysan albatross had close encounters with a fishing boat 35.7 percent of the time they detected a vessel.
Juvenile short-tailed albatross had a lower frequency of close encounters (28.6 percent),
Understanding close encounters and their duration
Due to a low sample size of encounters, we were unable to investigate the reason for close encounters or their duration for black-footed albatrosses. More tracking data is critical to understand factors influencing the impact of vessels on this vulnerable species.
Laysan albatross were more likely to approach fishing vessels when fishing effort was high, but fishing boat density was low. Laysan albatross also had close encounters with vessels more frequently while they were foraging. Due to sample size, we could not further investigate the reason for the duration of encounters for this species.
Short-tailed albatrosses were also more likely to approach fishing vessels when they were searching for prey, fishing effort was high, and fishing boat density was low. They were more likely to have close encounters with vessels during the day and in habitats with water depths from 75-1500 meters.
Vessel attendance by short-tailed albatrosses was longer when sea surface temperatures were warmer and less productive, and during periods with lower wind speeds.
A useful approach
The information available to fisheries managers in order to reduce bycatch is most often limited to data collected from the perspective of the fishing vessels. Our analysis provides an alternative view – an albatross’ view of when and where boats are encountered in the seascape. While our analysis didn’t specifically look at bycatch, our estimates of proximity between birds and boats can be considered a proxy for increased bycatch risk.
For the endangered short-tailed albatross, bycatch events are few, but they come with high consequences for the bird population and fishing industry. Extending our study in a dynamic ocean management framework to provide an early warning system to predict when short-tailed albatross might make close and longer encounters with fishing vessels could be the next step. Furthermore, our analysis methods to assess when, where and why marine animals interact with fishing vessels can be applied to many other marine species in order to understand and reduce conflicts with fisheries.
This blog was for the Global Fishing Watch blog at globalfishingwatch.org
References
Collet, J., Patrick, S. C., & Weimerskirch, H. (2015). Albatrosses redirect flight towards vessels at the limit of their visual range. Marine Ecology Progress Series, 526, 199–205. http://doi.org/10.3354/meps11233
Orben, RA J Adams, M Hester, SA Shaffer, R Suryan, T Deguchi, K Ozaki, F Sato, LC Young, C Clatterbuck, MG Conners, DA Kroodsma, LG Torres. 2021. Across borders: External factors and prior behavior influence North Pacific albatross associations with fishing vessels. Journal of Applied Ecology. https://besjournals.onlinelibrary.wiley.com/doi/10.1111/1365-2664.13849
Last week marked the one year anniversary of the pandemic reality we have all been living. It has been an extremely challenging year, with everyone experiencing different kinds of difficulties and hurdles. One challenge that likely unites the majority of us is having to forego seeing our loved ones. For me personally, this is the longest time I have not seen my family (445 days and counting) and I know I am not alone in this situation. My homesickness started a train of thought about cetacean parental care and inspired me to write a blog about this topic. As you can see from the title, this post focuses on maternal care, rather than parental care. This bias isn’t due to my lack of research on this topic or active exclusion, but rather because there are currently no known cetacean species where paternal participation in offspring production and development extends beyond copulation (Rendell et al. 2019). Thus, this blog is all about the role of mothers in the lives of cetacean calves.
Like humans, cetacean mothers invest a lot of energy into their offspring. Most species have a gestation period of 10 or more months (Oftedal 1997). For baleen whale females in particular, pregnancy is not an easy feat given that they only feed during summer feeding seasons. They must therefore acquire all of the energy they will need for two migrations, birth, and (almost) complete lactation, before they will have access to food on feeding grounds again. For pregnant gray whales, a mere 4% loss in average energy intake on the foraging grounds will prevent her from successfully producing and/or weaning a calf (Villegas-Amtmann et al. 2015), demonstrating how crucial the foraging season is for a pregnant baleen whale. Once a calf is born, lactation ensues, ranging in length between approximately 6-8 months for most baleen whale species to upwards of one or two years in odontocetes (Oftedal 1997). The very short lactation period in baleen whales is offset by the large volume (for blue whales, up to 220 kg per day) and high fat percentage (30-50%) of milk that mothers provide for their calves (Oftedal 1997). In contrast, odontocetes (or toothed whales) have a more prolonged period of lactation with less fatty milk (10-30%). This discrepancy in lactation period lengths is in part because odontocete species do not undertake long migrations, which allows females to feed year-round and therefore allocate energy to nursing young for a longer time.
Blue whale calf nursing in New Zealand in 2016. Footage captured via unmanned aerial system (UAS; drone) piloted by Todd Chandler for GEMM Lab’s OBSIDIAN project. Source: GEMM Lab.
Aside from the energetically costly task of lactation, cetacean mothers must also assist their calves as they learn to swim. Echelon swimming is a common position of mother-calf pairs whereby the calf is in very close proximity to its mother’s mid-lateral flank and provides calves with hydrodynamic benefits. Studies in bottlenose dolphins have shown that swimming in echelon results in a 24% reduction in mean maximum swim speeds and a 13% decrease in distance per stroke (Noren 2008) for mothers, while concurrently increasing average swim speeds and distance per stroke of calves by 28% and 19%, respectively (Noren et al. 2007). While these studies have only been conducted in odontocete species, echelon swimming is also observed in baleen whales (Smultea et al. 2017), indicating that baleen whale females may experience the same reductions in swimming efficiency. Furthermore, mothers will forgo sleep in the first days after birth (killer whales & bottlenose dolphins; Lyamin et al. 2005) and/or shorten their dive foraging times to accommodate calf diving ability (bottlenose dolphins [Miketa et al. 2018] & belugas [Heide-Jørgensen et al. 2001]). Females must endure these losses in foraging opportunities and decreased swimming efficiency when they are at their most nutritionally stressed to ensure the well-being and success of their offspring.
It is at the time of weaning (when a calf becomes independent), that we start to see differences in the maternal role between baleen and toothed whale mothers. Odontocetes have much stronger sociality than baleen whales causing offspring to stay with their mothers for much longer periods. Among the largest toothed whales, such as killer and sperm whales, offspring stay with their mothers in stable matrilineal units for often a lifetime. Among the smaller toothed whales, such as bottlenose dolphins, maternal kin maintain strong bonds in dynamic fission-fusion societies. In contrast, post-weaning maternal care in baleen whales is limited, with the mother-calf pair typically separating soon after the calf is weaned (Rendell et al. 2019).
Conceptual diagram depicting where baleen (Mysticeti) and toothed (Odontoceti) whales fall on the continuum of low to high social structure and matrilineal kinship structure. The networks at the top depict long-term datasets of photo-identified individuals (red nodes = females, blue nodes = males, yellow nodes = calves) with thickness of connecting lines representing strength of association between individuals. Figure and caption [adapted] from Rendell et al. 2019.
The long-term impact of social bonds in odontocetes is evident through examples of vertically transmitted behaviors (from mother to calf) in a number of species. For example, the use of three unique foraging tactics (sponge carrying, rooster-tail foraging, and mill foraging) by bottlenose dolphin calves in Shark Bay, Australia, was only significantly explained by maternal use of these tactics (Sargeant & Mann 2009). In Brazil, individuals of four bottlenose dolphin populations along the coast cooperatively forage with artisanal fishermen, which involves specialized and coordinated behaviors from both species. This cooperative foraging tactic among dolphins is primarily maintained across generations via social learning from mothers to calves (Simões-Lopeset al. 2016). The risky tactic of intentional stranding by killer whales on beaches to capture elephant seal pups requires a high degree of skill and high parental investment to reduce the associated risk of stranding (Guinet & Bouvier 1995).
Evidence for vertical transmission of specialized foraging tactics in baleen whales currently does not exist. Bubble-net feeding is a specialized tactic employed by humpback whales in three oceanic regions where multiple individuals work together to herd and trap prey (Wiley et al. 2011). However, it remains unknown whether this behavior is vertically transmitted. Simultaneous video tags from a mother-calf humpback whale pair in the Western Antarctic Peninsula documented synchrony in dives, with the calf’s track lagging behind the mother’s by 4.5 seconds, suggesting that the calf was following its mother (Tyson et al. 2012). Synchronous diving likely allows calves to observe their mothers and practice their diving, and could offer a pathway for them to mimic foraging behaviors and tactics displayed by mothers.
While there currently may not be evidence for vertical transmission of specialized foraging tactics among the baleen whales, there is documentation of matrilineal fidelity to both foraging (Weinrich 1998, Barendse et al. 2013, Burnham & Duffus 2020) and breeding grounds (Carroll et al. 2015). Matrilineal site fidelity to foraging grounds is not exclusive to baleen whales and has also been documented in a number of odontocete species (Palsbøll et al. 1997, Turgeon et al. 2012).
A gallery of some GEMM Lab documented mother-calf gray whale pairs. All images captured under NOAA/NMFS permit #21678. Exclamation (mom; left) and Angie (calf; right).
Scarlett (mom; left) and Brown (calf; right).
Clouds (mom; back) and Cheetah (calf; front).
Xena (mom; back) and Evie (calf; front).
Knife (mom; left) and Daffodil (calf; right).
Pristine (mom; front) and Three Stars (calf; back).
Triplet (mom; right) and White Knuckle (calf; left).
In the GEMM Lab, we are interested in exploring the potential long-term bonds, role and impact of Pacific Coast Feeding Group (PCFG) gray whale mothers on their calves. GEMM Lab PhD student Clara Bird is digging into whether specialized foraging tactics, such as bubble blasts and headstands, are passed down from mothers to calves. I hope to assess whether using the PCFG range as a foraging ground (rather than the Arctic region) is a vertically transmitted behavior or whether environmental factors may play a larger role in the recruitment and dynamics of the PCFG. It will take us a while to get to the bottom of these questions, so in the meantime hug your loved ones if it’s safe to do so or, if you’re in my boat, continue to talk to them virtually until it is safe to be reunited.
References
Barendse, J., Best, P. B., Carvalho, I., and C. Pomilla. 2013. Mother knows best: occurrence and associations of resighted humpback whales suggest maternally derived fidelity to a southern hemisphere coastal feeding ground. PloS ONE 8:e81238.
Burnham, R. E., and D. A. Duffus. 2020. Maternal behaviors of gray whales (Eschrichtius robustus) on a summer foraging site. Marine Mammal Science 36:1212-1230.
Carroll, E. L., Baker, C. S., Watson, M., Alderman, R., Bannister, J., Gaggiotti, O. E., Gröcke, D. R., Patenaude, N., and R. Harcourt. 2015. Cultural traditions across a migratory network shape the genetic structure of southern right whales around Australia and New Zealand. Scientific Reports 5:16182.
Guinet, C., and J. Bouvier. 1995. Development of intentional stranding hunting techniques in killer whale (Orcinus orca) calves at Crozet Archipelago. Canadian Journal of Zoology 73:27-33.
Heide-Jørgensen, M. P., Hammeken, N., Dietz, R., Orr, J., and P. R. Richard. 2001. Surfacing times and dive rates for narwhals and belugas. Arctic 54:207-355.
Lyamin, O., Pryaslova, J., Lance, V., and J. Siegel. 2005. Continuous activity in cetaceans after birth. Nature 435:1177.
Miketa, M. L., Patterson, E. M., Krzyszczyk, E., Foroughirad, V., and J. Mann. 2018. Calf age and sex affect maternal diving behavior in Shark Bay bottlenose dolphins. Animal Behavior 137:107-117.
Noren, S. R. 2008. Infant carrying behavior in dolphins: costly parental care in an aquatic environment. Functional Ecology 22:284-288.
Noren, S. R., Biedenbach, F., Redfern, J. V., and E. F. Edwards. 2007. Hitching a ride: the formation locomotion strategy of dolphin calves. Functional Ecology 22:278-283.
Oftedal, O. T. Lactation in whales and dolphins: evidence of divergence between baleen- and toothed-species. Journal of Mammary Gland Biology and Neoplasia 2:205-230.
Palsbøll, P. J., Heide-Jørgensen, M. P., and R. Dietz. 1996. Population structure and seasonal movements of narwhals, Monodon monoceros, determined from mtDNA analysis. Heredity 78:284-292.
Rendell, L., Cantor, M., Gero, S., Whitehead, H., and J. Mann. 2019. Causes and consequences of female centrality in cetacean societies. Philosophical Transactions of the Royal Society B 374:20180066.
Sargeant, B. L., and J. Mann. 2009. Developmental evidence for foraging traditions in wild bottlenose dolphins. Animal Behavior 78:715-721.
Simões-Lopes, P. C., Daura-Jorge, F. G., and M. Cantor. 2016. Clues of cultural transmission in cooperative foraging between artisanal fishermen and bottlenose dolphins, Tursiops truncatus (Cetacea: Delphinidae). Zoologia (Curitiba) 33:e20160107.
Smultea, M. A., Fertl, D., Bacon, C. E., Moore, M. R., James, V. R., and B. Würsig. 2017. Cetacean mother-calf behavior observed from a small aircraft off Southern California. Animal Behavior and Cognition 4:1-23.
Turgeon, J., Duchesne, P., Colbeck, G. J., Postma, L. D., and M. O. Hammill. 2011. Spatiotemporal segregation among summer stocks of beluga (Delphinapterus leucas) despite nuclear gene flow: implication for the endangered belugas in eastern Hudson Bay (Canada). Conservation Genetics 13:419-433.
Tyson, R. B., Friedlaender, A. S., Ware, C., Stimpert, A. K., and D. P. Nowacek. 2012. Synchronous mother and calf foraging behaviour in humpback whales Megaptera novaeangliae: insights from multi-sensor suction cup tags. Marine Ecology Progress Series 457:209-220.
Villegas-Amtmann, S., Schwarz, L. K., Sumich, J. L., and D. P. Costa. 2015. A bioenergetics model to evaluate demographic consequences of disturbance in marine mammals applied to gray whales. Ecosphere 6:1-19.
Weinrich, M. 1998. Early experience in habitat choice by humpback whales (Megaptera novaeaengliae). Journal of Mammalogy 79:163-170.
Wiley, D., Ware, C., Bocconcelli, A., Cholewiak, D., Friedlaender, A., Thompson, M., and M. Weinrich. 2011. Underwater components of humpback whale bubble-net feeding behavior. Behavior 148:575-602.
Clara Bird, PhD Student, OSU Department of Fisheries and Wildlife, Geospatial Ecology of Marine Megafauna Lab
When I started working on my thesis, I anticipated many challenges related to studying the behavioral ecology of gray whales. From processing five-plus years of drone footage to data analysis, there has been no shortage of anticipated and unexpected issues. I recently hit an unexpected challenge when I started video processing that piqued my interest. As I’ve discussed in a previous blog, ethograms are lists of defined behaviors that help us properly and consistently collect data in a standardized approach. Ethograms form a crucial foundation of any behavior study as the behaviors defined ultimately affect what questions can be asked and what patterns are detected. Since I am working off of the thorough ethogram of Oregon gray whales from Torres et al. (2018), I had not given much thought to the process of adding behaviors to the ethogram. But, while processing the first chunk of drone videos, I noticed some behaviors that were not in the original ethogram and struggled to decide whether or not to add them. I learned that ethogram development can lead down several rabbit holes. The instinct to try and identify every movement is strong but dangerous. Every minute movement does not necessarily need to be included and it’s important to remember the ultimate goal of the analysis to avoid getting bogged down.
Fundamental behavior questions cannot be answered without ethograms. For example, Baker et al. (2017) developed an ethogram for bottlenose dolphins in Ireland in order to conduct an initial quantitative behavior analysis. They did so by reviewing published ethograms for bottlenose dolphins, consulting with multiple experts, and revising the ethogram throughout the study. They then used their data to test inter-observer variability, calculate activity budgets, and analyze how the activity budgets varied across space and time.
Howe et al. (2015) also developed an ethogram in order to conduct quantitative behavior analyses. Their goals were to use the ethogram and subsequent analyses to better understand the behavior of beluga whales in Cook Inlet, AK, USA and to inform conservation. They started by writing down all behaviors they observed in the field, then they consolidated their notes into a formal ethogram that they used and refined during subsequent field seasons. They used their data to analyze how the frequencies of different behaviors varied throughout the study area at different times. This study served as an initial analysis investigating the effect of anthropogenic disturbance and was refined in future studies.
My research is similarly geared towards understanding behavior patterns to ultimately inform conservation. The primary questions of my thesis involve individual specialization, patterns of behavior across space, the relationship between behavior and body condition, and social behavior (check out this blog to learn more). While deciding what behaviors to add to my ethogram I’ve had to remind myself of these main questions and the bigger picture. The drone footage lets us see so much detail that it’s tempting to try to define every movement we can observe. One rabbit hole I’ve had to avoid a few times is locomotion. From the footage, it is possible to document fluke beats and pectoral fin strokes. While it could be interesting to investigate how different whales move in different ways, it could easily become a complicated mess of classifying different movements and take me deep into the world of whale locomotion. Talking through what that work would look like reminded me that we cannot answer every question and trying to assess all exciting side projects can cause us to lose focus on the main questions.
While I avoided going down the locomotion rabbit hole, there were some new behaviors that I did add to my ethogram. I’ll illustrate the process with the examples of two new behaviors I recently added: fluke swish and pass under (Clips 1 and 2). Clip 1 shows a whale rapidly moving its fluke to the side. I chose to add fluke swish because it’s such a distinct movement and I’m curious to see if there’s a pattern across space, time, individual, or nearby human activity that might explain its function. Clip 2 shows a calf passing under its mom. It’s not nursing because the calf doesn’t spend time under its mom, it just crosses underneath her. The calf pass under behavior could be a type of mom-calf tactile interaction. Analyzing how the frequency of this behavior changes over time could show how a calf’s dependency on its mom changes over as it ages.
In defining these behaviors, I had to consider how many different variations of this behavior would be included in the definition. This process involves considering at what point a variation of that behavior could serve a different function, even without knowing the function of the original behavior. For fluke swish this process involved deciding to only count a behavior as a fluke swish if it was a big, fast movement. A small and slow movement of the fluke a little to the side could serve a different function, such as turning, or be a random movement.
Clip 1: Fluke swish behavior (Video filmed under NOAA/NMFS research permit #16111 by certified drone pilot Todd Chandler).
Clip 2: Pass under behavior (Video filmed under NOAA/NMFS research permit #16111 by certified drone pilot Todd Chandler).
The next step involved deciding if the behavior would be a ‘state’ or ‘point’ event. A state event is a behavior with a start and stop moment; a point event is instantaneous and assigned to just a point in time. I would categorize a behavior as a state event if I was interested in questions about its duration. For example, I could ask “what percentage of the total observation time was spent in a certain behavior state?” A point event would be a behavior where duration is not applicable, but I could ask a question like “Did whale 1 perform more point event A than whale 2?”. Both fluke swish and pass under are point events because they only happen for an instant. In a pass under the calf is passing under its mom for just a brief point in time, making it a point event. The final step was to name the behavior. As I discussed in this blog, the name of the behavior does not matter as much as the definition but it is important that the name is clear and descriptive. We chose the name fluke swish because the fluke rapidly moves from side to side and pass under because the calf crosses under its mom.
Frankly, in the beginning, I was a bit overwhelmed by the realization that the content of my ethogram would ultimately control the questions I could answer. I could not help but worry that after processing all the videos, I would end up regretting not defining more behaviors. However, after reading some of the literature, chatting with Leigh, and reviewing the initial chunk of videos several times, I am more confidence in my judgment and my ethogram. I have accepted the fact that I can’t anticipate everything, and I am confident that the behaviors I need to answer my research questions are included. The process of reviewing and updating my ethogram has been a rewarding challenge that resulted in a valuable lesson that I will take with me for the rest of my career.
References
Baker, I., O’Brien, J., McHugh, K., & Berrow, S. (2017). An ethogram for bottlenose dolphins (Tursiops truncatus) in the Shannon Estuary, Ireland. Aquatic Mammals, 43(6), 594–613. https://doi.org/10.1578/AM.43.6.2017.594
Howe, M., Castellote, M., Garner, C., McKee, P., Small, R. J., & Hobbs, R. (2015). Beluga, Delphinapterus leucas, ethogram: A tool for cook inlet beluga conservation? Marine Fisheries Review, 77(1), 32–40. https://doi.org/10.7755/MFR.77.1.3
Torres, L. G., Nieukirk, S. L., Lemos, L., & Chandler, T. E. (2018). Drone up! Quantifying whale behavior from a new perspective improves observational capacity. Frontiers in Marine Science, 5(SEP). https://doi.org/10.3389/fmars.2018.00319
By Alejandro Fernandez Ajo, PhD student at the Department of Biology, Northern Arizona University, Visiting scientist in the GEMM Lab working on the gray whale physiology and ecology project
About four years ago, I was in Patagonia, Argentina deciding where to focus my research and contribute to whale conservation efforts. At the same time, I was doing fieldwork with the Whale Conservation Institute of Argentina at the “Whale Camp” in Península Valdés. I read tons of papers and talked with my colleagues about different opportunities and gaps in knowledge that I could tackle during my Ph.D. program. One of the questions that caught my attention was about the unknown cause (or causes) for the recurrent high calf mortalities that the Southern Right Whale (SRW) population that breeds at Peninsula Valdés experienced during the 2000s (Rowntree et al. 2013). Still, at that time, I was unsure how to tackle this research question.
Golfo San José, Península Valdés – Argentina. Collecting SRW behavioral data from the cliff’s vantage point. Source: A. Fernandez Ajo.
Between 2003 and 2013, at least 672 SRWs died, of which 91% were calves (Sironi et al. 2014). These mortalities represented an average total whale death per year of 80 individuals in the 2007-2013 period, which vastly exceeded the 8.2 average deaths per year of previous years by a ten-fold increase (i.e., 1993-2002) (Rowntree et al. 2013). In fact, this calf mortality rate was the highest ever documented for any population of large whales. During this period, from 2006 to 2009, I was the Coordinator of the Fauna Area in the Patagonian Coastal Zone Management Plan, and I collaborated with the Southern Right Whale Health Monitoring Program (AKA: The Stranding Program) that conducted field necropsies on stranded whales along the coasts of the Península and collected many different samples including whale baleen.
Southern Right Whale, found stranded in Patagonia Argentina. Source: Instituto de Conservación de Ballenas.
In this process, I learned about the emerging field of Conservation Physiology and the challenges of utilizing traditional approaches to studying physiology in large whales. Basically, the problem is that there is no possible way to obtain blood samples (the gold standard sample type for physiology) from free-swimming whales; whales are just too large! Fortunately, there are currently several alternative approaches for gathering physiological information on large whales using a variety of non-lethal and minimally invasive (or non-invasive) sample matrices, along with utilizing valuable samples recovered at necropsy (Hunt et al. 2013). That is how I learned about Dr. Kathleen Hunt’s novel research studying hormones from whale baleen (Hunt et al., 2018, 2017, 2014). Thus, I contacted Dr. Hunt and started a collaboration to apply these novel methods to understand the case of calf mortalities of the SRW calves in Patagonia utilizing the baleen samples that we recovered with the Stranding program at Península Valdés (see my previous blog post).
What is conservation physiology?
Conservation physiology is a multidisciplinary field of science that utilizes physiological concepts and tools to understand underlying mechanisms of disturbances to solve conservation problems. Conservation physiology approaches can provide sensitive biomarkers of environmental change and allow for targeted conservation strategies. The most common Conservation Physiology applications are monitoring environmental stressors, understanding disease dynamics and reproductive biology, and ultimately reducing human-wildlife conflict, among other applications.
I am now completing the last semester of my Ph.D. program. I have learned much about the amazing field of Conservation Physiology and how much more we need to know to achieve our conservation goals. I am still learning, yet I feel that through my research I have contributed to understanding how different stressors impact the health and wellbeing of whales, and about aspects of their biology that have long been obscured or unknown for these giants. One contribution I am proud of is our recent publication of, “A tale of two whales: putting physiological tools to work for North Atlantic and southern right whales,” which was published in January 2021 as a book chapter in “Conservation Physiology: Applications for Wildlife Conservation and Management” published by Oxford University Press: Oxford, UK.
This book outlines the significant avenues and advances that conservation physiology contributes to the monitoring, management, and restoration of wild animal populations. The book also defines opportunities for further growth in the field and identifies critical areas for future investigation. The text and the contributed chapters illustrate several examples of the different approaches that the conservation physiology toolbox can tackle. In our chapter, “A tale of two whales,” we discuss developments in conservation physiology research of large whales, with the focus on the North Atlantic right whale (Eubalaena glacialis) and southern right whale (Eubalaena australis), two closely related species that differ vastly in population status and conservation pressures. We review the advances in Conservation Physiology that help overcome the challenges of studying large whales via a suite of creative approaches, including photo-identification, visual health assessment, remote methods of assessing body condition, and endocrine research using non-plasma sample types such as feces, respiratory vapor, and baleen. These efforts have illuminated conservation-relevant physiological questions for both species, such as discrimination of acute from chronic stress, identification of likely causes of mortality, and monitoring causes and consequences of body condition and reproduction changes.
This book provides an overview of the different applications of Conservation Physiology, outlining the significant avenues and advances by which conservation physiology contributes to the monitoring, management, and restoration of wild animal populations. By using a series of global case studies, contributors illustrate how approaches from the conservation physiology toolbox can tackle a diverse range of conservation issues, including monitoring environmental stress, predicting the impact of climate change, understanding disease dynamics, and improving captive breeding, and reducing human-wildlife conflict. The variety of taxa, biological scales, and ecosystems is highlighted to illustrate the far-reaching nature of the discipline and allow readers to appreciate the purpose, value, applicability, and status of the field of conservation physiology. This book is an accessible supplementary textbook suitable for graduate students, researchers, and practitioners in conservation science, ecophysiology, evolutionary and comparative physiology, natural resources management, ecosystem health, veterinary medicine, animal physiology, and ecology.
References
Hunt KE, Fernández Ajó A, Lowe C, Burgess EA, Buck CL. 2021. A tale of two whales: putting physiological tools to work for North Atlantic and southern right whales. In: “Conservation Physiology: Integrating Physiology Into Animal Conservation And Management”, ch. 12. Eds. Madliger CL, Franklin CE, Love OP, Cooke SJ. Oxford University press: Oxford, UK.
Sironi, M., Rowntree, V., Di Martino, M. D., Beltramino, L., Rago, V., Franco, M., and Uhart, M. (2014). Updated information for 2012-2013 on southern right whale mortalities at Península Valdés, Argentina. SC/65b/BRG/06 report presented to the International Whaling Commission Scientific Committee, Portugal. <https://iwc.int/home>.
Rowntree, V.J., Uhart, M.M., Sironi, M., Chirife, A., Di Martino, M., La Sala, L., Musmeci, L., Mohamed, N., Andrejuk, J., McAloose, D., Sala, J., Carribero, A., Rally, H., Franco, M., Adler, F., Brownell, R. Jr, Seger, J., Rowles, T., 2013. Unexplained recurring high mortality of southern right whale Eubalaena australis calves at Península Valdés, Argentina. Marine Ecology Progress Series, 493, 275-289. DOI: 10.3354/meps10506
Hunt KE, Moore MJ, Rolland RM, Kellar NM, Hall AJ, Kershaw J, Raverty SA, Davis CE, Yeates LC, Fauquier DA, et al., 2013. Overcoming the challenges of studying conservation physiology in large whales: a review of available methods. Conserv Physiol 1: cot006–cot006.
Hunt, K.E., Stimmelmayr, R., George, C., Hanns, C., Suydam, R., Brower, H., Rolland, R.M., 2014. Baleen hormones: a novel tool for retrospective assessment of stress and reproduction in bowhead whales (Balaena mysticetus). Conserv. Physiol. 2, cou030. https://doi.org/10.1093/conphys/cou030
Hunt, K.E., Lysiak, N.S., Moore, M.J., Rolland, R.M., 2016. Longitudinal progesterone profiles in baleen from female North Atlantic right whales (Eubalaena glacialis) match known calving history. Conserv. Physiol. 4, cow014. https://doi.org/10.1093/conphys/cow014
Over the roughly 2.5 years that I have researched the Pacific Coast Feeding Group (PCFG) of gray whales, I have thought more and more about what makes a population, a population. From a management standpoint, the PCFG is currently not considered a separate population or even a sub-population of the Eastern North Pacific (ENP) gray whales. Rather, the PCFG is most commonly referred to as a ‘sub-group’ of the ENP. In my opinion, there are valid arguments both for and against the PCFG being designated as its own population. I will address those arguments briefly at the end of this post, but first, I want you to join on me on a journey that is tangential to my question of ‘what makes a population, a population?’ and one that started at the last Marine Mammal Institute Monthly Meeting (MMIMM).
During 2021’s first MMIMM, our director Dr. Lisa Ballance proposed that we lengthen our monthly meeting duration from 1 hour to 1.5 hours. The additional 30 minutes was to allow for an open-ended, institute-wide discussion of a current hot topic in marine mammal science. This proposal was immediately adopted, and the group dove into a discussion about the discovery of a new baleen whale species in the northeastern Gulf of Mexico: Rice’s whale (Balaenoptera ricei). Let me pause here very briefly to reiterate – the discovery of a new baleen whale species!! The fact that anything as large as 12 m could remain undiscovered in our oceans is really quite fascinating and shows that our scientific quest will likely never run out of discoverable subjects. Anyway, the discovery of this new species is supported by several lines of evidence. Unfortunately (but understandably), MMI’s discussion of these topics had to cease after 30 minutes, however I had more questions. I wanted to know what had sparked the researchers to believe that they had discovered a new cetacean species.
Scientific illustration of Bryde’s whale. Source: NOAA Fisheries.
I started my research by skimming through some news articles about the Rice’s whale discovery. In a Smithsonian Magazine article, I saw a quote by Dr. Patricia Rosel, the lead author of the study detailing Rice’s whale, that read: “But we didn’t have a skull.”. That quote made me pause. A skull? Is that what it takes to discover and establish a new species? This desired piece of evidence seemed rather puzzling and a little antiquated to me, given that the field of genetics is so advanced now and since it is no longer an accepted practice to kill a wild animal just to study it (i.e., scientific whaling). I backtracked through the article to learn that in the 1990s, renowned marine mammal scientist Dale Rice (after whom Rice’s whale was named) recognized that a small population of baleen whale occurred in the northeastern part of the Gulf of Mexico year-round. At the time, this population was believed to be a sub-population of Bryde’s whale. It wasn’t until 2008, that NOAA scientists were able to conduct a genetic analysis of tissue samples from this population, only to find that these whales were genetically distinct from other Bryde’s whales (Rosel & Wilcox 2014). Yet, this information was not enough for these whales to be established as their own species. A skull really was needed to prove that these whales were in fact a new species. Thankfully (for the scientists) but sadly (for the whale), one of these individuals stranded in Sandy Key, Florida, in 2019, and a dedicated team of stranding responders from Florida Fish and Wildlife, Mote Marine Lab, NOAA, Dolphins Plus, and Marine Animal Rescue Society worked tirelessly in difficult conditions to comprehensively document and preserve this animal. Through the diligent work of this, and previous, stranding response teams, Dr. Rosel and her team were provided the opportunity to examine the skull needed to determine population-status. The science team determined that the bones atop the skull around the blowhole provided evidence that these whales were not only genetically, but also anatomically, different from Bryde’s whales. It was this incident, triggered by that short quote in the Smithsonian article, that brought me to my journey of asking ‘what makes a species, a species?’.
Given that I had just read that Dr. Rosel needed a skull to establish Rice’s whale as its own species, I assumed that my search for ‘how to establish a new species’ would end quickly in me finding a list of requirements, one of which would be ‘must present anatomical/skeletal evidence’. To my surprise, my search did not end quickly, and I did not find a straightforward list of requirements. Instead, I discovered that my question of ‘what makes a species, a species?’ does not have a black-and-white answer and involves a lot of debate.
The skull of this stranded whale was a large piece of evidence in establishing Rice’s whale as its own species. Source: Smithsonian Magazine from NOAA / Florida Fish and Wildlife Commission
Kevin de Queiroz, a vertebrate, evolutionary, and systematic biologist who has published extensively on theoretical and conceptual topics in systematic and evolutionary biology, believes that the issue of species delimitation (‘what makes a species, a species?’) has been made more complicated by a larger issue involving the concept of species itself (‘what is a species?’) (De Queiroz 2007). To date, there are 24 recognized species concepts (Mayden 1997). In other words, there are 24 different definitions of what a species is. Perhaps the most common example is the biological species concept where a species is defined as a group of individuals that are able to produce viable and fertile offspring following natural reproduction. Another example is the ecological species concept whereby a species is a group of organisms adapted to a particular set of resources and conditions, called a niche, in the environment. Problematically, many of these concepts are incompatible with one another, meaning that applying different concepts leads to different conclusions about the boundaries and numbers of species in existence (De Queiroz 2007).
This large number of species concepts is due to the different interests of certain subgroups of biologists. For example, highlighting morphological differences between species is central to paleontologists and taxonomists, whereas ecologists will focus on niche differences. Population geneticists will attribute species differences to genes, while for systematists, monophyly will be paramount. It goes on and on. And so does the debate about the concept of species. It seems that there currently is not one clear, defined consensus on what a species is. Some biologists argue that a species is a species if it is genetically different, while others will insist that skeletal and morphological evidence must be present. From what I can tell, it seems that scientists describe and (attempt to) establish a new species by publishing their lines of evidence, after which experts in the field discuss and evaluate whether a new species should be established.
In the field of marine mammal science, the Society of Marine Mammalogy’s Taxonomic Committee is charged with maintaining a standard, accepted classification and nomenclature of marine mammals worldwide. The committee annually considers and evaluates new, peer-reviewed literature that proposes changes (including additions) to marine mammal taxonomy. I expect that the case of Rice’s whale will be on the committee’s docket this year. Given that Rosel and co-authors presented geographic, morphological, and genetic evidence to support the establishment of Rice’s whale, I would not be surprised if the committee adds it to their curated list.
After taking this dive into the ‘what makes a species, a species?’ question, let’s see if we can apply some of what we’ve learned to the ‘what makes a population, a population?’ question regarding the PCFG and ENP gray whales. Following the ecological species concept, an argument for the recognition of the PCFG as its own population would be that they occupy an entirely different environment during their summer foraging season than the ENP whales. Not only are the geographic ranges different, but PCFG whales also show behavioral differences in their foraging tactics and targeted prey. The argument against the PCFG being classified as its own population is largely supported by genetic analysis that has revealed ambiguous evidence that the PCFG and ENP are not genetically isolated from one another. While one study has shown that there is maternal cultural affiliation within the PCFG (meaning that calves born to PCFG females tend to return to the PCFG range; Frasier et al. 2011), another has revealed that mixing between ENP and PCFG gray whales on the breeding grounds does occur (Lang et al. 2014). So, even though these two groups feed in areas that are very far apart (ENP: Arctic vs PCFG: US & Canadian west coast) and certain individuals do show a propensity for a specific feeding ground, the genetic evidence suggests that they mix when on their breeding grounds in Baja California, Mexico. Depending on which species concept you align with, you may see better arguments for either side.
PCFG gray whale along the Oregon coast during the GEMM Lab’s 2020 GRANITE summer field season. Image captured under NOAA/NMFS permit #21678. Source: GEMM Lab.
You may be wondering why it is important to even ponder questions like ‘what makes a species, a species?’ and ‘what makes a population, a population?’. Does it really matter if the PCFG are considered their own population? Would anything really change? The answer is, most likely, yes. If the PCFG were to be recognized as their own population, it would likely have an immediate effect on their conservation status and subsequently on how the population needed to be managed. Rather than being under the umbrella of a large, (mostly) stable population of ~25,000 individuals, the PCFG would consist of only ~250 individuals. A group this small would possibly be considered “endangered”, which would require much stricter monitoring and management to ensure that their numbers did not decline from year to year, especially due to anthropogenic activities.
For a long time, I felt like taxonomy was a bit of an archaic scientific field. In my mind, it was something that biologists had focused their time and energy on in the 18th century (most notably Carl Linnaeus, whose taxonomic classification system is still used today), but something that many biologists have moved on from focusing on in the 21st century. However, as I have developed and grown over the last years as a scientist, I have learned that scientific disciplines are often heavily intertwined and co-dependent on one another. As a result, I am able to see the enormous value and need for taxonomic work as it plays a large part in understanding, managing, and ultimately, conserving species and populations.
Literature cited
De Queiroz, K. 2007. Species concepts and species delimitation. Systematic Biology 56(6):879-886.
Frasier, T. R., Koroscil, S. M., White, B. N., and J. D. Darling. 2011. Assessment of population substructure in relation to summer feeding ground use in the eastern North Pacific gray whale. Endangered Species Research 14:39-48.
Lang, A. R., Calambokidis, J., Scordino, J., Pease, V. L., Klimek, A., Burkanov, V. N., Gearin, P., Litovka, D. I., Robertson, K. M., Mate, B. R., Jacobsen, J. K., and B. L. Taylor. 2014. Assessment of genetic structure among eastern North Pacific gray whales on their feeding grounds. Marine Mammal Science 30(4):1473-1493.
Mayden, R. L. 1997. A hierarchy of species concepts: the denouement of the species problem in The Units of Biodiversity – Species in Practice Special Volume 54 (M. F. Claridge, H. A. Dawah, and M. R. Wilson, eds.). Systematics Association.
Rosel, P. E., and L. A. Wilcox. 2014. Genetic evidence reveals a unique lineage of Bryde’s whale in the northeastern Gulf of Mexico. Endangered Species Research 25:19-34.
As we all try to find ways to be together safely this winter, the GEMM Lab has started a fun series of virtual movie nights. Just before the holidays, we watched “Big Miracle,” which tells the story of the historic whale entrapment event in Utqiagvik, Alaska (formerly called “Barrow”) that captured the world’s attention.
The 2012 film stars Drew Barrymore, who plays a Greenpeace activist, and John Krasinski, a television reporter covering the story.
In late September 1988, three gray whales became trapped in the sea ice just off Point Barrow. Local attempts to free the whales quickly became national news that captured the attention of millions, including President Ronald Reagan, pop legend Michael Jackson – and elementary-schooler Leigh Torres.
After the movie, Leigh told us about how she had religiously followed television updates on the rescue as a child. Hearing her memories of the event and its part in inspiring her to pursue a career in whale research was one of the best parts of watching the movie together as a lab.
Tuning in from my parents’ house in Fairbanks, Alaska, the story felt surprisingly close to home for me too. I had never heard Inupiaq spoken in a feature film before, and I was stunned to recognize the landscape around Utqiaġvik and realize that some of the movie was filmed on location. It was also the first movie I’d seen represent the myriad of human dimensions that surround whale research and policy, including Indigenous rights, oil and fishing industry interests, and environmental perspectives.
Certain elements of the movie also made me uncomfortable, and thus made me wonder about the movie’s accuracy. Why were the main characters in the film people from outside Alaska? How did the rescue logistics and decision-making processes really play out in Utqiaġvik? Why did the whales become trapped in the first place?
I was curious to learn more about the whales, and how Utqiaġvik experienced both the massive rescue effort and the Hollywood-ized retelling of its story. During a great Zoom conversation, I learned more from Craig George, a whale biologist who has worked in Utqiaġvik since the 1970s and was involved during the entire 1988 rescue mission.
Like all Hollywood movies based on real events, “Big Miracle” mixes facts with a healthy dose of fiction and storytelling. The movie portrays the three entrapped whales as a family unit, given the names Wilma, Fred, and Bam Bam. Craig described them in more scientific terms – three subadult gray whales, all 25-30 feet in length. He and the other biologists onsite collected data throughout the three-week rescue effort, recording the whales’ behavior, dive times, and vocalizations. They calculated that the whales’ respiration rates were double that of typical rates, revealing the whales’ distress.
The rescue team named the whales Crossbeak, Bone, and Bonnet based on each individual’s notable morphological traits. Photo: Craig George
“The community effort to free the whales was amazing,” Craig said. “Low-tech approaches and local knowledge are typically most effective in the Arctic, and all the best ideas relied on the Inupiaq knowledge of the area.”
With the aim of leading the whales offshore to safer waters, a team of volunteers cut a series of breathing holes at regular intervals in the sea ice. The approach seemed to work well, and so the ice-breaking crew was puzzled when the whales stopped using the new holes – until they realized the area was underlain by shoals that the whales were unwilling to cross. They began cutting in a new direction, and the whales appeared in the new hole instantly, before the opening was even completed.
“The whales were trying to tell us the direction they wanted to go,” Craig said. “It was really astonishing, because there was definitely a dynamic between us. We tried to train them to work with us, and they also trained us.”
A team of volunteers cut holes in the sea ice, creating a path to open water, while journalists document the moment. Photo: Craig George
Over three weeks, the rescue effort grew from local to international. Companies donated chainsaws and fuel, and people following the news outside Alaska flew to Utqiaġvik to volunteer their help. Several attempts to break the ice, including an ice-based pontoon tractor and an ice-breaking helicopter, failed. Working around the clock, and in temperatures below -20F, volunteers continued cutting breathing holes in the ice for the whales.
Finally, one hurdle remained between the whales and open water – a massive pressure ridge of grounded sea ice, about 20 ft high and just as deep. It was impossible to cut through with chainsaws. Two Russian icebreakers, the Vladimir Arseniev and the Admiral Makarov were enlisted to come break the ridge and clear the way to open water – no small diplomatic feat during the Cold War.
Ultimately, Craig said, the real story’s ending isn’t quite as picture-perfect as the one in “Big Miracle” – no oneactually knows whether the whales made it out or not.
“We know that the whales swam out the icebreaker track, because their blood was found on ice shards,” he said. “They might have made it out, but we never saw them again and don’t know for sure.”
This map shows the path of holes cut through the sea ice, icebreaker track, and pressure ridge of ice. “Barrow” is the former name of Utqiaġvik. Source: Geoff Carroll and Craig George
Nearly 40 years later, Craig says the story still comes up often in Utqiaġvik, but in a different context – climate change. In 1988, the sea ice froze up in late September. In 2020, however, there was no shore-fast ice until early December. Craig remembers that, during the rescue, temperatures dropped to -24°F one night — colder than Utqiaġvik had experienced yet in January 2020, when we last spoke. Today’s dramatically different conditions have impacts for the entire Arctic ecosystem, as well as the people who rely on it to survive.
Watching “Big Miracle” sparked so many questions about the past, and talking with Craig gave me just as many questions about the future. How will changing ocean conditions impact gray whales, and other Arctic whales? How will the social and environmental dynamics that “Big Miracle” depicted – environmentalism, resource exploitation, and Indigenous rights – adapt and evolve in a changing Arctic? What will the Alaskan Arctic look like in another 40 years?
2PhD Student, Northwestern University Department of Economics
The Greek word “oikos” refers to the household and serves as the root of the words ecology and economics. Although perhaps surprising, the common origin reflects a shared set of basic questions and some shared theoretical foundations related to the study of how lifeforms on earth use scarce resources and find equilibrium in their respective “households”. Early ecological and economic theoretical texts drew inspiration from one another in many instances. Paul Samuelson, fondly referred to as “the father of modern economics,” observed in his defining work Foundations of Economic Analysis that the moving equilibrium in a market with supply and demand is “essentially identical with the moving equilibrium of a biological or chemical system undergoing slow change.” Likewise, early theoretical ecologists recognized the strength of drawing on theories previously established in economics (Real et al. 1991). Similar broad questions are central to researchers in both fields; in a large and dynamic system (termed “macro” in economics) scale, ecologists and economists alike work to understand where competitive forces find equilibrium, and an in individual (or micro) scale, they ask how individuals make behavior choices to maximize success given constraints like time, energy, wealth, or physical resources.
The central model economists have in mind when trying to understand human choices involves “constrained optimization”: what decision will maximize a person, family, firm, or other agent’s objectives given their limitations? For example, someone that enjoys relaxing but also seeks a livable income must choose how much time to devote to working versus relaxing, given the constraint of having just 24 hours in the day, and given the wage they receive from working. An economist studying this decision may want to learn about how changes in the wage will affect that person’s choice of working hours, or how much they dislike working relative to relaxing. Along similar lines, early ecologists theorized that organisms could be selected for one of two optimization strategies: minimizing the time spent acquiring a given amount of energy (i.e., calories from food), or maximizing total energy acquisition per unit of time (Real et al. 1991). Foundational work in the field of economics clarified numerous technical details about formulating and solving such optimization problems. Returning to the example of the leisure time decision, economic theory asks: does it matter if we model this decision as maximizing income given wages and limited time, or as minimizing hours spent working given a desired lifetime income?; can we formulate a “utility function” that describes how well-off someone is with a given income and amount of leisure?; can we solve for the optimal amount of leisure with pen and paper? The toolkit arising from this work serves as a jumping off point for all contemporary economic research, and the kinds of choices understood under this framework is vast, from, where should a child attend school?; to, how should a government allocate its budget across public resources?
Early work in ecology drew from foundational concepts in economics, following the realization that the strategies by which organisms exploit resources most efficiently also involve optimization. This parallel was articulated by MacArthur and Pianka in their foundational 1966 paper Optimal Use of a Patchy Environment, in which they state: “In this paper we undertake to determine in which patches a species would feed and which items would form its diet if the species acted in the most economical fashion. Hopefully, natural selection will often have achieved such optimal allocation of time and energy expenditures.” Subsequently, this idea was refined into what is known in ecology as the marginal value theorem, which states that an animal should remain in a prey patch until the rate of energy gain drops below the expected energy gain in all remaining available patches (Charnov 1976). In other words, if it is more profitable to switch prey patches than to stay, an animal should move on. These optimization models therefore allow ecologists to pose specific evolutionary and behavioral hypotheses, such as examining energy acquisition over time to understand selective forces on foraging behavior.
As the largest animals on the planet, blue whales have massive prey requirements to meet energy demands. However, they must balance their need to feed with costs such as oxygen consumption during breath-holding, the travel time it takes to reach prey patches at depth, the physiological constraints of diving, and the necessary recuperation time at the surface. It has been demonstrated that blue whales forage selectively to optimize this energetic budget. Therefore, blue whales should only feed on krill aggregations when the energetic gain outweighs the cost (Fig. 1), and this pattern has been empirically demonstrated for blue whale populations in the Gulf of St. Lawrence, Canada (Doniol-Valcroze et al. 2011), in the California Current, (Hazen et al. 2015) and in New Zealand (Torres et al. 2020).
Figure 1. Figure reprinted from Hazen et al. 2015, illustrating how a blue whale should theoretically optimize foraging success in two scenarios. Energy gained from feeding is shown by the blue lines, whereas the cost of foraging in terms of declining oxygen stores during a dive is illustrated by the red lines. On the left (panel B), the whale maximizes its energy gain by increasing the number of feeding lunges (shown by black circles) at the expense of declining oxygen stores when prey density is high. On the right (panel C), the whale minimizes oxygen use by reducing the number of feeding lunges when prey density is low.
The notion of the marginal value theorem is likewise at work in countless economic settings. Economic theory predicts that a farmer cultivating two crops would allocate resources into each crop such that the returns to adding more resources into each crop are the same. If not, she should move resources from the less productive crop to the one where marginal gains are larger. A fisherman, according to this notion, continues to fish longer into the season until the marginal value of one additional day at sea equals the marginal cost of their time, effort, and expenses. These predictions are intuitive by the same logic as the blue whale choosing where to forage, and derive from the mathematics of constrained and unconstrained optimization. Reassuringly, empirical work finds evidence of such profit-maximizing behavior in many settings. In a recent working paper, Burlig, Preonas, and Woerman explore how farmers’ water use in California responds to changes in the price of electricity, which effectively makes groundwater irrigation more expensive due to electric pumping. They find that farmers are very responsive to these changes in marginal cost. Farmers achieve this reduction in water use predominantly by switching to less water-intensive crops and fallowing their land (Burlig, Preonas, and Woerman 2020).
Undoubtedly there are fundamental differences between an ecosystem with interacting biotic and abiotic components and the human-economic environment with its many social and political structures. But for certain types of questions, the parallels across the shared optimization problems are striking. The foundational theories discussed here have paved the way for subsequent advances in both disciplines. For example, the field of behavioral ecology explores how competition and cooperation between and within species affects fitness of populations. Reflecting on early seminal work lends some perspective on how an area of research has evolved. Likewise, exploring parallels between disciplines sheds light on common threads, in turn revealing insights into each discipline individually.
References:
Burlig, Fiona, Louis Preonas, and Matt Woerman (2020). Groundwater, energy, and crop choice. Working Paper.
Charnov EL (1976) Optimal foraging: The marginal value theorem. Theoretical Population Biology 9:129–136.
Doniol-Valcroze T, Lesage V, Giard J, Michaud R (2011) Optimal foraging theory predicts diving and feeding strategies of the largest marine predator. Behavioral Ecology 22:880–888.
Hazen EL, Friedlaender AS, Goldbogen JA (2015) Blue whales (Balaenoptera musculus) optimize foraging efficiency by balancing oxygen use and energy gain as a function of prey density. Science Advces 1:e1500469–e1500469.
MacArthur RH, Pianka ER (1966) On optimal use of a patchy environment. The American Naturalist 100:603–609.
Real LA, Levin SA, Brown JH (1991) Part 2: Theoretical advances: the role of theory in the rise of modern ecology. In: Foundations of ecology: classic papers with commentaries.
Samuelson, Paul (1947). Foundations of Economic Analysis. Harvard University Press.
Torres LG, Barlow DR, Chandler TE, Burnett JD (2020) Insight into the kinematics of blue whale surface foraging through drone observations and prey data. PeerJ 8:e8906.