As I sit down at my desk during the first week of 2022 to write the first blog of this new year, more than ever before I feel like I am at a pivotal time. Standing in front of an invisible frontier, contemplating the past, and anxiously looking ahead.
Globally, 2021 was yet another challenging year. The COVID pandemic is persisting in endless waves of contamination and new variants. Climate change is all the more on our minds as the COP26 failed to live up to the expectations of many.
For me personally, 2021 was a very strange year too. I recovered from an accident I had in November 2020 that shook me to the bones and pushed me into living life to its fullest. On the other hand, the pandemic prevented me from moving to Oregon and I have been remotely working on the OPAL project for a year. I feel very lucky to participate in this work and I have enjoyed every bit of time I have spent on my computer processing data and teasing out the ecological drivers of whale distribution in Oregon. Yet, despite the numerous zoom meeting and email exchanges, I have been frustrated by the long-distance relationship I had with my dear GEMM lab colleagues and friends. Like so many others, I have felt the tow of the virtual life the pandemic has imposed on us.
As I reflect on the mixed feelings I am experiencing in this first week of 2022, I realize that the global context we live in and my individual questionings are intertwined. The pandemic and environmental issues triggered the same ethical and philosophical questions about individual responsibility, freedom, and equity. For instance, why should I make sacrifices that will cost me a lot personally but only have a very minor effect on the broader scale? The year 2021 has confronted us with a harsh reality: however strongly you believe your answer to the above question is the right one, other people might think otherwise.
The term eco-anxiety has emerged in recent years to describe people suffering from ‘persistent worries about the future of Earth and the life it shelters’. These symptoms of chronic fear are rising worldwide, which sadly but frankly, is only normal given that the degradation of our climate and biosphere deserves our full attention. More disturbingly, I found out that eco-anxiety is mostly affecting children and young people around the globe. Despite acting for the environment on an everyday basis and working as a conservation biologist, I can relate to this feeling of overwhelming helplessness.
In the first week of this new year, I would like to turn this distress into motivation to act and do better. To that extent, ‘adaptation’ is the word that keeps coming up to my mind. In biology, adaptation is the process of change by which an organism or species becomes better suited to its environment. Contrary to ‘acclimation’ that refers to a temporary change occurring on the short term, adaptation is a more profound evolution occurring at the scale of multiple generations. Somewhat, we need to combine the best of both worlds, adapt profoundly but adapt fast.
As I stayed at my family house in Toulouse (France) during the last couple weeks, I went through my old stuff in the room I occupied as a teenager and found a note book written by a 13 year-old Solène. I smiled at my words “One day, I will become a Biologist so that maybe I can save our beautiful planet, […] it’s the only thing that matters”. I was both impressed by the strength of the conviction I was holding to back then and stunned that I have now reached a place, as an independent adult and early career marine ecologist, where I could actually put these words in action.
So here is my 2022 New Year’s resolution: despite the waves of anxiety that sometimes hit us, let’s keep fighting our battles and trust that we can make this world a better place!
“Sometimes you have the feeling that nothing makes sense anymore, and sometimes it just feels right.” A picture of myself taken during a research cruise in New Caledonia this summer. We were searching for humpback whales in the Chesterfield archipelago (South Pacific), one of the most remote and pristine reef in the world (Photo credit: Marine Reveilhac, mission MARACAS/IRD/Opération Cétacés/WWF/GouvNC/Parc naturel de la mer de Corail).
Another year has come and gone, and the GEMM Lab has expanded in many facets! Every year it gets just a little bit harder to succinctly summarize all of the research, outreach, and successes that the GEMMs accomplish but it is an absolute honor and thrill to be a member of this lab. So, please enjoy the 6th edition of a GEMM Lab Year in the Life!
Our lab has almost doubled in size since I wrote the 2020 edition of this blog! This year we welcomed the arrival of two postdocs (Drs. Alejandro [Ale] Fernández Ajó and KC Bierlich) and two Master’s students (Allison Dawn and Miranda Mayhall). Ale and KC joined us as freshly minted Drs., as Ale defended his doctoral thesis from Northern Arizona University in April, while KC graduated in May from Duke University. Both of them immediately jumped into GRANITE fieldwork, scooping gray whale poop and flying drones (more below). Allison also dove headfirst into gray whale fieldwork as she co-led the TOPAZ and JASPER projects with me (Lisa) after defending her undergraduate thesis and graduating from the University of North Carolina with highest honors in the spring. Miranda, a U.S. Army Intelligence veteran, also joins us from the East Coast as she moved from Virginia to Oregon with her 10-year-old daughter, Mia, and two dogs, Angus and Mr. Gibbs. Unlike our other new arrivals, Miranda’s research does not relate to gray whales as she is part of the GEMM Lab’s newest research project…
There are exciting developments in the research project realm of the GEMM Lab every year. This year’s new project, HALO (Holistic Assessment of Living marine resources off Oregon), is particularly exciting as it is a joint project with the Cornell Lab, with GEMM Lab PI Leigh collaborating with Dr. Holger Klinck to better understand cetacean distributions off Oregon. HALO will involve monthly survey cruises aboard MMI’s R/V Pacific Storm along the Newport Hydrographic line (65 nm to 5 nm off Newport), where three Rockhopper hydrophones have been deployed and are passively monitoring cetacean acoustics. The HALO team, which includes GEMM students Miranda and PhD candidates Dawn Barlow and Rachel Kaplan, has already had two successful cruises this year! Check out the HALO website to stay tuned for updates throughout 2022. In addition to starting new research projects in our Oregon backyard, the GEMM Lab has also ventured further north, to the more frigid waters of Kodiak, Alaska. Postdocs KC and Ale went on a scouting mission to Kodiak Island to see whether the multidisciplinary methods we use in the GRANITE project to study PCFG gray whales in Oregon, can also be applied to other gray whales in other study areas. The reconnaissance trip was a huge success with KC and Ale making vital connections with potential collaborators and managing to collect some pilot data (drone flights, prey samples, and one fecal sample!). Both of these new ventures are funded by sales and renewals of the special Oregon gray whale license plate, which benefits MMI. We gratefully thank all the gray whale license plate holders, who made this research possible, and encourage any Oregonians that don’t have a whale on their tale yet, to do so in 2022!
These new research ventures certainly do not mean that we neglected our already established field research projects – in fact, most of them have flourished and thrived this year! Rachel and Dawn returned as marine mammal observers to the R/V Shimada for the May stint of the Northern California Current research cruise. They observed Dall’s porpoise, Northern right whale & Pacific white-sided dolphins, as well as killer, humpback, & fin whales. These sightings will add to the growing OPAL (Overlap Predictions About Large whales) dataset that both Rachel and postdoc Solène Derville are analyzing to better understand whale distribution patterns in Oregon waters. Speaking of OPAL, MMI Faculty Research Assistant Craig Hayslip and Leigh continued to take to the skies in U.S. Coast Guard helicopters to obtain monthly cetacean distribution data, which is also being used in the OPAL project to identify the co-occurrence between whales and fishing effort in Oregon to reduce entanglement risk. Both of our gray whale projects, GRANITE (Gray whale Response to Ambient Noise Informed by Technology & Ecology) and TOPAZ (Theodolite Overlooking Predators & Zooplankton)/JASPER (Journey for Aspiring Scientists Pursuing Ecological Research) had another year of successful field seasons. The GRANITE team, which includes Leigh, Todd Chandler, Ale, KC, PhD student Clara Bird, and myself, headed out in search of gray whales earlier than usual this year to document the potential effects of a National Science Foundation (NSF) funded seismic survey, which was conducted off the Oregon coast, on gray whales in the area. By the end of October, we had conducted 80 drone flights, collected 48 and 66 fecal and prey samples (respectively), and seen 36 individual whales during 201 sightings. Down south in Port Orford, the TOPAZ/JASPER project experienced a passing of the torch as I stepped down from the team lead position (which I held since 2018) and handed the project reins over to Allison. We co-led another fantastic field season this year. While whale sightings were much lower than in previous years (read some musings here), the project continued to be successful at making real impacts on young people’s lives as we once again engaged a local Pacific High School student (Damian Amerman-Smith) and two OSU undergraduates (Nadia Leal & Jasen White) in the field work. While our annual reach may be small in terms of numbers, the impact we have is huge, with many of the high school interns (including this year’s) deciding to go to college and/or to study biology directly as a result of our project.
TOPAZ/JASPER certainly is not the only project in our lab that engages students in ecological research. This year, we collectively oversaw and mentored 13 students. The OBSIDIAN (Observing Blue whale Spatial ecology to Investigate Distribution In Aotearoa New Zealand) project was assisted by three interns (Grace Hancock, Mateo Estrada Jorge, and Mattea Holt Colberg) overseen by Dawn and Leigh. Grace worked on maintaining the New Zealand blue whale photo-ID catalogue and won best student poster at our department’s annual student conference (RAFWE) for this work. Mattea, a 2020 TOPAZ/JASPER team member, switched study species and assisted Dawn in validating blue whale calls and songs. Mateo was a NSF Research Experience Undergraduate (REU) who conducted an analysis on blue whales and earthquakes. Clara also supervised a REU student with Leigh: Marc Donnelly, who created a habitat map for the GRANITE project. Rachel mentored Amanda Kent, an Undergraduate Research, Scholarship, & the Arts (URSA) Engage student, who helped her conduct a literature review about two Oregon krill species that are primary prey of whales. Over the summer, we had two student workers (Noah Goodwin-Rice & Julia Parker) join us in our efforts to better understand gray whale prey. Noah assisted us by sorting and identifying gray whale prey samples collected this summer and Julia wrapped up the microplastics analysis of gray whale prey and fecal samples. In the fall, both Clara and Allison supervised students (Kathryne Macallan & Jasen White, respectively) taking the Coastal & Estuarine Research Management class in our department who produced independent research projects during the term. Kathryne investigated the relationship between body length and blow intervals of gray whales during different behavioral states, while Jasen dove into the relationship between zooplankton abundance and environmental covariates.
The sharing of our research and expertise was not limited to mentoring students. Despite most conferences and seminars still occurring virtually this year due to the pandemic, the GEMM Lab presented numerous talks including at the State of the Coast (Rachel, Dawn, Leigh, & myself), International Biologging Symposium (Solène), HMSC Research Seminar (Ale & Solène, KC), and the Northwest Student Society of Marine Mammalogy chapter conference (Clara, Dawn, & myself), to name a few. Furthermore, Clara and I were guest lecturers once again for Dr. Renee Albertson’s marine mammal classes in our department, and Solène gained her first teaching experience by creating and leading a data visualization workshop (called “Pimp my figure!”) for RAFWE in May, which she reiterated at the University of New Caledonia in October.
Another huge accomplishment comes from the southern hemisphere as the hard work and time that Leigh and Dawn dedicated to OBSIDIAN and the results generated contributed to the denial of a seabed mining permit to extract iron sands in the South Taranaki Bight. This milestone has been years in the making, starting in 2013 when Leigh published her hypothesis that an unrecognized blue whale foraging ground existed in New Zealand. Since then, Leigh and Dawn have been building a tower of knowledge about these resident New Zealand blue whales block-by-block. They first confirmed Leigh’s hypothesis by presenting a bounty of evidence in support of this resident population, then assessed the skin condition of these whales, modeled the functional relationships between oceanography, krill and the distribution of blue whales, discovered temporal and spatial lags between wind, coastal upwelling, and blue whale occurrence, and most recently, developed dynamic models to forecast blue whale distribution three weeks into the future. We are extremely proud of the direct applications that the OBSIDIAN research outputs have had on the management and conservation of these New Zealand blue whales – hurrah to Leigh & Dawn!!
Other hurrahs this year include that Rachel passed her College of Earth, Ocean, & Atmospheric Sciences qualifying exam, now making her a PhD Candidate. Clara also reached a graduate milestone this year as she not only formed her PhD committee but also successfully defended her research review in the spring. Additionally, Clara became a certified drone pilot right before the start of the GRANITE field season and joined Todd and KC as pilots this summer. The lab and its members also received numerous grants and awards. There are too many to name for this blog, but we are very grateful for all of them! I do want to highlight two here: Dawn was awarded the Bob Moch memorial endowment award that recognizes service to the Hatfield Marine Science Center (HMSC) and broader Oregon coast community. I cannot think of anyone more deserving of this award than Dawn who truly does so much to serve and better the HMSC and Oregon coast communities! Clara was awarded a prestigious ARCS (Achievement Rewards for College Scientists) scholarship which provides awards to academically outstanding students to further their scientific knowledge.
We have once again been prolific writers, contributing 24 total peer-reviewed publications to 17 different scientific journals. If you are in the mood for some holiday reading, you will find the full list of publications at the end of this post.
And YOU, our awesome, supportive readers, have once again been supportive viewers, with a whopping 27,135 views of our blog this year!!! Thank you for joining us on our 2021 journey! We hope you have enjoyed the tales that we have told and the knowledge we have (hopefully) conveyed. We wish you all restful, happy, and most importantly, healthy holidays and hope you will join us again in 2022!
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get weekly updates and more! Just add your name into the subscribe box on the left panel.
Publications
Andréfouët, S., Derville, S., Buttin, J., Dirberg, G., Wabnitz, C.C.C., Garrigue, C., & Payri, C. E. 2021. Nation-wide hierarchical and spatially-explicit framework to characterize seagrass meadows in New Caledonia, and its potential application to the Indo-Pacific. Marine Pollution Bulletin 173:113036. https://doi.org/10.1016/j.marpolbul.2021.113036.
Barlow, D.R., & Torres, L.G. 2021. Planning ahead: Dynamic models forecast blue whale distribution with applications for spatial management. Journal of Applied Ecology. (Link)
Barlow, D.R., Klinck, H., Ponirakis, D., Garvey, C., & Torres, L.G. (2021). Temporal and spatial lags between wind, coastal upwelling, and blue whale occurrence. Scientific Reports 11(1):1-10. (Link)
Beal, M., … Torres, L.G., et al. 2021. Global political responsibility for the conservation of albatrosses and large petrels. Science Advances 7(10):eabd7225.
Bierlich, K.C., Schick, R.S., Hewitt, J., Dale, J., Goldbogen, J.A., Friedlaender, A.S., & Johnston D.J. 2021. A Bayesian approach for predicting photogrammetric uncertainty in morphometric measurements derived from UAS. Marine Ecology Progress Series. DOI: https://doi.org/10.3354/meps13814
Bierlich, K.C., Hewitt, J., Bird, C.N., Schick R.S., Friedlaender, A.S., Torres, L.G., Dale, J., Goldbogen, J.A., Read, A., Calambokidis J., & Johnston, D.W. 2021. Comparing uncertainty associated with 1-, 2-, and 3D aerial photogrammetry-based body condition measurements of baleen whales. Frontiers in Marine Science 8:749943. doi: 10.3389/fmars.2021.749943
Bonneville, C.D., Derville, S., Luksenburg, J.A., Oremus, M., Garrigue, C. 2021. Social structure, habitat use and injuries of Indo-Pacific Bottlenose Dolphins (Tursiops aduncus) reveal isolated, coastal, and threatened communities in the South Pacific. Frontiers in Marine Science 8:1–14. https://doi.org/10.3389/fmars.2021.606975.
D’Agostino, V.C., Fernandez, A.A.A., Degrati M., Krock, B., Hunt, K.E., Uhart, M.M., & Buck, C.L. 2021. Potential endocrine correlation with exposure to domoic acid in Southern Right Whale (Eubalaena australis) at the Península Valdés breeding ground. Oecologia 1-14.
Dillon, D., Fernandez, A.A.A., Hunt, K.E., & Buck, C.L. 2021. Investigation of keratinase digestion to improve steroid hormone extraction from diverse keratinous tissues. General and Comparative Endocrinology 309:113795.
Fernandez, A.A.A., Hunt, K.H., Sironi, M., Uhart, M., Rowntree, V., Giese, A.C., Maron, C.F., DiMartino, M., Dillon, D., & Buck, C.L. 2021. Retrospective analysis of the lifetime endocrine response of southern right whales calves to gull wounding and harassment: a baleen hormone approach. Integrative and Comparative Biology 61.
Fernandez, A.A.A., Hunt, K.E., Dillon, D., Uhart, M., Sironi, M., Rowntree, V., & Buck, C.L. 2021. Optimizing hormone extraction protocols for whale baleen: tackling questions of solvent: sample ratio and variation. General and Comparative Endocrinology 113828.
Garrigue, C., & Derville, S. 2021. Behavioral responses of humpback whales to biopsy sampling on a breeding ground : the influence of age-class , reproductive status , social context , and repeated sampling. Marine Mammal Science 1–16. https://doi.org/10.1111/mms.12848.
Gough, W.T., Smith, H.J., Savoca, M.S., Czapanskiy M.F., Fish, F.E., Potvin, J., Bierlich, K.C., Cade, D.E., Di Clemente, J., Kennedy, J., Segre, P., Stanworth, A., Weir, C., & Goldbogen, J.A. 2021. Scaling of oscillatory kinematics and Froude efficiency in baleen whales. Journal of Experimental Biology224(13):jeb237586. DOI: https://doi.org/10.1242/jeb.237586
Hildebrand, L., Bernard, K.S., & Torres, L.G. 2021. Go gray whales count calories? Comparing energetic values of gray whale prey across two different feeding grounds in the eastern North Pacific. Frontiers in Marine Science, https://doi.org/10.3389/fmars.2021.683634
Jones, D.C., Ceia, F.R., Murphy, E., Delord, K., Furness, R.W., Verdy, A., Mazloff, M., Phillips, R.A., Sagar, P.M., Sallée, J-B., Schreiber, B., Thompson, D.R., Torres, L.G., Underwood, P.J., Weimerskirch, H., & Xavier J.C. 2021. Untangling local and remote influences in two major petrel habitats in the oligotrophic Southern Ocean. Global Change Biology 27(22):5773-5785.
Kone, D.V., Tinker, M.T., & Torres, L.G. 2021. Informing sea otter reintroduction through habitat and human interaction assessment. Endangered Species Research 55:159-176.
Lemos, L.S., Olsen, A., Smith, A., Burnett, J.D., Chandler, T.E., Larson, S., Hunt, K.E., & Torres, L.G. 2021. Stressed and slim or relaxed and chubby? A simultaneous assessment of gray whale body condition and hormone variability. Marine Mammal Science.
Lemos, L.S., Haxel, J.H., Olsen, A., Burnett, J.D., Smith, A., Chandler, T.E., Nieukirk, S.L., Larson, S.E., Hunt, K.E., & Torres, L.G. 2021. Sounds of stress: assessment of relationships between ambient noise, vessel traffic, and gray whale stress hormone. Scientific Reports. DOI:10.21203/rs.3.rs-923450/v1
Maron, C.F., Lábaque, M.C., Beltramino L., DiMartino, M., Alzugaray, L., Ricciardi, M., Fernandez, A.A.A., Adler, F.R., Seger, J., Sironi, M., Rowntree, V.J., & Uhart, M.M. 2021. Patterns of blubber fat deposition and evaluation of body condition in growing southern right whale calves (Eubalaena australis). Marine Mammal Science. DOI: 10/1111/mms.12818.
Orben, R.A., Adams, J., Hester, M., Shaffer, S.A., Suryan, R.M., Deguchi, T., Ozaki, K., Sato, F., Young, L.C., Clatterbuck, C., Conners, M.G., Kroodsma, D.A., & Torres, L.G. 2021. Across borders: External factors and prior behavior influence North Pacific albatross associations with vessel traffic. Journal of Applied Ecology.
Savoca, M.S. Czapanskiy, M.F., Kahane-Rapport, S.R., Gough, W.T., Falhbusch, J.A., Bierlich, K.C., Segre, P.S., Di Clemente, J., Penry G.S., Wiley, D.N., Calambokids, J., Nowacek, D.P., Johnston, D.W., Pyenson, N.D., Friedlaender, A.S., Hazen, E.L., & Goldbogen, J.A. 2021. Baleen whale prey consumption based on high-resolution foraging measurements. Nature 599:85–90. https://doi.org/10.1038/s41586-021-03991-5
Stephenson, F., Hewitt, J.E., Torres, L.G., Mouton, T.L., Brough, T., Goetz, K.T., Lundquist, C.J., MacDiarmid, A.B., Ellis, J., & Constantine, R. 2021. Cetacean conservation planning in a global diversity hotspot: dealing with uncertainty and data deficiencies. Ecosphere 12(7):e03633.
Thompson, D.R., Goetz, K.T., Sagar, P.M., Torres, L.G., Kroeger, C.E., Sztukowski, LA., Orben, R.A., Hoskins, A.J., & Phillips, R.A. 2021. The year-round distribution and habitat preferences of Campbell albatross (Thalassarche impavida). Aquatic Conservation: Marine and Freshwater Ecosystems 31(10):2967-2978.
Since we find ourselves well into the cozy winter season, I thought it was an appropriate time to update you all on our project COZI (Coastal Oregon Zooplankton Investigation). COZI is a cross-college collaborative effort, led by GEMM PI Leigh Torres, that aims to better understand the quality of Oregon coast zooplankton prey and its impacts on gray whale foraging ecology and health. Leigh is joined by three other early-career female scientists, Dr. Sarah Henkel, Dr. Kim Bernard, and Dr. Susanne Brander, that each contribute a different area of expertise to the project. The quartet recently graced the cover of the Oregon Stater in an article all about COZI written by Nancy Steinberg (which I highly recommend reading!). To date, the COZI team (which includes myself as well as many other students) has found that the caloric content of the six predominant zooplankton species in Oregon coastal waters differs significantly, with Dungeness crab megalopae coming out on top as a caloric goldmine (Hildebrand et al. 2021). We found that these Oregon prey are calorically competitive with the predominant benthic amphipod that gray whales feed on in the Arctic, which has interesting implications for foraging ground selection and use of gray whales in the eastern North Pacific (read about it in detail in my blog about the publication). Now that we know that Oregon zooplankton quality differs in terms of calories, we are curious to determine whether these species are impacted by microplastics in the environment, to what extent, and how gray whales may be affected.
What is in those zooplankton? A microscopic view of several mysid shrimp collected in Oregon coastal waters. Source: L. Hildebrand.
To answer these questions, we are analyzing both zooplankton and gray whale fecal samples for microplastics to see what kind, and how many, microplastics we find, and whether microplastics biomagnify up the food chain. The lab analysis has just been completed and we are working on interpreting the results. We can’t let the cat out of the bag yet, but a little sneak-peek of what we have found is that there are different levels of microplastic loads by zooplankton species, which also end up in the whale poop. So, until we finalize those results for sharing, I am going to review the field of microplastics research, with a particular focus on cetaceans. Avid readers of our blog may recall that I wrote a blog about marine plastics at the start of 2019. In that blog, I mentioned that a GoogleScholar search of “microplastics marine” generated 7,650 results. To get an idea of how microplastics research in the marine environment has progressed since I wrote my 2019 blog, I conducted the same GoogleScholar search for this blog but I limited the results to studies published between 2019-2021. GoogleScholar presented me with a whopping 18,000+ results, which shows the rapidity at which the field of marine microplastics research has grown in the last couple of years. The studies span all kinds of topics from distribution & occurrence, to chemical behaviors & interactions with other toxins, to sources & sinks (to name a few!). The results encompass both laboratory and field studies investigating samples from all five oceans of the world. Unfortunately, the title of my blog from two years ago still rings very true: plastics truly are ubiquitous in the marine environment.
In my last blog, I listed three cetacean species that had been found to contain microplastics: a True’s beaked whale (Lusher et al. 2015), a humpback whale (Besseling et al.2015) and an Indo-Pacific humpback dolphin (Zhu et al.2018). Reflective of the marine microplastics field in general, this list has also grown considerably in the last two years. Since 2019, microplastics have been detected in harbor porpoises (Philipp et al. 2021), common dolphins (Nelms et al. 2019), striped dolphins (Novillo et al. 2020), bottlenose dolphins (Battaglia et al. 2020), Atlantic white-sided dolphins (Nelms et al. 2019), beluga whales (Moore et al. 2020), and Bryde’s & sei whales (Zantis et al. 2021). At this point, I would posit that the main reason this list is not longer is due to the time it takes to collect and analyze samples for microplastics, rather than microplastics being absent in other cetacean species. During my research for this blog, I noticed that the studies on microplastics in cetaceans are starting to shift from focusing on simply determining microplastic occurrence to attempting to estimate levels of exposure and/or ingestion, determine the main source (from water vs. from prey), and long-term consequences.
Graphical abstract taken from Zantis et al. (2021) representing the pathway of microplastics exposure of large marine filter-feeders. Source: Zantis et al. (2021).
A study published this year examined fecal samples of Bryde’s and sei whales in coastal waters in New Zealand and detected 32 ± 24 microplastics per 6 g of feces (Zantis et al. 2021). By extrapolating these values to the proportions of prey species in the whales’ diet, the authors estimate that these whales consume over 24,000 pieces of microplastics per mouthful of prey, or more than 3 million microplastics per day. Another study (Shetty 2021) in the same geographic region investigated the levels of microplastics in coastal surface waters, which allowed the authors to estimate whether the source of the microplastics that the Bryde’s and sei whales ingest come from the water or the prey. They found that the estimated level of microplastics that the whales consume daily from their prey is four orders of magnitude higher than the microplastic levels in the coastal waters. This finding strongly suggests that the predominant mode of exposure of large filter feeders, such as baleen whales, for microplastic pollution comes from their prey through biomagnification (not just from the ambient sea water).
The GEMM Lab collecting a gray whale fecal sample along the Oregon coast captured from a drone. Source: GEMM Lab.
COZI aims to conduct similar analyses as these studies described above to understand the exposure of coastal Oregon zooplankton to microplastics and how this may be affecting gray whales. Stay tuned for those results!
I am aware that I have painted a very bleak (but true) picture of microplastic pollution in our oceans in this blog but there are things you can do to help reduce microdebris in the environment!
A major source of pollution in the ocean comes from microfibers through our laundry. You can help stop this pathway by simply using a Cora Ball or installing a filter (such as this one) in your washing machine that captures microfleece & polyester fibers.
Minimize your use of single-use plastics. There are so many ways to do so including reuseable water bottles, travel mugs for coffee or tea, fabric totes as shopping bags, carry a set of utensils for takeout food, beeswax wraps instead of plastic wrap or sandwich bags.
Use public transport when possible as another huge source of microplastics comes from tire treads! This solution also helps reduce your carbon footprint.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get a weekly message when we post a new blog. Just add your name into the subscribe box below.
References
Battaglia, F.M., Beckingham, B.A., & McFee, W.E. 2020. First report from North America of microplastics in the gastrointestinal tract of stranded bottlenose dolphins (Tursiops truncatus). Marine Pollution Bulletin 160:111677.
Besseling, E., et al. 2015. Microplastic in a macro filter feeder: humpback whale Megaptera novaeangliae. Marine Pollution Bulletin 95: 248-252.
Hildebrand, L., Bernard, K.S., & Torres, L.G. 2021. Do gray whales count calories? Comparing energetic values of gray whale prey across two different feeding grounds in the eastern North Pacific. Frontiers in Marine Science. https://doi.org/10.3389/fmars.2021.683634
Lusher, A.L., et al. 2015. Microplastic and macroplastic ingestion by a deep diving, oceanic cetacean: the True’s beaked whales Mesoplodon mirus. Environmental Pollution 199: 185-191.
Moore, R.C., et al. 2020. Microplastics in beluga whales (Delphinapterus leucas) from the eastern Beaufort Sea. Marine Pollution Bulletin 150:110723.
Nelms, S.E., et al. 2019. Microplastics in marine mammals stranded around the British coast: ubiquitous bus transitory? Scientific Reports 9:1075.
Novillo, O., Raga, J. A., & Tomás, J. 2020. Evaluating the presence of microplastics in striped dolphins (Stenella coeruleoalba) stranded in the western Mediterranean Sea. Marine Pollution Bulletin 160:111557.
Philipp, C., et al. 2021. First evidence of retrospective findings of microplastics in harbor porpoises (Phocoena phocoena) from German waters. Frontiers in Marine Science. https://doi.org/10.3389/fmars.2021.682532
Shetty, D. 2021. Incidence of microplastics in coastal inshore fish species and surface waters in the Hauraki Gulf, New Zealand. Master’s thesis, University of Auckland, New Zealand.
Zantis, L.J., et al. 2021. Assessing microplastic exposure of large marine filter-feeders. Science of The Total Environment 151815.
Zhu, J., et al. 2018. Cetaceans and microplastics: First report of microplastic ingestion by a coastal delphinid, Sousa chinensis. Science of the Total Environment 659: 649-654.
Over six field seasons the GEMM lab team has conducted nearly 500 drone flights over gray whales, equaling over 100 hours of footage. These hours of footage are the central dataset for my PhD dissertation, so it’s up to me to process them all. This process can be challenging, tedious, and daunting, but it is also quite fun and a privilege to be the one person who gets to watch all the footage. It’s fascinating to get to know the whales and their behaviors and pick up on patterns. It motivates me to get through this video processing step and start doing the data analysis. Recently, it’s been especially fun to notice patterns that I’ve seen mentioned in the literature. One example is adult social behavior.
There are two categories of social behavior that I’m interested in studying: maternal behavior, defined as interactions between a mom and its calf, and general social behaviors, defined as social interactions between non-mom/calf pairs. In this blog I’ll focus on general social behaviors, but if you’re interested in maternal behavior check out this blog. General social behavior, which I’ll refer to as social behavior moving forward, includes tactile interactions and promiscuous behaviors (Torres et al. 2018; Clip 1). While gray whales in the PCFG range are primarily foraging, researchers have observed increases in social behavior towards the end of the foraging season (Stelle et al., 2008; Torres et al., 2018). We think that this indicates that the whales are starting to focus less on feeding and more on breeding. This tradeoff of foraging vs. socializing time is interesting because it comes at an energetic cost.
Clip 1. Example of social interaction between a male and female gray whale off the coast of Oregon, USA. Collected under NOAA/NMFS permit #21678
Broadly, animals need to balance the energetic demands of survival with those of reproduction. They need to reproduce to pass on their genes, but reproduction is energetically demanding, and animals also need to survive and grow to be able to reproduce. The decision to reproduce is costly because reproduction requires energetic investment and time investment since animals do not forage (gaining energy) when they are socializing. Consequently, only animals with sufficient energy reserves (i.e., body condition) to invest in reproduction actually engage in reproduction. Given these costs associated with reproduction, we expect to see a relationship between social behavior and body condition (Green, 2001) with mainly animals in good body condition engaging in social behavior because these animals have sufficient reserves to sustain the cost. Furthermore, since body condition is an indicator of foraging success and prey availability, environmental conditions can also affect social behavior and reproduction through this pathway.
Rahman et al. (2014) used a lab experiment to study the relationship between nutritional stress and male guppy courtship behavior (Figure 1). In their experiment they tested for the effects of both decreased diet quantity and quality on the frequency of male courtship behaviors. Rahman et al (2014) found that individuals in the low-quantity group were significantly smaller than those in the high-quality group and that diet quantity had a significant effect on the frequency of courtship behaviors. Males fed a low-quantity diet performed fewer courtship behaviors. Interestingly, there was no significant effect of diet quality on courtships behavior, although there was some evidence of an interaction effect, which suggests that within the low-quantity group, males fed with high-quality food performed more courtship behaviors that those fed with low-quality food. This study is interesting because it shows how foraging success (diet quantity and quality) can affect courting behavior.
Figure 1. A guppy (Rahman et al., 2013)
However, guppies are not the ideal species for comparison to gray whales because gray whales and guppies have quite different life history traits. A more fitting comparison would be with an example species with more in common with gray whales, such as viviparous capital breeders. Viviparous animals develop the embryo inside the body and give live birth. Capital breeders forage to build energy reserves and then rely on those energy reserves during reproduction. Surprisingly, I found asp vipers to be a good example species for comparison to gray whales.
Asp vipers (Figure 2) are viviparous snakes who are considered capital breeders because they forage prior to hibernation, and then begin reproduction immediately following hibernation without additional foraging. Naulleau & Bonnet (1996) conducted a field study on female asp vipers to determine if there was a difference in body condition at the start of the breeding season between females who reproduced or not during that season. To do this they marked individuals and measured their body condition at the start of the breeding season and then recaptured those individuals at the end of the breeding season and recorded whether the individual had reproduced. Interestingly, they found that there was a strongly significant difference in body condition between females that did and did not reproduce. In fact, they discovered that no female below a certain body condition value reproduced, meaning that they found a body condition threshold for reproduction.
Figure 2. An asp viper
Additionally, a study on water pythons found that their body condition threshold for reproduction shifted over time in response to prey availability (Madsen & Shine, 1999). These authors found that females lowered their threshold after several consecutive years of poor prey availability. These studies are really exciting to me because they address questions that the GRANITE project team is interested in tackling.
Understanding the relationship between body condition and reproduction in gray whales is an important puzzle piece for our work. The aim of the GRANITE project is to understand how the effects of stressors on individual whales scales up to population level impacts (read Lisa’s blog to learn more). Reproduction rates play a big role in population dynamics, so it is important to understand what factors affect reproduction. Since we’re studying these whales on their foraging grounds, assessing body condition provides an important link between foraging behavior and reproduction.
For example, if an individual’s response to a stressor is to forage less, that may lead to poorer body condition, meaning that they may be less likely to reproduce. While reduced reproduction in one individual may not have a big effect on the population, the same response from multiple individuals could impact the population’s dynamics (i.e., increasing or decreasing abundance). Understanding these different relationships between behavior, body condition, and reproduction rates is a big undertaking, but it’s exciting to be a member of the GRANITE team as this strong group of scientists works to bring together different data streams to work on this big picture question. We’re all deep into data processing right now so stay tuned over the next few years to learn more about gray whale social behavior and to find out if fat whales are more social than skinny whales.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get weekly updates and more! Just add your name into the subscribe box on the left panel.
References
Green, A. J. (2001). Mass/Length Residuals: Measures of Body Condition or Generators of Spurious Results? Ecology, 82(5), 1473–1483. https://doi.org/10.1890/0012-9658(2001)082[1473:MLRMOB]2.0.CO;2
Madsen, T., & Shine, R. (1999). The adjustment of reproductive threshold to prey abundance in a capital breeder. Journal of Animal Ecology, 68(3), 571–580. https://doi.org/10.1046/j.1365-2656.1999.00306.x
Naulleau, G., & Bonnet, X. (1996). Body Condition Threshold for Breeding in a Viviparous Snake. Oecologia, 107(3), 301–306.
Rahman, M. M., Kelley, J. L., & Evans, J. P. (2013). Condition-dependent expression of pre- and postcopulatory sexual traits in guppies. Ecology and Evolution, 3(7), 2197–2213. https://doi.org/10.1002/ece3.632
Rahman, M. M., Turchini, G. M., Gasparini, C., Norambuena, F., & Evans, J. P. (2014). The Expression of Pre- and Postcopulatory Sexually Selected Traits Reflects Levels of Dietary Stress in Guppies. PLOS ONE, 9(8), e105856. https://doi.org/10.1371/journal.pone.0105856
Stelle, L. L., Megill, W. M., & Kinzel, M. R. (2008). Activity budget and diving behavior of gray whales (Eschrichtius robustus) in feeding grounds off coastal British Columbia. Marine Mammal Science, 24(3), 462–478. https://doi.org/10.1111/j.1748-7692.2008.00205.x
Torres, L. G., Nieukirk, S. L., Lemos, L., & Chandler, T. E. (2018). Drone up! Quantifying whale behavior from a new perspective improves observational capacity. Frontiers in Marine Science, 5(SEP). https://doi.org/10.3389/fmars.2018.00319
Dr. Alejandro Fernández Ajó, OSU Department of Fisheries, Wildlife, & Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
Rises in ocean temperatures can lead to multiple alterations in marine ecosystems, including the increase and the frequency of Harmful Algal Blooms (HABs). HABs are characterized by the rapid growth of toxin-producing species of algae that can be harmful to people, animals, and the local ecology, even causing death in severe cases. Species of marine diatom within the genus Pseudo-nitzschia and Nitzschia can form HABs when they produce domoic acid (DA), a potent neurotoxin responsible for amnesic shellfish poisoning (D’Agostino et al., 2018, 2017).
Figure 1. Southern right whale (E. australis) mother and calf swimming at the gulfs of Peninsula Valdes, Argentina, during a phytoplankton bloom. Photo: Mariano Sironi / Instituto de Conservacion de Ballenas de Argentina.
During HABs, DA is transferred to higher organisms through the pelagic food web and is accumulated by intermediate vectors, such as copepods, euphausiids (i.e., krill), shellfish, and fish. As this neurotoxin affects top predators, DA poisoning poses a risk to the safety and health of humans and wildlife. This neurotoxin has caused mortality in many marine mammal species, including both pinnipeds and cetaceans (Gulland 1999; Lefebvre et al. 1999; Fire et al. 2010, 2021; Broadwater et al. 2018). In addition, the exposure to DA constitutes a stressor that may affect glucocorticoids (hormones involved in the stress response) concentrations.
The glucocorticoids (GCs; cortisol and corticosterone) are adrenal steroid hormones that maintain the essential functions of metabolism and energy balance in mammals. GCs can increase sharply in response to environmental stressors to elicit physiological and behavioral adaptations by individuals to support survival (Sapolsky et al. 2000; Bornier et al. 2009). However, with the chronic exposure to a stressor, this relationship can reverse, with GCs sometimes declining below its baseline levels (Dickens and Romero, 2013; Fernández Ajó et al., 2018). Moreover, DA can interfere with the stress response in mammals, and cause alterations in their physiological response. DA is an excitatory amino acid analog of glutamate (Pulido 2008), a well-known brain neurotransmitter that play an important role in the activation of the adrenal axis (which in turn regulate the production and secretion of the GCs) and regulate many of the pituitary hormones involved in the stress response (Brann and Mahesh 1994; Johnson et al. 2001). Hence, monitoring GC levels in marine mammals can be a potential useful metric for assessing the physiological impacts of exposure to DA.
Glucocorticoids are traditionally measured in plasma, but given that plasma sampling from free-ranging large whales is currently impossible, alternative sample types such as fecal samples, among others, can be utilized to quantify GCs in large whales (Ajó et al., 2021; Burgess et al., 2018, 2016; Fernández Ajó et al., 2020, 2018; Hunt et al., 2019, 2014, 2006; Rolland et al., 2017, 2005)(Figure 2). The analyses of fecal glucocorticoid metabolites (fGCm) is particularly useful for endocrine assessments of free-swimming whales, with several studies showing that fGCm correlate in meaningful ways with presumed stressors. For example, high levels of fGCm in North Atlantic right whales (NARW, Eubalaena glacialis) and in gray whales (Eschrichtius robustus) correlate with poor body condition (Hunt et al., 2006; Lemos et al., 2021), and fGCm increases were associated with whale entanglements and ship strikes (i.e., Lemos et al., 2020; Rolland et al., 2017).
Figure 2. Alternative samples types can be used to study hormones in large whales. 1-2-3 are sample types that can be obtained from free-living whales and provide a more instantaneous and acute measurement of the whales´ physiology. 4-5 can be obtained at necropsy when the whale is found dead at the beach and provide an integrated measure of the whale physiology that can expand through years or even the lifespan of an individual.
In Península Valdés, Argentina, southern right whales (SRW, E. australis) gather in large numbers to mate and nurse their calves during the austral winter months (Bastida and Rodríguez, 2009). SRWs are capital breeders, largely fasting during the breeding season and instead relying on stored blubber fuel reserves. However, they can occasionally feed on calanoid copepods (D’Agostino et al., 2018, 2016), particularly during the phytoplankton blooms that are dominated by diatoms of the genus Pseudo-nitzschia (Sastre et al. 2007; D’Agostino et al. 2015, 2018). Therefore, feeding SRWs in Península Valdés temporally overlap with these Pseudo-nitzschia blooms (D’Agostino et al. 2018, 2015) and represents a test case for assessing the relationship of DA exposure with GC levels (Figure 3).
Figure 3. Southern right whale (E. australis) skim feeding at the Peninsula Valdes breeding ground. Photo: Lucas Beltranino.
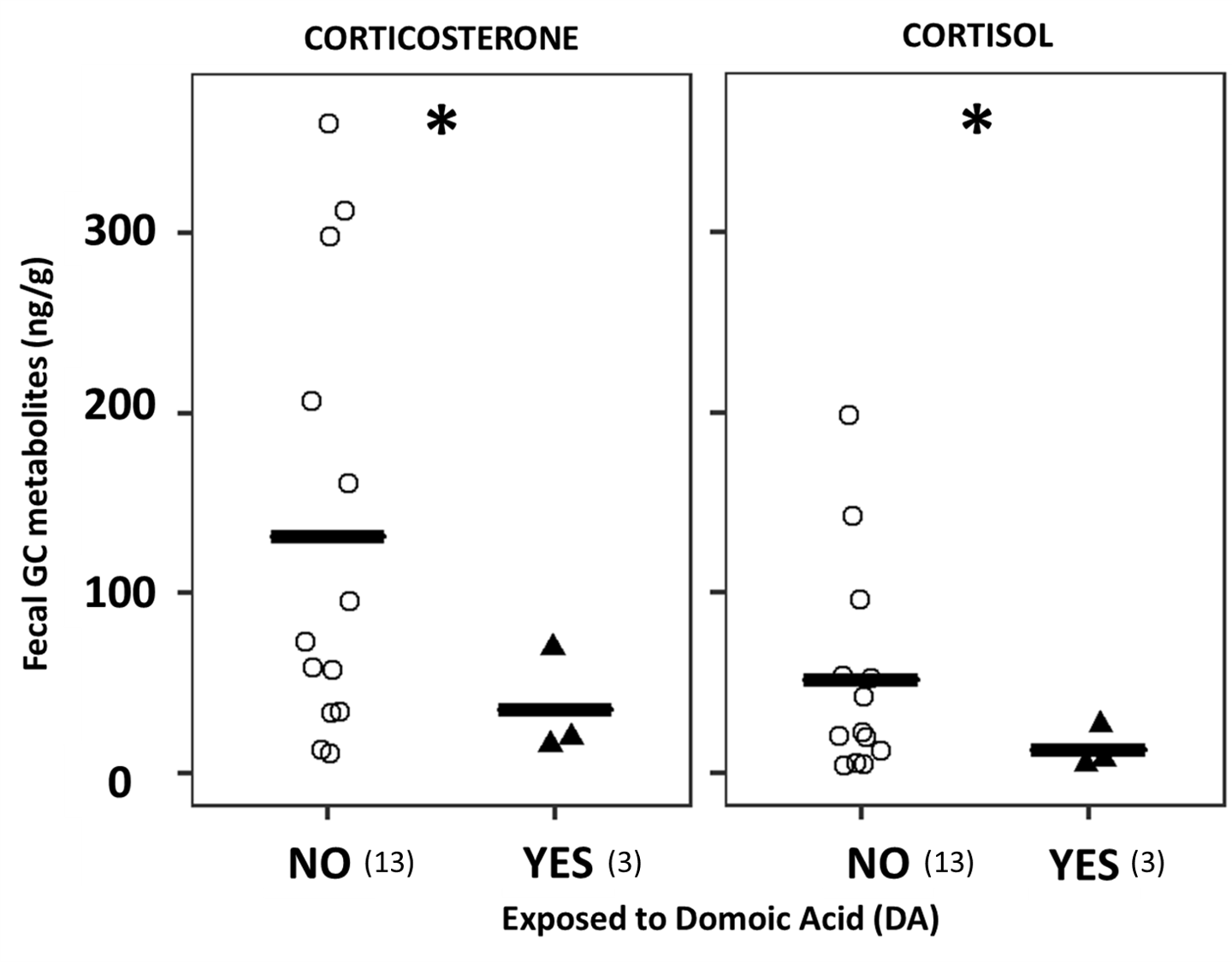
In our recent scientific publication (D’Agostino et al. 2021), we investigate SRW exposure to DA at their breeding ground in Peninsula Valdes and assessed its effects on fecal glucocorticoid concentrations. Although the sample size of this study is unavoidably small due to the difficulties of obtaining fecal samples from whales at their calving grounds where defecation is infrequent, we observed significantly lower fGCm in samples from whales exposed to DA (Figure 4). Our results agree with findings from a previous study in California sea lions (Zalophus californianus) exposed to DA, where these authors found a significant association of DA exposure with reduced serum cortisol (Gulland et al., 2009), which can be tentatively attributed to abnormal function of the adrenal axis due to the exposure.
Figure 4. Fecal glucocorticoid metabolite levels in southern right whales exposed (YES, solid triangles) and not-exposed (NO, open circles) to DA. Left panel: immunoreactive fecal corticosterone metabolites. Right panel: immunoreactive fecal cortisol metabolites. Hormone concentrations are expressed in ng of immunoreactive hormone per gram of dry fecal sample. Significant differences between groups are denoted with an asterisk (P<0.05). The black solid line indicates the mean for each group, and in parenthesis is the sample size for each group. Adapted from D’Agostino et al. 2021.
If ingestion of toxins produced by phytoplankton can result in long-term suppression of baseline GCs, whales and marine mammals in general, could suffer reduced ability to cope with additional stressors. The adrenal function is essential to maintain circulating blood glucose and other aspects of metabolism within normal bounds. Additionally, the ability to elevate GCs facilitates energy mobilization to physiologically cope with a stressful event and to initiate appropriate behavioral responses (i.e., flee from predators, heal wounds). Various toxicants have been shown to reduce adrenal function across taxa (Romero and Wingfield, 2016) and could have negative consequences on the ability of cetaceans to respond and adapt to ongoing environmental and anthropogenic changes. Compounding this problem, whales are exposed to an increasing number of stressors from multiple sources and with cumulative effects and they need to be able to physiologically respond to continue to reproduce and survive.
To our knowledge, this study provides the first quantification of fGCm levels in whales exposed to DA; and we hope this effort starts a growing dataset to which other researchers can add. Sampling and analysis of non-traditional matrices, such as feces, blubber, baleen and others, would likely increase sample sizes and thus our understanding of the interrelationships among DA exposure and age, sex, and reproductive status of cetaceans. Given that chronic exposure to DA could alter the capacity of animals to respond to stress, and indications that HABs are becoming more frequent and intense world-wide (Van Dolah 2000; Masó et al. 2006; Erdner et al. 2008), we believe that research evaluating the health status of marine mammal populations should include the assessment of stress physiology relative to natural and anthropogenic stressors including exposure to toxicants.
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get weekly updates and more! Just add your name into the subscribe box on the left panel.
References
Bastida R, Rodríguez D (2009) Ballena franca austral [Southern right whale]. In: Mazzini V (ed) Mamíferos marinos de Patagonia y Antártida [Marine mammals of Patagonia and Antarctica]. Zagier & Urruty Publications, Buenos Aires, pp 72–84.
Bonier F, Moore IT, Martin PR, Robertson RJ (2009) The relationship between fitness and baseline glucocorticoids in a passerine bird. Gen Comp Endocrinol 163:208–213. https:// doi. org/ 10. 1016/j.ygcen. 2008. 12. 013.
Brann DW, Mahesh VB (1994) Excitatory amino acids: function and significance in reproduction and neuroendocrine regulation. Front Neuroendocrinol 15:3–49. https:// doi. org/ 10. 1006/ frne. 1994. 1002.
Broadwater MH, Van Dolah FM, Fire SE (2018) Vulnerabilities of marine mammals to harmful algal blooms. Harmful Algal Blooms 2:191–222.
Burgess, E.A., Hunt, K.E., Kraus, S.D., Rolland, R.M., 2016. Get the most out of blow hormones: Validation of sampling materials, field storage and extraction techniques for whale respiratory vapour samples. Conserv. Physiol. 4, cow024. https://doi.org/10.1093/conphys/cow024
Burgess, E.A., Hunt, K.E., Kraus, S.D., Rolland, R.M., 2018. Quantifying hormones in exhaled breath for physiological assessment of large whales at sea. Sci. Rep. 8, 10031. https://doi.org/10.1038/s41598-018-28200-8
D’Agostino VC, Hoffmeyer MS, Almandoz GO, Sastre V, Degrati M., 2015. Potentially toxic Pseudo-nitzschia species in plankton and fecal samples of Eubalaena australis from Península Valdés calv-ing ground, Argentina. J Sea Res 106:39–43. https:// doi. org/ 10.1016/j. seares.
D’Agostino, V.C., Degrati, M., Santinelli, N., Sastre, V., Dans, S.L., Hoffmeyer, M.S., 2018. The seasonal dynamics of plankton communities relative to the foraging of the southern right whale (Eubalaena australis) in northern Patagonian gulfs, Península Valdés, Argentina. Cont. Shelf Res. 164, 45–57. https://doi.org/10.1016/j.csr.2018.06.003
D’Agostino, V.C., Degrati, M., Sastre, V., Santinelli, N., Krock, B., Krohn, T., Dans, S.L., Hoffmeyer, M.S., 2017. Domoic acid in a marine pelagic food web: Exposure of southern right whales Eubalaena australis to domoic acid on the Península Valdés calving ground, Argentina. Harmful Algae 68, 248–257. https://doi.org/10.1016/j.hal.2017.09.001
D’Agostino, V.C., Hoffmeyer, M.S., Degrati, M., 2016. Faecal analysis of southern right whales (Eubalaena australis) in Península Valdés calving ground, Argentina: Calanus australis, a key prey species. J. Mar. Biol. Assoc. United Kingdom 96, 859–868. https://doi.org/10.1017/S0025315415001897
Dickens, M.J., Romero, L.M., 2013. A consensus endocrine profile for chronically stressed wild animals does not exist. Gen. Comp. Endocrinol. 191, 177–189. https://doi.org/10.1016/j.ygcen.2013.06.014
Erdner DL, Dyble J, Parsons ML, Stevens RC, Hubbard KA, Wrabel ML, Moore SK, Lefebvre KA, Anderson DM, Bienfang P, Bidi-gare RR, Parker MS, Moeller P, Brand LE, Trainer VL (2008) Centers for Oceans and Human Health: a unified approach to the challenge of harmful algal blooms. Environ Health 7:S2. https://doi. org/ 10. 1186/ 1476- 069X-7- S2- S2.
Fernández Ajó, A., Hunt, K.E., Dillon, D., Uhart, M., Sironi, M., Rowntree, V., Buck, C.L., 2021. Optimizing hormone extraction protocols for whale baleen: tackling questions of solvent:sample ratio and variation. Gen. Comp. Endocrinol. 113828. https://doi.org/10.1016/j.ygcen.2021.113828
Fernández Ajó, A.A., Hunt, K.E., Giese, A.C., Sironi, M., Uhart, M., Rowntree, V.J., Marón, C.F., Dillon, D., DiMartino, M., Buck, C.L., 2020. Retrospective analysis of the lifetime endocrine response of southern right whale calves to gull wounding and harassment: A baleen hormone approach. Gen. Comp. Endocrinol. 296, 113536. https://doi.org/10.1016/j.ygcen.2020.113536
Fernández Ajó, A.A., Hunt, K.E., Uhart, M., Rowntree, V., Sironi, M., Marón, C.F., Di Martino, M., Buck, C.L., 2018. Lifetime glucocorticoid profiles in baleen of right whale calves: potential relationships to chronic stress of repeated wounding by Kelp Gulls. Conserv. Physiol. 6, 1–12. https://doi.org/10.1093/conphys/coy045
Fire SE, Bogomolni A, DiGiovanni RA Jr, Early G, Leighfield TA, Matassa K, Miller GA, Moore KM, Moore M, Niemeyer M, Pugliares K (2021) An assessment of temporal, spatial and taxonomic trends in harmful algal toxin exposure in stranded marine mammals from the US New England coast. PLoS ONE 16(1):e0243570. https:// doi. org/ 10. 1371/ journ al. pone. 02435 70
Fire SE, Wang Z, Berman M, Langlois GW, Morton SL, Sekula-Wood E, Benitez-Nelson CR (2010) Trophic transfer of the harmful algal toxin domoic acid as a cause of death in a minke whale (Balaenoptera acutorostrata) stranding in southern California. Aquat Mamm 36(4):342–350. https:// doi. org/ 10. 1578/ AM. 36.4.2010. 342.
Gulland F.M., 1999. Domoic acid toxicity in California sea lions stranded along the central California Coast, May-October 1998. NOAA Tech. Memo. NMFS-OPR-8. USA National Marine Fisheries Service, US Department of Commerce.
Gulland FMD, Hall AJ, Greig DJ, Fram ER, Colegrove KM, Booth RKN, Wasser SK, S.-M.C., 2009. Gulland, Hall – 2012 – Evaluation of circulating eosinophil count and adrenal gland function in California sea lions naturally exposed t. J. Am. Vet. Med. Assoc. 241, 943–949.
Gulland, F., Pérez-Cortés, H., Urbán, J.R., Rojas-Bracho, L., Ylitalo, G., Weir, J., Norman, S., Muto, M., Rugh, D., Kreuder, C., Rowles, T., 2005. Eastern North Pacific gray whale (Eschrichtius robustus) unusual mortality event, 1999-2000. U.S. Dep. Commer. NOAA Tech. Memo. NMFS-AFSC-150. 33 pp.
Hunt, K.E., Robbins, J., Buck, C.L., Bérubé, M., Rolland, R.M., 2019. Evaluation of fecal hormones for noninvasive research on reproduction and stress in humpback whales (Megaptera novaeangliae). Gen. Comp. Endocrinol. 280, 24–34. https://doi.org/10.1016/j.ygcen.2019.04.004
Hunt, K.E., Rolland, R.M., Kraus, S.D., Wasser, S.K., 2006. Analysis of fecal glucocorticoids in the North Atlantic right whale (Eubalaena glacialis). Gen. Comp. Endocrinol. 148, 260–272. https://doi.org/10.1016/j.ygcen.2006.03.012
Hunt, K.E., Stimmelmayr, R., George, C., Hanns, C., Suydam, R., Brower, H., Rolland, R.M., 2014. Baleen hormones: a novel tool for retrospective assessment of stress and reproduction in bowhead whales (Balaena mysticetus). Conserv. Physiol. 2, cou030–cou030. https://doi.org/10.1093/conphys/cou030
Johnson MP, Kelly G, Chamberlain M (2001) Changes in rat serum corticosterone after treatment with metabotropic glutamate receptor agonists or antagonists. J Neuroendocrinol 13:670–677. https:// doi. org/ 10. 1046/j. 1365- 2826. 2001. 00678.x.
Lefebvre KA, Powell CL, Busman M, Doucette GJ, Moeller PDR, Sliver JB, Miller PE, Hughes MP, Singaram S, Silver MW, Tjeer-dema RS (1999) Detection of domoic acid in northern anchovies and California sea lions associated with an unusual mortality event. Nat Toxins 7(3):85–92. https:// doi. org/ 10. 1002/ (SICI) 1522-7189(199905/ 06)7: 3% 3C85:: AID- NT39% 3E3.0. CO;2-Q.
Lefebvre, K.A., Kendrick, P.S., Ladiges, W., Hiolski, E.M., Ferriss, B.E., Smith, D.R., Marcinek, D.J., 2017. Chronic low-level exposure to the common seafood toxin domoic acid causes cognitive deficits in mice. Harmful Algae 64, 20–29. https://doi.org/10.1016/j.hal.2017.03.003
Lemos, L.S., Olsen, A., Smith, A., Burnett, J.D., Chandler, T.E., Larson, S., Hunt, K.E., Torres, L.G., 2021. Stressed and slim or relaxed and chubby? A simultaneous assessment of gray whale body condition and hormone variability. Mar. Mammal Sci. 1–11. https://doi.org/10.1111/mms.12877
Lemos, L.S., Olsen, A., Smith, A., Chandler, T.E., Larson, S., Hunt, K., Torres, L.G., 2020. Assessment of fecal steroid and thyroid hormone metabolites in eastern North Pacific gray whales. Conserv. Physiol. 8. https://doi.org/10.1093/conphys/coaa110
Masó M, Garcés E., 2006. Harmful microalgae blooms (HAB); prob-lematic and conditions that induce them. Mar Pollut Bull 53:620–630. https:// doi. org/ 10. 1016/j. marpo lbul. 2006. 08. 006.
Pulido O.M., 2008. Domoic acid toxicologic pathology: a review. Mar Drugs 6(2):180–219. https:// doi. org/ 10. 3390/ md602 0180.
Rolland, R., McLellan, W., Moore, M., Harms, C., Burgess, E., Hunt, K., 2017. Fecal glucocorticoids and anthropogenic injury and mortality in North Atlantic right whales Eubalaena glacialis. Endanger. Species Res. 34, 417–429. https://doi.org/10.3354/esr00866
Rolland, R.M., Hunt, K.E., Kraus, S.D., Wasser, S.K., 2005. Assessing reproductive status of right whales (Eubalaena glacialis) using fecal hormone metabolites. Gen. Comp. Endocrinol. 142, 308–317. https://doi.org/10.1016/j.ygcen.2005.02.002
Romero, M.L., Wingfield, J.C., 2016. Oxford series in behavioral neuroendocrinology. Tempests, poxes, predators, and people: stress in wild animals and how they cope 1–2.
Sastre V, Santinelli N, Marino G, Solís M, Pujato L, Ferrario M., 2007. First detection of domoic acid produced by Pseudo-nitzschia spe-cies, Chubut coastal waters, Patagonia, Argentina. Harmful Algae News 34:12–14.
Van Dolah FM., 2000. Marine algal toxins: origins, health effects, and their increased occurrence. Environ Health Perspect 108:133–141. https:// doi. org/ 10. 1289/ ehp. 00108 s1133
Allison Dawn, GEMM Lab Master’s student, OSU Department of Fisheries, Wildlife and Conservation Sciences, Geospatial Ecology of Marine Megafauna Lab
As the first term of my master’s program comes to an end and we head toward winter break, I am excited by the course material that has already helped direct my research and development as a scientist. There have been new, challenging topics to tackle, and each assignment has fostered deeper thinking into the formation of my thesis. While I learned new methods and analysis approaches this term, a single phrase pervades throughout my studies of ecology – “it depends!”. Ecologists work to uncover patterns driven by natural processes, and this single phrase seems to answer many questions about whether the pattern always exists. A reasonable follow up to that frequently used phrase is, “depends on what?” or “when or where would this pattern change?” In the context of foraging ecology, predator-prey patterns are frequently driven by environmental processes that depend on the scale you choose for your study.
What do we mean by scale? Simply stated, scale is a graduation from one level of measurement to another. You can imagine a ruler, for example. You can measure how tall you are in inches with a ruler or in yards with a yard stick. When we think about scale in ecology, the “ruler” can have traditional units of space (meters, kilometers, etc.), units of time (minutes, days, hours, months, years, etc.), or sometimes both!
The ocean is dynamic and heterogeneous, which simply means there is a lot going on at once. Oceanographic processes influence predator-prey interactions but due to the inherent variability in the system, it is important to explore which factors drive processes that influence patterns at different spatial and temporal scales.
In marine ecology, the “explanatory power” of a factors’ influence on a given process depends on which scale you choose to build your research upon. Ocean ecosystems are hierarchical, with patterns happening at many temporal and spatial scales all at once. So, we could choose to study the same predator-prey interactions at the scale of meters and minutes or 100s of km and months, and we would likely find very different drivers of patterns. The topic of scale is particularly relevant in regard to whale foraging, as marine mammals employ different sensory methods to locate prey at different spatial scales (Torres 2017).
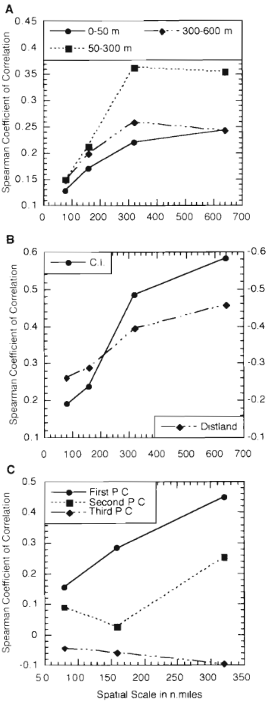
Among the first papers to conduct multi-scale research on whale foraging was Jaquet and Whitehead, 1996. Here, they studied sperm whale distribution in relation to various physical and environmental variables. Analysis showed that the main drivers of sperm whale distribution were secondary productivity (e.g., bacteria and zooplankton), underwater topography, and the gradient between deep water and surface water productivity. However, these drivers had a different impact depending on the spatial scale. There was no correlation between the drivers and sperm whale distribution at small scales < 320 nautical miles. However, at large scales >= 320 nautical miles, female sperm whale distribution was correlated with high secondary productivity and steep underwater topography. These important findings demonstrate that small scale distribution of prey alone does not drive the distribution of sperm whale predators in this study region, while other factors contribute to predator movement.
Figure 1. Figure reproduced from Jaquet & Whitehead, 1996. Plots show how the Spearman correlation results between sperm whale density and environmental variables change across multiple spatial scales. (A) Prey distribution, (B) distance to shore and bathymetric contour, and (C) the three main environmental drivers (secondary productivity, topography, and the deep water productivity gradient).
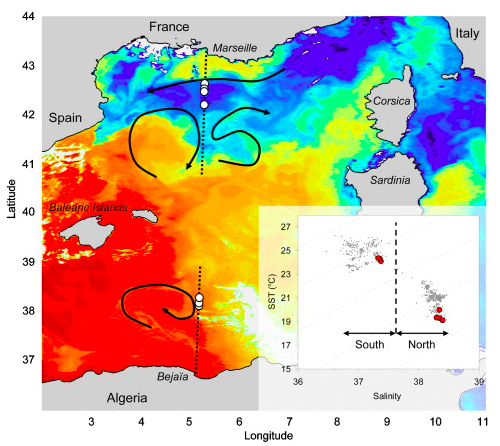
Ten years later, a study on Mediterranean fin whales tackled a similar question of how interactions between prey and predator change at multiple scales. However, their work investigated responses to both spatial and temporal scale changes. Through spatial modeling relative to oceanographic factors, Cotté et al. 2009 found that at a large-scale (year and ocean basin-wide), fin whales demonstrated two distinct distribution patterns: in the summer they were aggregated, and in the winter they were more dispersed. However, at the meso-scale (weeks -months, and 20-100 km) fin whale fidelity switched to colder, saltier waters with steeper topography and temperature gradients. Based on these results, the authors concluded that at the large scale, whale movement was driven by annually persistent prey abundance. At smaller scales, prey aggregations are less predictable, thus the authors suggest that whale movement at the meso-scale is driven by physical processes, such as frontal zones and strong currents.
Figure 2. Figure reproduced from Cotté et. al 2009. Map shows Mediterranean fin whale distribution against oceanographic conditions. Color gradient indicates sea surface temperature (SST), fin whale observations shown in white and red circles, black arrows show current direction, with inset temperature/salinity diagram for September 28-30th 2006.
A key takeaway from these papers is that it is important to investigate how processes and responses can vary at different scales, because results can sometimes depend on the time and space measurement applied in the analysis. For my thesis, I will explore which drivers take a front seat role in gray whale foraging at both fine and meso-scales. I am interested to compare my results on the relationships between PCFG gray whales and their zooplankton prey to the results from the above described studies. Stay tuned for more updates!
Did you enjoy this blog? Want to learn more about marine life, research, and conservation? Subscribe to our blog and get weekly updates and more! Just add your name into the subscribe box on the left panel.
References:
Cotté, C., Guinet, C., Taupier-Letage, I., Mate, B., & Petiau, E. (2009). Scale-dependent habitat use by a large free-ranging predator, the Mediterranean fin whale. Deep Sea Research Part I: Oceanographic Research Papers, 56(5), 801-811.
Jaquet, N., & Whitehead, H. (1996). Scale-dependent correlation of sperm whale distribution with environmental features and productivity in the South Pacific. Marine ecology progress series, 135, 1-9.
Torres, L. G. (2017). A sense of scale: Foraging cetaceans’ use of scale‐dependent multimodal sensory systems. Marine Mammal Science, 33(4), 1170-1193.
In human cultures, how you sound is often an indicator of where you are from. Have you ever taken a linguistics quiz that tries to guess what part of the United States you grew up in? Questions about whether you pronounce the sugary sweet treat caramel as “carr-mul” or “care-a-mel”, whether you say “soda” or “pop”, or whether a certain type of intersection is called a “roundabout”, “rotary”, or “traffic circle” are used to make a guess at where in the country you were raised. I have spent time in the United States, Australia, and New Zealand, I was amused to learn that the shoes you might wear in summertime can be called flip flops, slippers, thongs, or jandals, depending on which English-speaking country you are in. We know that listening to how someone speaks can tell us about their heritage or culture. As it turns out, the same is true for blue whales. We can learn a lot about blue whales by listening to them.
A blue whale comes up for air in the South Taranaki Bight, New Zealand. We catch only a short glimpse of these ocean giants when they are at the surface. By listening to their vocalizations using acoustic recordings, we can gain a whole new perspective on their lives. Photo by D. Barlow.
Sound is an incredibly important sense to marine mammals, particularly since sound waves can efficiently transmit over long distances in the ocean where other senses, such as vision or smell, are limited. Therefore, passive acoustic monitoring—placing hydrophones underwater to listen for an extended period of time and record the sounds of animals and their environment—is a highly effective tool for studying marine mammals, including blue whales. Throughout the world, blue whales sing. In this case, “song” is defined as a limited number of sound types that are produced in succession to form a recognizable pattern (McDonald et al. 2006). These songs are presumed to be produced by males only, most likely used to maintain associations and mediate social interactions, and seem to play a role in reproduction (Oleson et al. 2007, Lewis et al. 2018). Furthermore, these songs are highly stereotyped, and stable over decadal scales (McDonald et al. 2006).
Figure reproduced from McDonald et al. (2006), illustrating the variation and in blue whale songs from different geographic regions, and their stability over time: Recordings from New Zealand (A), the Central North Pacific (B), Australia (C), the Northeast Pacific (D) and North Indian Ocean (E) illustrate the stable character of the blue whale song over long time periods. All song types for which long time spans of recordings are available show some frequency drift through time, but only minor change in character. These examples were chosen because recordings over a significant time span were available to the authors in raw form, and not because these song types are more stable than the others.
Fascinatingly, blue whale songs have acoustic characteristics that are distinct between geographic regions. A blue whale in the northeast Pacific sings a different song than a blue whale in the north Atlantic; the song heard around Australia is distinct from the one sung off the coast of Chile, and so on. Therefore, differences in blue whale songs between areas can be used as a provisional hypothesis about population structure (McDonald et al. 2006, Samaran et al. 2013, Balcazar et al. 2015). Vocalizations may evolve more rapidly than traditional markers such as genetics or morphology that are often used to delineate populations, particularly in long-lived mammalian species such as blue whales (McDonald et al. 2006).
Figure reproduced from McDonald et al. (2006): Blue whale residence and population divisions suggested from their song types. Arrows indicate the direction of seasonal movements.
Despite the general rule of thumb that population-specific blue whale songs occur in separate geographic regions, there are examples throughout the southern hemisphere where songs from different populations overlap and are recorded in the same location (Samaran et al. 2010, 2013, Tripovich et al. 2015, McCauley et al. 2018, Buchan et al. 2020, Leroy et al. 2021). However, these examples may be instances where the populations temporally or ecologically partition their use of the area. For example, there may be differences in the timing of peak occurrence so that overlap is minimized by alternating which population is predominantly present in different seasons (Leroy et al. 2018). Alternatively, whales from different populations may overlap in space and time, but occupy different ecological niches at the same site. In this case, an area may simultaneously be a migratory corridor for one population and a foraging ground for another (Tripovich et al. 2015).
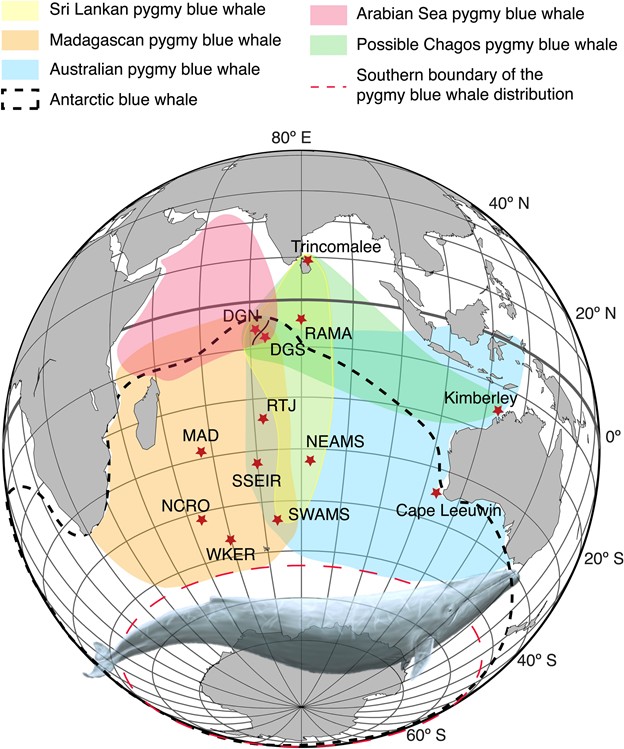
Figure reproduced from Leroy et al. (2021): Distribution of the five blue whale acoustic populations of the Indian Ocean: the Sri Lankan—NIO (yellow); Madagascan—SWIO (orange); Australian—SEIO (blue); and Arabian Sea—NWIO (red) pygmy blue whales; the hypothesized Chagos pygmy blue whale (green); and the Antarctic blue whale (black dashed line). These distributions have been inferred from the acoustic recordings conducted in the area. The long-term recording sites used to infer these distribution areas are indicated by red stars. Blue whale illustration by Alicia Guerrero.
In the South Taranaki Bight (STB) region of New Zealand, where the GEMM lab has been studying blue whales for the past decade (Torres 2013), the New Zealand song type is recorded year-round (Barlow et al. 2018). New Zealand blue whales rely on a productive upwelling system in the STB that supports an important foraging ground (Barlow et al. 2020, 2021). Antarctic blue whales also seasonally pass through New Zealand waters, likely along their migratory pathway between polar feeding grounds and lower latitude areas (Warren et al. 2021). What does it mean in terms of population connectivity or separation when two different populations occasionally share the same waters? How do these different populations ecologically partition the space they occupy? What drives their differing occurrence patterns? These are the sorts of questions I am diving into as we continue to explore the depths of our acoustic recordings from the STB region. We still have a lot to learn about these blue whales, and there is a lot to be learned through listening.
References:
Balcazar NE, Tripovich JS, Klinck H, Nieukirk SL, Mellinger DK, Dziak RP, Rogers TL (2015) Calls reveal population structure of blue whales across the Southeast Indian Ocean and the Southwest Pacific Ocean. J Mammal 96:1184–1193.
Barlow DR, Bernard KS, Escobar-Flores P, Palacios DM, Torres LG (2020) Links in the trophic chain: Modeling functional relationships between in situ oceanography, krill, and blue whale distribution under different oceanographic regimes. Mar Ecol Prog Ser 642:207–225.
Barlow DR, Klinck H, Ponirakis D, Garvey C, Torres LG (2021) Temporal and spatial lags between wind, coastal upwelling, and blue whale occurrence. Sci Rep 11:1–10.
Barlow DR, Torres LG, Hodge KB, Steel D, Baker CS, Chandler TE, Bott N, Constantine R, Double MC, Gill P, Glasgow D, Hamner RM, Lilley C, Ogle M, Olson PA, Peters C, Stockin KA, Tessaglia-hymes CT, Klinck H (2018) Documentation of a New Zealand blue whale population based on multiple lines of evidence. Endanger Species Res 36:27–40.
Buchan SJ, Balcazar-Cabrera N, Stafford KM (2020) Seasonal acoustic presence of blue, fin, and minke whales off the Juan Fernández Archipelago, Chile (2007–2016). Mar Biodivers 50:1–10.
Leroy EC, Royer JY, Alling A, Maslen B, Rogers TL (2021) Multiple pygmy blue whale acoustic populations in the Indian Ocean: whale song identifies a possible new population. Sci Rep 11:8762.
Leroy EC, Samaran F, Stafford KM, Bonnel J, Royer JY (2018) Broad-scale study of the seasonal and geographic occurrence of blue and fin whales in the Southern Indian Ocean. Endanger Species Res 37:289–300.
Lewis LA, Calambokidis J, Stimpert AK, Fahlbusch J, Friedlaender AS, Mckenna MF, Mesnick SL, Oleson EM, Southall BL, Szesciorka AR, Širović A (2018) Context-dependent variability in blue whale acoustic behaviour. R Soc Open Sci 5.
McCauley RD, Gavrilov AN, Jolli CD, Ward R, Gill PC (2018) Pygmy blue and Antarctic blue whale presence , distribution and population parameters in southern Australia based on passive acoustics. Deep Res Part II 158:154–168.
McDonald MA, Mesnick SL, Hildebrand JA (2006) Biogeographic characterisation of blue whale song worldwide: using song to identify populations. J Cetacean Res Manag 8:55–65.
Oleson EM, Wiggins SM, Hildebrand JA (2007) Temporal separation of blue whale call types on a southern California feeding ground. Anim Behav 74:881–894.
Samaran F, Adam O, Guinet C (2010) Discovery of a mid-latitude sympatric area for two Southern Hemisphere blue whale subspecies. Endanger Species Res 12:157–165.
Samaran F, Stafford KM, Branch TA, Gedamke J, Royer J, Dziak RP, Guinet C (2013) Seasonal and Geographic Variation of Southern Blue Whale Subspecies in the Indian Ocean. PLoS One 8:e71561.
Torres LG (2013) Evidence for an unrecognised blue whale foraging ground in New Zealand. New Zeal J Mar Freshw Res 47:235–248.
Tripovich JS, Klinck H, Nieukirk SL, Adams T, Mellinger DK, Balcazar NE, Klinck K, Hall EJS, Rogers TL (2015) Temporal Segregation of the Australian and Antarctic Blue Whale Call Types (Balaenoptera musculus spp.). J Mammal 96:603–610.
Warren VE, Širović A, McPherson C, Goetz KT, Radford CA, Constantine R (2021) Passive Acoustic Monitoring Reveals Spatio-Temporal Distributions of Antarctic and Pygmy Blue Whales Around Central New Zealand. Front Mar Sci 7:1–14.
Part of what makes being a graduate student so exciting is the way that learning can flip the world around: you learn a new framework or method, and suddenly everything looks a little different. I am experiencing this fabulous phenomenon lately as I learn to collect and process active acoustic data, which can reveal the distribution and biomass of animals in the ocean – including those favored by foraging whales off of Oregon, like the tiny shrimp-like krill.
We know that whales seek out the dense, energy-rich swarms that krill form, and that knowing where to expect krill can give us a leg up in anticipating whale distributions. Project OPAL (Overlap Predictions About Large whales) seeks to model and provide robust predictions of whale distributions off the coast of Oregon, so that managers can make spatially discrete decisions about potential fishery closures, minimizing burdens to fishermen while also maximizing protection of whales. We hope that including prey in our ecosystem models will help this effort, and working on this aim is one of the big tasks of my PhD.
So, how do we know where to expect krill to be off the coast of Oregon? Acoustic tools give us the opportunity to flip the world upside down: we use a tool called an echosounder to eavesdrop on the ocean, yielding visual outputs like the ones below that let us “see” and interpret sound.
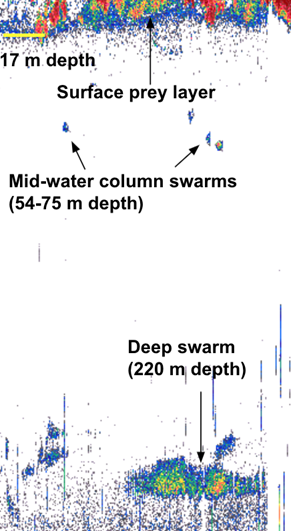
Echograms like these reveal features in the ocean that scatter “pings” of sound, and interpreting these signals can show life in the water column.
This is how it works. The echosounder emits pulses of sound at a known frequency, and then it listens for their return after it bounces of the sea floor or things in the water column. Based on sound experiments in the laboratory, we know to expect our krill species, Euphausia pacifica and Thysanoessa spinifera, to return those echoes at a characteristic decibel level. By constantly “pinging” the water column with this sound, we can record a continuous soundscape along the cruise track of a vessel, and analyze it to identify the animals and features recorded.
I had the opportunity to use an echosounder for the first time recently, on the first HALO cruise. We deployed the echosounder soon after sunrise, 65 miles offshore from Newport. After a little fiddling and troubleshooting, I was thrilled to start “listening” to the water; I was able to see the frothy noise at its surface, the contours of the seafloor, and the pixelated patches that indicate prey in between. Although it’s difficult to definitively identify animals only based on the raw output, we saw swarms that looked like our beloved krill, and other aggregations that suggested hake. Sometimes, at the same time that the team of visual observers on the flying bridge of the vessel sighted whales, I also saw potential prey on the echogram.
I spent much of the HALO cruise monitoring incoming data from the transducer on the SIMRAD EK60. Photo: Marissa Garcia.
I’m excited to keep collecting these data, and grateful that I can also access acoustic data collected by others. Many research vessels use echosounders while they are underway, including the NOAA Ship Bell M. Shimada, which conducts cruises in the Northern California Current several times a year. Starting in 2018, GEMM Lab members have joined these cruises to conduct marine mammal surveys.
This awesome pairing of data types means that we can analyze the prey that was available at the time of marine mammal sightings. I’ve been starting to process acoustic data from past Northern California Current cruises, eavesdropping on the preyscape in places that were jam-packed with whales, such as this echogram from the September 2020 cruise, below.
An echogram from the September 2020 NCC cruise shows a great deal of prey at different depths.
Like a lot of science, listening to animals in the sea comes down to occasional bursts of fieldwork followed by a lot of clicking on a computer screen during data analysis. This analysis can be some pretty fun clicking, though – it’s amazing to watch the echogram unfurl, revealing the preyscape in a swath of ocean. I’m excited to keep clicking, and learn what it can tell us about whale distributions off of Oregon.
With the GRANITE field season officially over, we are now processing all of the data we collected this summer. For me, I am starting to go through all the drone videos to take snapshots of each whale to measure their body condition. As I go through these videos, I am reflecting on the different experiences I am fortunate enough to have with flying different drones, in different environments, over different species of baleen whales: blue, humpback, Antarctic minke, and now gray whales. Each of these species have a different morphological design and body shape (Woodward et al., 2006), which leads to different behaviors that are noticeable from the drone. Drones create immense opportunity to learn how whales thrive in their natural environments [see previous blog for a quick history], and below are some of my memories from above.
I first learned how drones could be used to study the morphology and behavior of large marine mammals during my master’s degree at Duke University, and was inspired by the early works of John Durban (Durban et al., 2015, 2016) Fredrick Christiansen (Christiansen et al., 2016) and Leigh Torres (Torres et al., 2018). I immediately recognized the value and utility of this technology as a new tool to better monitor the health of marine mammals. This revelation led me to pursue a PhD with the Duke University Marine Robotics and Remote Sensing (MaRRS) Lab led by Dr. Dave Johnston where I helped further develop tools and methods for collecting drone-based imagery on a range of species in different habitats.
When flying drones over whales, there are a lot of moving parts; you’re on a boat that is moving, flying something that is moving, following something that is moving. These moving elements are a lot to think about, so I trained hard, so I did not have to think about each step and flying felt intuitive and natural. I did not grow up playing video games, so reaching this level of comfort with the controls took a lot of practice. I practiced for hours over the course of months before my first field excursion and received some excellent mentorship and training from Julian Dale, the lead engineer in the MaRRS Lab. Working with Julian and the many hours of training helped me establish a solid foundation in my piloting skills and feel confident working in various environments on different species.
Blue whales offshore of Monterey, California.
In 2017 and 2018 I was involved in collaborative project with the MaRRS Lab and Goldbogen Lab at Stanford University, where we tagged and flew drones over blue whales offshore of Monterey, California. We traveled about an hour offshore and reliably found groups of blue whales actively feeding. Working offshore typically brought a large swell, which can often make landing the drone back into your field partner’s hands tricky as everything is bobbing up and down with the oscillations of the swell. Fortunately, we worked from a larger research vessel (~56 ft) and quickly learned that landing the drone in the stern helped dampen the effects of bobbing up and down. The blue whales we encountered often dove to a depth of around 200 m for about 20-minute intervals, then come to the surface for only a few minutes. This short surface period provided only a brief window to locate the whale once it surfaced and quickly fly over it to collect the imagery needed before it repeated its dive cycle. We learned to be patient and get a sense of the animal’s dive cycle before launch in order to time our flights so the drone would be in the air a couple of minutes before the whale surfaced.
Once over the whales, the streamlined body of the blue whales was noticeable, with their small, high aspect ratio flippers and fluke that make them so well adapted for fast swimming in the open ocean (Fig. 1) (Woodward et al., 2006). I also noticed that because these whales are so large (often 21 – 24 m), I often flew at higher altitudes to be able fit them within the field of view of the camera. It was also always shocking to see how small the tagging boat (~8 m) looked when next to Earth’s largest creatures.
Figure 1. Two blue whales surface after a deep dive offshore of Monterey, Ca. (Image credit: Duke University Marine Robotics and Remote Sensing under NOAA permit 14809-03)
Antarctic minke whales and humpback whales along the Western Antarctic PeninsulaA lot of the data included in my dissertation came from work along the Western Antarctic Peninsula (WAP), which had a huge range of weather conditions, from warm and sunny days to cold and snowy/foggy/rainy/windy/icy days. A big focus was often trying to keep my hands warm, as it was often easier to fly without gloves in order to better feel the controls. One of the coldest days I remember was late in the season in mid-June (almost winter!) in Wilhemina Bay where ice completely covered the bay in just a couple hours, pushing the whales out into the Gerlache Strait; I suspect this was the last ice-free day of the season. Surprisingly though, the WAP also brought some of the best conditions I have ever flown in. Humpback and Antarctic minke whales are often found deep within the bays along the peninsula, which provided protection from the wind. So, there were times where it would be blowing 40 mph in the Gerlache Strait, but calm and still in the bays, such as Andvord Bay, which allowed for some incredible conditions for flying. Working from small zodiacs (~7 m) allowed us more maneuverability for navigating around or through the ice deep in the bays (Fig. 2)
Figure 2. Navigating through ice-flows along the Western Antarctic Peninsula. (Image credit: Duke University Marine Robotics and Remote Sensing under NOAA permit 14809-03 and ACA permits 2015-011 and 2020-016.)
Flying over Antarctic minke whale was always rewarding, as they are very sneaky and can quickly disappear under ice flows or in the deep, dark water. Flying over them often felt like a high-speed chase, as their small streamlined bodies makes them incredibly quick and maneuverable, doing barrel rolls, quick banked turns, and swimming under and around ice flows (Fig. 3). There would often be a group between 3-7 individuals and it felt like they were playing tag with each other – or perhaps with me!
Figure 3. Two Antarctic minke whales swimming together along the Western Antarctic Peninsula. (Image credit: Duke University Marine Robotics and Remote Sensing under NOAA permit 14809-03 and ACA permits 2015-011 and 2020-016.)
Humpbacks displayed a wide range of behaviors along the WAP. Early in the season they continuously fed throughout the entire day, often bubble net feeding in groups typically of 2-5 animals (Fig. 4). For as large as they are, it was truly amazing to see how they use their pectoral fins to perform quick accelerations and high-speed maneuvering for tight synchronized turns to form bubble nets, which corral and trap their krill, their main food source (Fig. 4) (Woodward et al., 2006). Later in the season, humpbacks switched to more resting behavior in the day and mostly fed at night, taking advantage of the diel vertical migration of krill. This behavior meant we often found humpbacks snoozing at the surface after a short dive, as if they were in a food coma. They also seemed to be more curious and playful with each other and with us later in the season (Fig. 5).
We also encountered a lot of mom and calf pairs along the WAP. Moms were noticeably skinny compared to their plump calf in the beginning of the season due to the high energetic cost of lactation (Fig. 6). It is important for moms to regain this lost energy throughout the feeding season and begin to wean their calves. I often saw moms refusing to give milk to their nudging calf and instead led teaching lessons for feeding on their own.
Figure 4. Two humpback whales bubble-net feeding early in the feeding season (December) along the Western Antarctic Peninsula. (Image credit: Duke University Marine Robotics and Remote Sensing under NOAA permit 14809-03 and ACA permits 2015-011 and 2020-016.)Figure 5. A curious humpback whale dives behind our Zodiac along the Western Antarctic Peninsula. (Image credit: Duke University Marine Robotics and Remote Sensing under NOAA permit 14809-03 and ACA permits 2015-011 and 2020-016.)Figure 6. A mom and her calf rest at the surface along the Western Antarctic Peninsula. Note how the mom looks skinnier compared to her plump calf, as lactation is the most energetically costly phase of the reproductive cycle. (Image credit: Duke University Marine Robotics and Remote Sensing under NOAA permit 14809-03 and ACA permits 2015-011 and 2020-016.)
Gray whales off Newport, Oregon
All of these past experiences helped me quickly get up to speed and jump into action with the GRANITE field team when I officially joined the GEMM Lab this year in June. I had never flown a DJI Inspire quadcopter before (the drone used by the GEMM Lab), but with my foundation piloting different drones, some excellent guidance from Todd and Clara, and several hours of practice to get comfortable with the new setup, I was flying over my first gray whale by day three of the job.
The Oregon coast brings all sorts of weather, and some days I strangely found myself wearing a similar number of layers as I did in Antarctica. Fog, wind, and swell could all change within the hour, so I learned to make the most of weather breaks when they came. I was most surprised by how noticeably different gray whales behave compared to the blue, Antarctic minke, and humpback whales I had grown familiar with watching from above. For one, it is absolutely incredible to see how these huge whales use their low-aspect ratio flippers and flukes (Woodward et al., 2006) to perform low-speed, highly dynamic maneuvers to swim in very shallow water (5-10 m) so close to shore (<1m sometimes!) and through kelp forest or surf zones close to the beach. They have amazing proprioception, or the body’s ability to sense its movement, action, and position, as gray whales often use their pectoral fins and fluke to stay in a head standing position (see Clara Bird’s blog) to feed in the bottom sediment layer, all while staying in the same position and resisting the surge of waves that could smash them against the rocks (Video 1) . It is also remarkable how the GEMM Lab knows each individual whale based on natural skin marks, and I started to get a better sense of each whale’s behavior, including where certain individuals typically like to feed, or what their dive cycle might be depending on their feeding behavior.
Video 1. Two Pacific Coast Feeding Group (PCFG) gray whales “head-standing” in shallow waters off the coast of Newport, Oregon. NOAA/NMFS permit #21678
I feel very fortunate to be a part of the GRANITE field team and to contribute to data collection efforts. I look forward to the data analysis phase to see what we learn about how the morphology and behavior of these gray whales to help them thrive in their environment.
References:
Christiansen, F., Dujon, A. M., Sprogis, K. R., Arnould, J. P. Y., and Bejder, L. (2016).Noninvasive unmanned aerial vehicle provides estimates of the energetic cost of reproduction in humpback whales. Ecosphere 7, e01468–18.
Durban, J. W., Fearnbach, H., Barrett-Lennard, L. G., Perryman, W. L., & Leroi, D. J. (2015). Photogrammetry of killer whales using a small hexacopter launched at sea. Journal of Unmanned Vehicle Systems, 3(3), 131-135.
Durban, J. W., Moore, M. J., Chiang, G., Hickmott, L. S., Bocconcelli, A., Howes, G., et al.(2016). Photogrammetry of blue whales with an unmanned hexacopter. Mar. Mammal Sci. 32, 1510–1515.
Torres, L. G., Nieukirk, S. L., Lemos, L., & Chandler, T. E. (2018). Drone up! Quantifying whale behavior from a new perspective improves observational capacity. Frontiers in Marine Science, 5, 319.
Woodward, B. L., Winn, J. P., and Fish, F. E. (2006). Morphological specializations of baleen whales associated with hydrodynamic performance and ecological niche. J. Morphol. 267, 1284–1294.
Tagging a whale is no easy feat, nor is it without some impact to the whale – no matter how minimized through the use of non-penetrating suction cup tags. Yet, in August 2021 the GEMM Lab initiated a new phase in our research on gray whales, aimed at obtaining a better understanding of the underwater lives and energetics of a gray whale (Figure 1, top image). We captured some amazing data through these specialized, non-invasive tags that provide a brief window into their world and physiology. The video recordings from the tags showed us whales digging their heads into the benthos generating billowing clouds of sediment, likely exploiting desirable prey patches (Figure 1, middle images). We also saw foraging whales undertake dizzying spins and headstands for hours, demonstrating the fascinating maneuverability and flexibility of gray whales (Figure 1, bottom image). But what is motivating us to capture this information?
Figure 1. Top image:One of the suction cup tags attached to the right side of the gray whale “RAT”. Middle images: still images from the video recorded on the tag attached to gray whale “DICE”, where left image depicts the whale “seeing” our research vessel RUBY as “DICE” comes to the surface to breathe, and the right image shows a large sediment cloud generated by “DICE” as it digs its head into the benthos feeding on benthic prey. Bottom image: a visualization of a trackline (grey, blue, & yellow line) of a foraging gray whale (indicated by the blue arrow). The yellow segments depict times during which the whale was on its side or headstanding (suggesting feeding behavior), while the blue segments represent when the whale was traveling or surfacing. You can see how often in the track the whale is presumably feeding based on the amount of yellow segments in the track. Also, note the amount of circular loops the whale performs, displaying the maneuverability of foraging gray whales. Source: GEMM Lab.
The GEMM Lab has researched the ecology and physiology of Pacific Coast Feeding Group (PCFG) gray whales since 2015. Our efforts have filled crucial knowledge gaps to better understand this sub-group of the Eastern North Pacific (ENP) gray whale population. We now know that gray whale body condition increases throughout a foraging season and can fluctuate considerably between years (Soledade Lemos et al. 2020). Additionally, body condition varies significantly by reproductive state, with calves and pregnant females displaying higher body conditions (Soledade Lemos et al. 2020). We have also validated and quantified fecal steroid and thyroid hormone metabolite concentrations, providing us with thresholds to identify a stressed vs. a not stressed whale based on its hormone levels (Lemos et al. 2020). These validations have allowed us to make correlations between poor body condition and the steroid hormone cortisol which confirm that slim whales are stressed, while chubby whales are relaxed (Lemos et al. 2021). These physiological results are particularly salient in the light of our recent findings that PCFG gray whales select prey quality over prey quantity when foraging (Hildebrand et al. in review) and that the caloric content of available prey species in the PCFG range vary significantly (Hildebrand et al. 2021).
While we have addressed several fundamental questions about the PCFG in the last 7 years, answering one question has led to asking 10 more questions – a common pattern in science. Given that we know (1) PCFG whales improve their body condition over the course of the foraging season (Soledade Lemos et al. 2020), (2) PCFG females are able to successfully give birth to and wean calves (Calambokidis & Perez 2017), and (3) certain prey in the PCFG region are of higher caloric value than prey in the ENP Arctic foraging grounds (Hildebrand et al. 2021), a big question that we continue to scratch our heads about is why does the PCFG sub-group have such a small abundance (~250 individuals; Calambokidis et al. 2017) in comparison to the much larger ENP population (~21,000 individuals; Stewart & Weller 2021). Several hypotheses have been suggested including that the energetic costs of feeding may differ between ENP and PCFG whales, with the latter having to expend more energy to obtain prey due to the different foraging behaviors employed (Torres et al. 2018) to obtain diverse prey types, thus justifying the larger abundance of the ENP (Hildebrand et al. 2021).
Quantifying the energetic cost of baleen whale behaviors is not simple. However, the development of animal-borne tags has allowed scientists to make big strides regarding behavioral cost quantification. The majority of this work has focused on rorqual whales (i.e., blue, humpback, fin whales; e.g., Goldbogen et al. 2013; Cade et al. 2016) as their characteristic lunge-feeding strategy produces a distinct signal in the accelerometer sensors integrated within the tags, making feeding events easier to identify. Gray whales, unlike rorquals, do not lunge-feed. ENP gray whales predominantly feed benthically; diving down to the benthos where they turn onto their side and suction mouthfuls of soft sediment (mud) that contains amphipods that they filter out of the mud (Nerini & Oliver 1983). PCFG whales feed benthically as well, but they also use a number of other feeding behaviors to obtain a variety of prey in a variety of benthic habitats, including headstands, bubble blasts, and sharking (Torres et al. 2018). The above-mentioned gray whale feeding behaviors involve much subtler movements than the powerful, distinctive lunges displayed by rorquals, yet they undoubtedly still incur some energetic cost to the whales. However, exactly how energetically costly the various gray whale feeding behaviors are remains unknown.
One of the three suction cup tags we deployed on gray whales. Dr. Cade printed special “kelp shields” (blue part of the tag) to prevent kelp from potentially getting caught underneath the tag since PCFG whales often forage on reefs with a lot of kelp. This tag includes a video camera (the lens can be seen in the center of the tag) to record video of the whale’s underwater behavior. Source: L. Torres.
This knowledge gap is one of the reasons why the GEMM Lab initiated a new project in close collaboration with Dr. Dave Cade from Stanford University and John Calambokidis from Cascadia Research Collective to quantify and understand the energetics and underwater behavior of gray whales using suction-cup tags. The project was kick-started with a very successful pilot effort the week of August 16th this year. Tags were placed on the backs of three different PCFG gray whales with a long carbon fiber pole and attached to the whales with four suction cups. The tags recorded video, position, accelerometry, and magnetometry data, which we will use to recreate the animal’s movements (pitch, roll), heading, trackline, and environment. Although the weather forecast did not look promising for most of the week, we lucked out with perfect conditions for one day during which we managed to deploy three tags on three different gray whales that are well-known, long-term study animals of the GEMM Lab. The tags stayed on the whales for 1-6 hours and were all recovered (including an adventurous trip up the Alsea River which involved a kayak deployment!).
Left: Postdoc Alejandro Ajo Fernandez uses a Yagi antenna to try and locate a tagged whale since the tags have a transmitter that sends a signal when the whale is at the surface to breathe. Right: Dr. Dave Cade proudly showing off the tag that floated up the Alsea River and required a kayak to recover. Source: L. Torres.
Dr. Cade spent the rest of the week teaching GEMM Lab PI Leigh Torres, University of British Columbia Master’s student Kate Colson (who is co-advised by Leigh and Dr. Andrew Trites), and myself the intricacies of data download, processing, and preliminary analysis of the tag data. For her Master’s research, Kate will develop a bioenergetics model for the PCFG sub-group that includes data on foraging energetics (estimated from the tag data) and prey availability in the PCFG foraging range. I plan on using the tag data to assess behavior patterns of PCFG whales relative to habitat as part of my PhD research. All together analysis of the data from these short-term tag deployments will help us get closer to understanding the behavioral choices, habitat needs, and energetic trade-offs of whales living in a rapidly changing ocean. With the success of this pilot effort, we plan to conduct another suction-cup tagging effort next summer to hopefully capture and explore more mysterious underwater behaviors of the PCFG.
An ecstatic team at the end of a very long yet successful day of suction cup tagging. Bottom (from left): Leigh Torres, Lisa Hildebrand, ClaraBird, Dave Cade, KC Bierlich. Top: John Calambokidis.
This project was funded by sales and renewals of the special Oregon whale license plate, which benefits MMI. We gratefully thank all the gray whale license plate holders, who made this research effort possible.
Literature cited
Cade, D. E., Friedlaender, A. S., Calambokidis, J., & Goldbogen, J. A. 2016. Kinematic diversity in rorqual whale feeding mechanisms. Current Biology 26(19):2617-2624. doi:10.1016/j.cub.2016.07.037.
Calambokidis, J., & Perez, A. 2017. Sightings and follow-up of mothers and calves in the PCFG and implications for internal recruitment. IWC Report SC/A17/GW/04 for the Workshop on the Status of North Pacific Gray Whales (La Jolla: IWC).
Calambokidis, J., Laake, J., & Perez, A. 2017. Updated analysis of abundance and population structure of seasonal gray whales in the Pacific Northwest, 1996-2015. IWC Report SC/A17/GW/05 for the Workshop on the Status of North Pacific Gray Whales (La Jolla: IWC).
Goldbogen, J. A., Friedlaender, A. S., Calambokidis, J., McKenna, M. F., Simon, M., & Nowacek, D. P. 2013. Integrative approaches to the study of baleen whale diving behavior, feeding performance, and foraging ecology. BioScience 63(2):90-100. doi:10.1525/bio.2013.63.2.5.
Hildebrand, L., Bernard, K. S., & Torres, L. G. 2021. Do gray whales count calories? Comparing energetic values of gray whale prey across two different feeding grounds in the eastern North Pacific. Frontiers in Marine Science 1008. doi:10.3389/fmars.2021.683634.
Lemos, L. S., Olsen, A., Smith, A., Burnett, J. D., Chandler, T. E., Larson, S., Hunt, K. E., & Torres, L. G. 2021. Stressed and slim or relaxed and chubby? A simultaneous assessment of gray whale body condition and hormone variability. Marine Mammal Science. doi:10.111/mms.12877.
Lemos, L.S., Olsen, A., Smith, A., Chandler, T.E., Larson, S., Hunt, K., and L.G. Torres. 2020. Assessment of fecal steroid and thyroid hormone metabolites in eastern North Pacific gray whales. Conservation Physiology 8:coaa110.
Nerini, M. K., & Oliver, J. S. 1983. Gray whales and the structure of the Bering Sea benthos. Oecologia 59:224-225. doi:10.1007/bf00378840.
Soledade Lemos, L., Burnett, J. D., Chandler, T. E., Sumich, J. L., & Torres, L. G. 2020. Intra- and inter-annual variation in gray whale body condition on a foraging ground. Ecosphere 11(4):e03094.
Stewart, J. D., & Weller, D. W. 2021. Abundance of eastern North Pacific gray whales 2019/2020. Department of Commerce, NOAA Technical Memorandum NMFS-SWFSC-639. United States: NOAA. doi:10.25923/bmam-pe91.
Torres, L.G., Nieukirk, S.L., Lemos, L., and T.E. Chandler. 2018. Drone Up! Quantifying Whale Behavior From a New Perspective Improves Observational Capacity. Frontiers in Marine Science: https://doi.org/10.3389/fmars.2018.00319.